Identification of Microflora in a Biological Brewer’s Wort Acidification Process Run Continuously for 20 Years

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Media and Growth Conditions
2.2. Preparing Cryogenic Cultures
2.3. PCR and Fingerprinting
2.4. Sequencing of 16S and 26S rDNA
2.5. Real-Time PCR
2.6. Phenolic Off-Flavor Test
3. Results
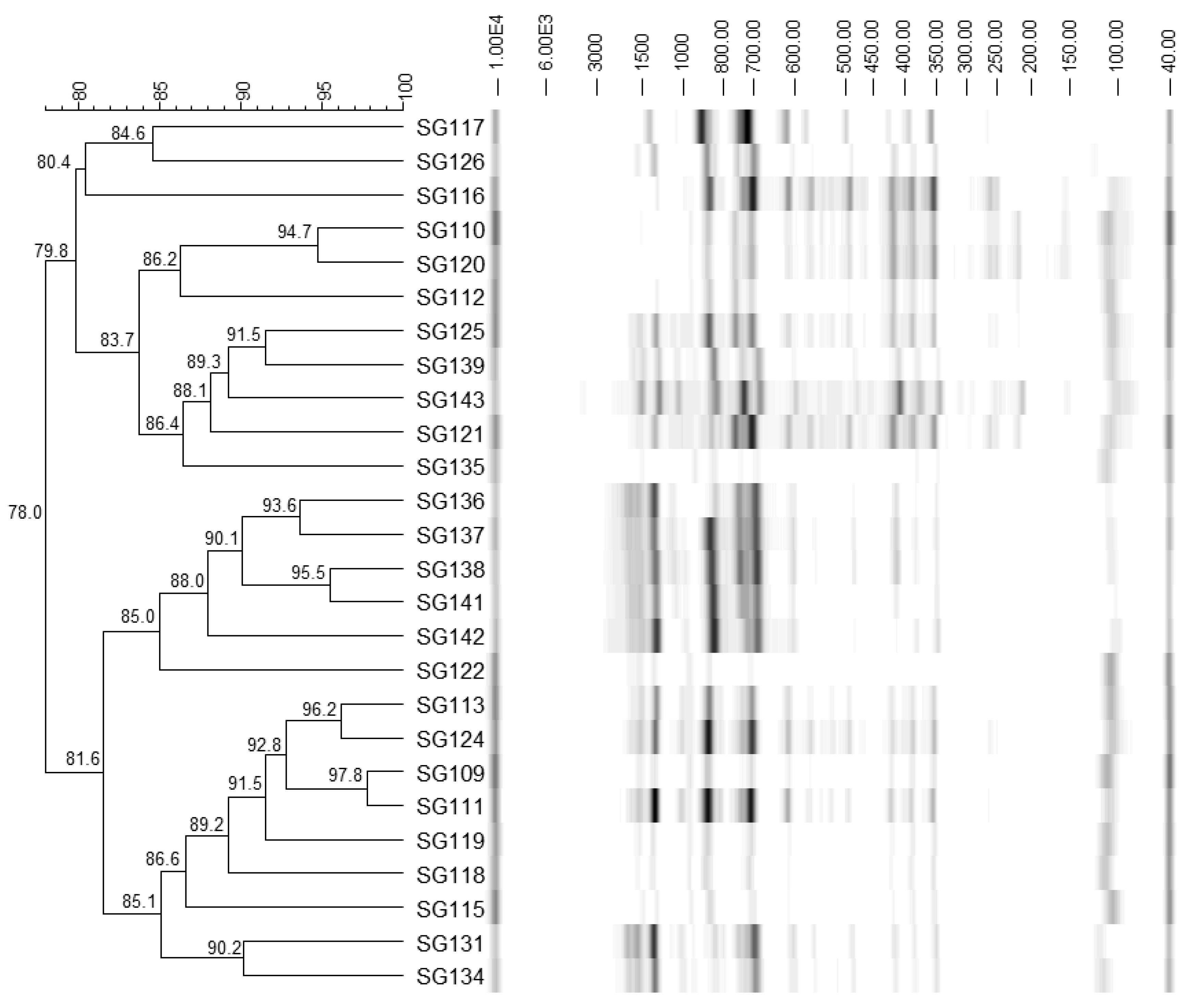
3.1. Identification of Bacteria
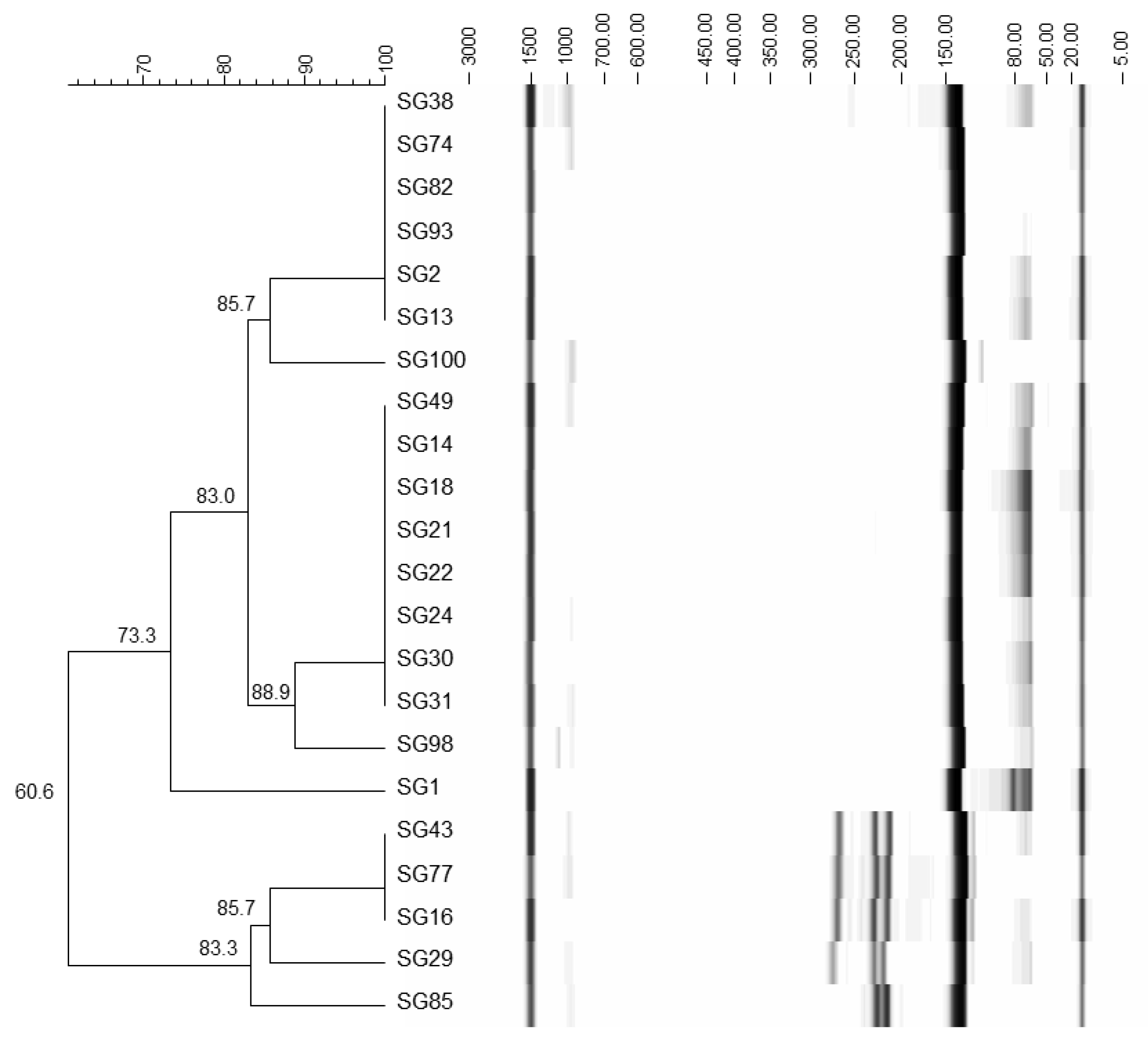
3.2. Identification of Yeasts
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Back, W. Biologische Säuerung. Monatsschr. Brauwiss. 1988, 41, 152–156. [Google Scholar]
- Lindner, P. Über ein neues in Malzmaischen vorkommendes Milchsäure bildendes Ferment. Wochenschr. Brau. 1887, 4, 437–440. [Google Scholar]
- Leichmann, G. Über die im Brennereiprozeß bei der Bereitung von Kunsthefe auftretende spontane Milchsäuregarung (Bac. Delbrücki). Zentralbl. Bakteriol. 1896, 2, 281–285. [Google Scholar]
- Francke, O. Herstellung säuerlich schmeckender, insbesondere milchsauerer Biere. Deutsches Reichspatent Nr. 180726, 5 April 1906. [Google Scholar]
- Oliver-Daumen, B. Biological acidification in the brewing process—Part 1: Review of the literature. Brauwelt Int. 1988, 256–264. [Google Scholar]
- Grützmacher, J. Biologische Säuerung in der Brauerei. Brauwelt 1991, 40, 1762–1769. [Google Scholar]
- Back, W. Ausgewählte Kapitel der Brauereitechnologie; Hans Carl: Nürnberg, Germany, 2005. [Google Scholar]
- Lowe, D.P.; Ulmer, H.M.; Barta, R.C.; Goode, D.L.; Arendt, E.K. Biological Acidification of a Mash Containing 20% Barley Using Lactobacillus amylovorus FST 1.1: Its Effects on Wort and Beer Quality. J. Am. Soc. Brew. Chem. 2005, 63, 96–106. [Google Scholar] [CrossRef]
- Lowe, D.P.; Arendt, E.K. The Use and Effects of Lactic Acid Bacteria in Malting and Brewing with Their Relationships to Antifungal Activity, Mycotoxins and Gushing: A Review. J. Inst. Brew. 2004, 110, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, A.; O’Sullivan, T.; Sinderen, D. Enhancing the Microbiological Stability of Malt and Beer—A Review. J. Inst. Brew. 2005, 111, 355–371. [Google Scholar] [CrossRef]
- Bohak, I.; Back, W.; Richter, L.; Ehrmann, M.; Ludwig, W.; Schleifer, K.H. Lactobacillus amylolyticus sp. nov., Isolated from Beer Malt and Beer Wort. Syst. Appl. Microbiol. 1998, 21, 360–364. [Google Scholar] [CrossRef]
- Goranov, B.; Denkova, Z.; Kostov, G.; Denkova, R. Possibilities for the biological acidification of mash in the production of wort: Kinetics of lactic acid production in a free cell culture of Lactobacillus delbrueckii ssp. bulgaricus M3. Presented at 50 Years FoodRDI. Food Technologies and Health, International Scientific-Practical Conference, Plovdiv, Bulgaria, 8 November 2012. [Google Scholar]
- Boulton, C.; Quain, D. Brewing Yeast and Fermentation; Blackwell Science; Iowa State University Press: Malden, MA, USA; Ames, IA, USA, 2001. [Google Scholar]
- Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Hutzler, M. Getränkerelevante Hefen—Identifizierung und Differenzierung. Wie Können Hefen Praxisrelevant Unterschieden Werden, und wie Können Identifizierungsergebnisse Technologisch Bewertet Werden? Südwestdt. Verlag für Hochschulschriften: Saarbrücken, Germany, 2010; ISBN-13: 9783838114828. [Google Scholar]
- Zhang, H.; You, C.; Ren, J.; Xu, D.; Han, M.; Liao, W. A simple one-step PCR walking method and its application of bacterial rRNA for sequencing identification. Curr. Microbiol. 2014, 68, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Ehrmann, M.A.; Müller, M.R.A.; Vogel, R.F. Molecular analysis of sourdough reveals Lactobacillus mindensis sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Nossa, C.W.; Oberdorf, W.E.; Yang, L.; Aas, J.A.; Paster, B.J.; Desantis, T.Z.; Brodie, E.L.; Malamud, D.; Poles, M.A.; Pei, Z. Design of 16S rRNA gene primers for 454 pyrosequencing of the human foregut microbiome. World J. Gastroenterol. 2010, 16, 4135–4144. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Phylogenetic relationships among yeasts of the ‘Saccharomyces complex’ determined from multigene sequence analyses. FEMS Yeast Res. 2003, 3, 417–432. [Google Scholar] [CrossRef]
- Brandl, A. Entwicklung und Optimierung von PCR-Methoden zur Detektion und Identifizierung von Brauereirelevanten Mikroorganismen zur Routine-Anwendung in Brauereien. Ph.D. Thesis, TU München, Freising, Germany, 2006. [Google Scholar]
- Salinas, F.; Garrido, D.; Ganga, A.; Veliz, G.; Martínez, C. Taqman real-time PCR for the detection and enumeration of Saccharomyces cerevisiae in wine. Food Microbial. 2009, 26, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Meier-Dörnberg, T.; Hutzler, M.; Michel, M.; Methner, F.-J.; Jacob, F. The Importance of a Comparative Characterization of Saccharomyces Cerevisiae and Saccharomyces Pastorianus Strains for Brewing. Fermentation 2017, 3, 41. [Google Scholar] [CrossRef]
- Stackebrand, E.; GOEBEL, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Thurston, P.A.; Tubb, R.S. Screening yeast strains for their ability to produce phenolic off-flavours: A simple method for determining phenols in wort and beer. J. Inst. Brew. 1981, 87, 177–179. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Goncalves, P.; Sampaio, J.P. Distinct domestication trajectories in top-fermenting beer yeasts and wine yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef] [PubMed]
- Röcken, W.; Marg, C. Nachweis von Fremdhefen in der obergärigen Brauerei. Vergleich verschiedener Nährböden. Monatsschr. Brauwiss. 1983, 1983, 276–279. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′ → 3′ | Reference |
|---|---|---|
| GTG5 | GTGGTGGTGGTGGTG | [14] |
| IGS2-314-fp | CGGGTAACCCAGTTCCTCACT GTAGCATATATTTCTTGTGTGAGAAAGGT | [15] |
| IGS2-314-rp | ||
| 16S-27f | AGAGTTTGATCM(C/A)TGGCTCAG TACGGY(C/T)TACCTTGTTACGACTT | [16] |
| 1492r | ||
| 612r | GTAAGGTTY(C/T)TNCGCGT | [17] |
| 926r | CCGTCAATTCM(C/A)TTTRAGT | [18] |
| NL1 | GCATATCAATAAGCGGAGGAAAAG GGTCCGTGTTTCAAGACGG | [19] |
| NL4 |
| Strain | Sequencing Primer | Accession Number (Length in bp) | NCBI Identification | Identities of the Target Microorganism (%) | Real-Time PCR Identification |
|---|---|---|---|---|---|
| SG1 | NL4 | KJ506733.1 (609) | S. cerevisiae | 99 | S. cerevisiae |
| SG14 | NL4 | CP006467.1 (1893211) | S. cerevisiae | 99 | S. cerevisiae |
| SG16 | NL4 | JQ672609.1 (601) | S. cerevisiae | 100 | S. cerevisiae |
| SG22 | NL4 | JQ277730.1 (9076) | S. cerevisiae | 99 | S. cerevisiae |
| SG82 | NL4 | JQ672587.1 (587) | S. cerevisiae | 99 | S. cerevisiae |
| SG85 | NL4 | JQ968592.1 (573) | S. cerevisiae | 100 | S. cerevisiae |
| SG93 | NL4 | JX103179.1 (620) | S. cerevisiae | 99 | S. cerevisiae |
| SG100 | NL4 | JX103179.1 (620) | S. cerevisiae | 100 | S. cerevisiae |
| SG109 | 1492r | Y17361.1 (1544) | L. amylolyticus | 99 | L. amylolyticus |
| NR_075048.1 (1575) | L. amylovorus | 99 | |||
| SG115 | 1492r | NR_117069.1 (1443) | L. amylolyticus | 99 | L. amylolyticus |
| NR_075048.1 (1575) | L. amylovorus | 99 | |||
| SG126 | 1492r | NR_029352.1 (1544) | L. amylolyticus | 99 | L. amylolyticus |
| EF120375.1 (1550) | L. amylovorus | 99 | |||
| SG139 | 1492r | Y17361.1 (1558) | L. amylolyticus | 99 | L. amylolyticus |
| NR_075048.1 (1575) | L. amylovorus | 99 |
| Yeast Strain | OG-COXII | SCTM | UG-LRE1 | UG300 | Sbp |
|---|---|---|---|---|---|
| S. cerevisiae SG1 | + | + | − | − | − |
| S. cerevisiae SG14 | + | + | − | − | − |
| S. cerevisiae SG16 | + | + | − | − | − |
| S. cerevisiae SG22 | + | + | − | − | − |
| S. cerevisiae SG82 | + | + | − | − | − |
| S. cerevisiae SG85 | + | + | − | − | − |
| S. cerevisiae SG93 | + | + | − | − | − |
| S. cerevisiae SG100 | + | + | − | − | − |
| Flavor | Precursor Cinnamic Acid, Product Styrene “Styrofoam-Like” | Precursor Ferulic Acid, 4-Vinylguaiacol “Clove-Like” | |
|---|---|---|---|
| Yeast | |||
| Frisinga-TUM 34/70® | − | − | |
| LeoBavaricus-TUM 68® | + | + | |
| S. cerevisiae SG1 | ++ | + | |
| S. cerevisiae SG14 | ++ | + | |
| S. cerevisiae SG16 | − | − | |
| S. cerevisiae SG22 | ++ | + | |
| S. cerevisiae SG24 | ++ | + | |
| S. cerevisiae SG29 | − | − | |
| S. cerevisiae SG82 | ++ | ++ | |
| S. cerevisiae SG85 | ++ | ++ | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutzler, M.; Čmielová, J.; Frank, T.; Brandl, A.; Jacob, F.; Michel, M. Identification of Microflora in a Biological Brewer’s Wort Acidification Process Run Continuously for 20 Years. Fermentation 2018, 4, 51. https://doi.org/10.3390/fermentation4030051
Hutzler M, Čmielová J, Frank T, Brandl A, Jacob F, Michel M. Identification of Microflora in a Biological Brewer’s Wort Acidification Process Run Continuously for 20 Years. Fermentation. 2018; 4(3):51. https://doi.org/10.3390/fermentation4030051
Chicago/Turabian StyleHutzler, Mathias, Jana Čmielová, Tobias Frank, Andreas Brandl, Fritz Jacob, and Maximilian Michel. 2018. "Identification of Microflora in a Biological Brewer’s Wort Acidification Process Run Continuously for 20 Years" Fermentation 4, no. 3: 51. https://doi.org/10.3390/fermentation4030051
APA StyleHutzler, M., Čmielová, J., Frank, T., Brandl, A., Jacob, F., & Michel, M. (2018). Identification of Microflora in a Biological Brewer’s Wort Acidification Process Run Continuously for 20 Years. Fermentation, 4(3), 51. https://doi.org/10.3390/fermentation4030051