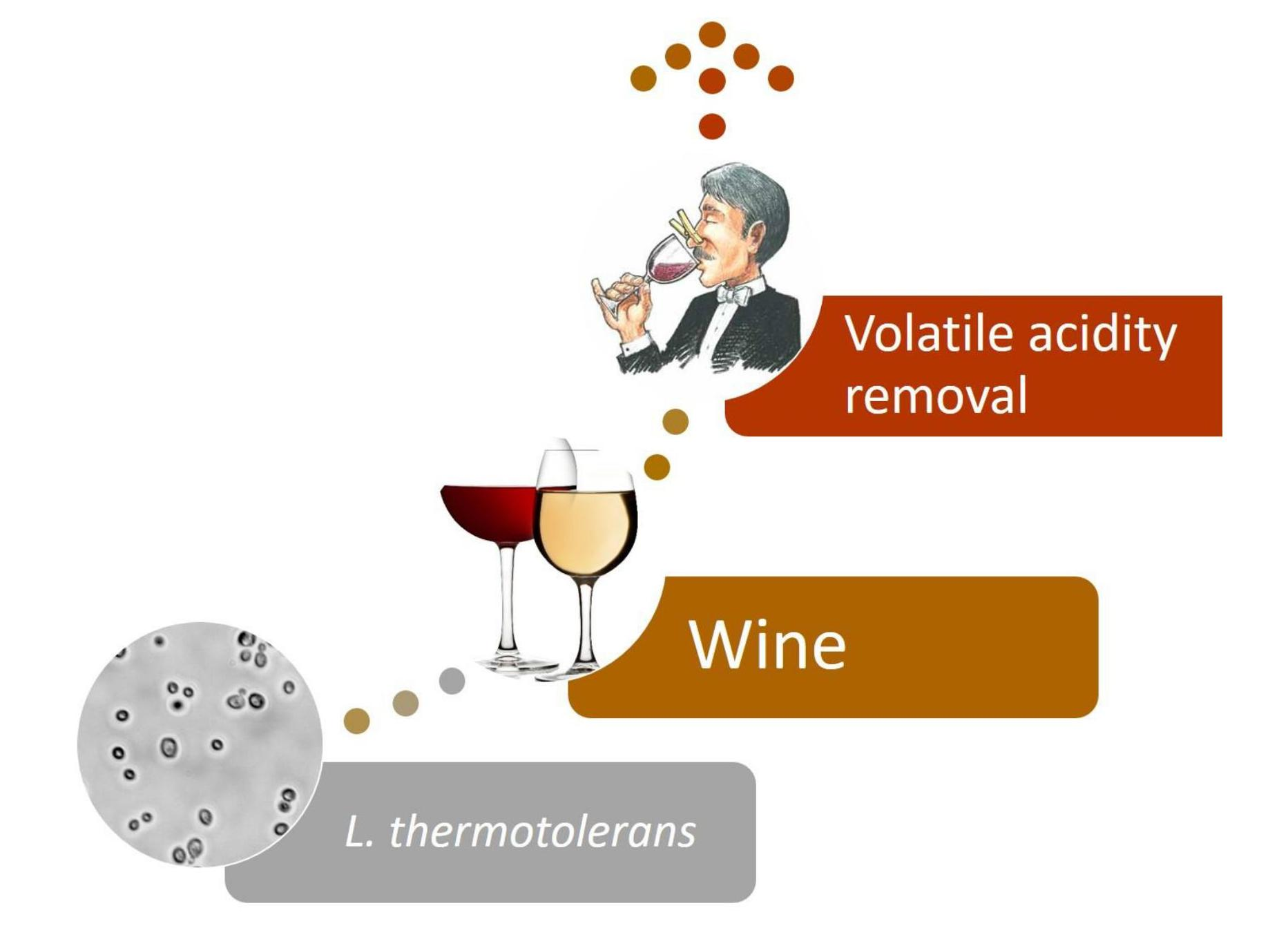
Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines


Abstract
:
1. Introduction
2. The Vinegar Taint Problem in Wine
3. L. thermotolerans’ Main Features in Alcoholic Drinks
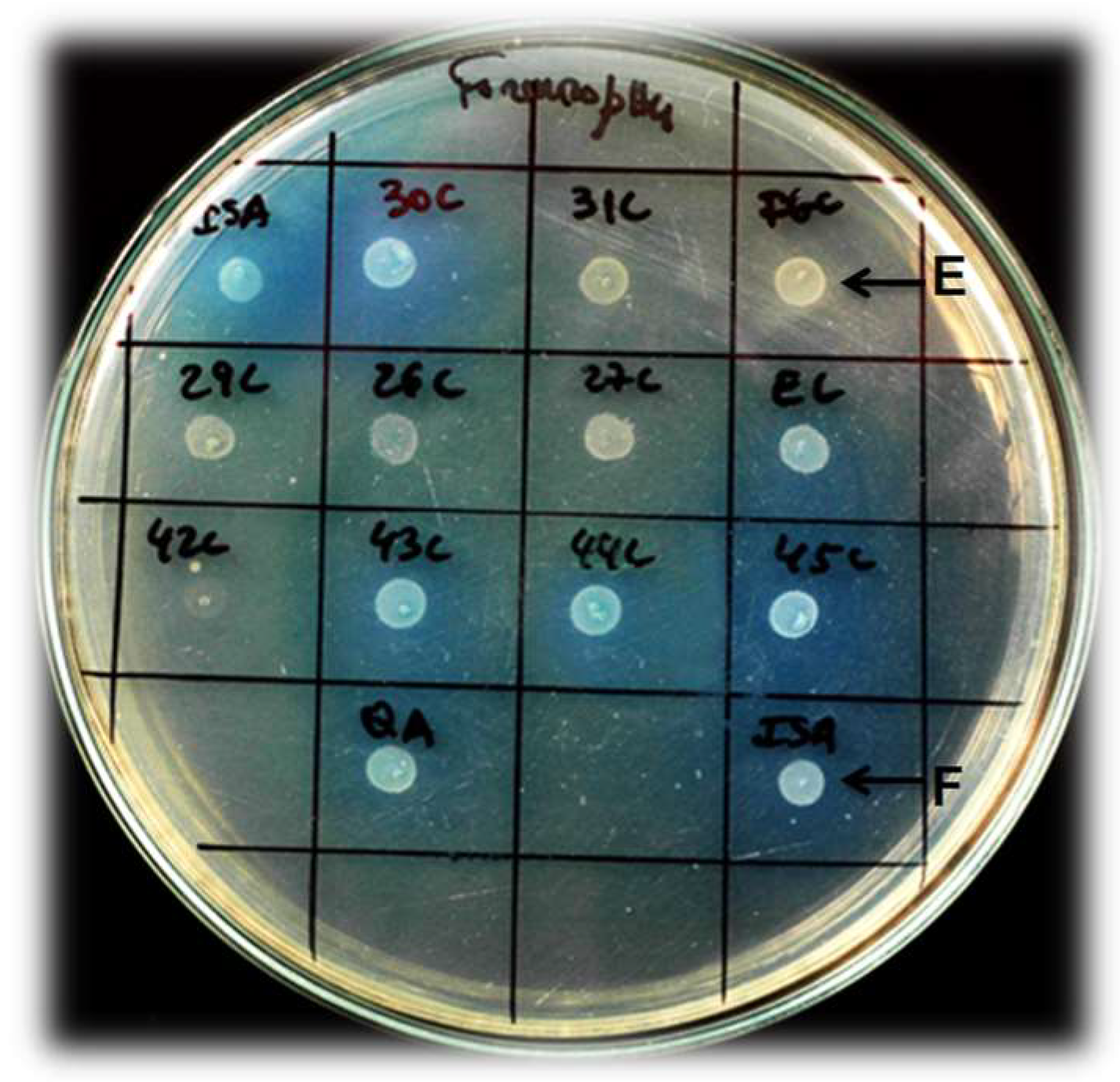
4. Strain Isolation and Wine Biodeacetification
5. Conclusions
Funding
Conflicts of Interest
References
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Gonzalez, R. Non-conventional Yeast Species for Lowering Ethanol Content of Wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past, and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.C. Saccharomyces and non-Saccharomyces competition during microvinification under different sugar and nitrogen conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef] [PubMed]
- Brice, C.; Cubillos, F.A.; Dequin, S.; Camarasa, C.; Martínez, C. Adaptability of the Saccharomyces cerevisiae yeasts to wine fermentation conditions relies on their strong ability to consume nitrogen. PLoS ONE 2018, 13, e0192383. [Google Scholar] [CrossRef] [PubMed]
- Dibalova-Culakova, H.; Alonso-del-Real, J.; Querol, A.; Sychrova, H. Expression of heterologous transporters in Saccharomyces kudriavzevii: A strategy for improving yeast salt tolerance and fermentation performance. Int. J. Food Microbiol. 2018, 268, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Minnaar, P.P.; Plessis, H.W.; du Paulsen, V.; Ntushelo, N.; Jolly, N.P.; du Toit, M. Saccharomyces cerevisiae, non-Saccharomyces yeasts and lactic acid bacteria in sequential fermentations: Effect on phenolics and sensory attributes of South African Syrah Wines. S. Afr. J. Enol. Vitic. 2017, 38, 237–244. [Google Scholar] [CrossRef]
- Hranilovic, A.; Bely, M.; Masneuf-Pomarede, I.; Jiranek, V.; Albertin, W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE 2017, 12, e0184652. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Phylogenetic circumscription of Saccharomyces, Kluyveromyces and other members of the Saccharomycetaceae, and the proposal of the new genera Lachancea, Nakaseomyces, Naumovia, Vanderwaltozyma, and Zygotorulaspora. FEMS Yeast Res. 2003, 4, 233–245. [Google Scholar] [CrossRef]
- Lachance, M.A.; Lachancea, K. The Yeasts, a Taxonomic Study; Kurtzman, C., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 511–519. [Google Scholar]
- Souciet, J.L.; Dujon, B.; Gaillardin, C.; Johnston, M.; Baret, P.V.; Cliften, P.; Sherman, D.J.; Weissenbach, J.; Westhof, E.; Wincker, P.; et al. Comparative genomics of protoploid Saccharomycetaceae. Genome Res. 2009, 19, 1696–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Dequin, S.; Barre, P. Mixed lactic acid–alcoholic fermentation by Saccharomyces cerevisiae expressing the Lactobacillus casei L (+)–LDH. Nat. Biotechnol. 1994, 12, 173–177. [Google Scholar] [CrossRef]
- Çakar, Z.P.; Turanli-Yildiz, B.; Alkim, C.; Yilmaz, U. Evolutionary engineering of Saccharomyces cerevisiae for improved industrially important properties. FEMS Yeast Res. 2012, 12, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Vanee, N.; Fisher, A.B.; Fong, S.S. Evolutionary Engineering for Industrial Microbiology. In Reprogramming Microbial Metabolic Pathways. Subcellular Biochemistry; Wang, X., Chen, J., Quinn, P., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 64. [Google Scholar]
- Fong, S.S. Evolutionary engineering of industrially important microbial phenotypes. In The Metabolic Pathway Engineering Handbook: Tools and Applications; Smolke, C.D., Ed.; CRC Press: New York, NY, USA, 2010; ISBN 978-142-0077-65-0. [Google Scholar]
- Sonderegger, M.; Sauer, U. Evolutionary engineering of Saccharomyces cerevisiae for anaerobic growth on xylose. Appl. Environ. Microbiol. 2003, 69, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- López-Malo, M.; García-Rios, E.; Melgar, B.; Sanchez, M.R.; Dunham, M.J.; Guillamón, J.M. Evolutionary engineering of a wine yeast strain revealed a key role of inositol and mannoprotein metabolism during low-temperature fermentation. BMC Genom. 2015, 16, 537. [Google Scholar] [CrossRef] [PubMed]
- Office Internationale de la Vigne et du Vin. International Code of Oenological Practices; OIV: Paris, France, 2010. [Google Scholar]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Alcohols, and other volatile compounds. The chemistry of wine stabilization and treatments. In Handbook of Enology, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006; Volume 2, pp. 51–64. [Google Scholar] [CrossRef]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Silva, R.F.; Chaves, S.R.; Sousa, M.J.; Côrte-Real, M. The impact of acetate metabolism on yeast fermentative performance and wine quality: Reduction of volatile acidity of grape-musts and wines—Minireview. Appl. Microbiol. Biotechnol. 2011, 89, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosme, F.; Vilela, A.; Filipe-Ribeiro, L.; Inês, A.; Nunes, F.-M. Wine microbial spoilage: Advances in defects remediation. In Microbial Contamination and Food Degradation, Handbook of Bioengineering, 1st ed.; Grumezescu, A., Holban, A.M., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: New York, NY, USA, 2017; Volume 10, pp. 271–314. Available online: https://www.elsevier.com/books/microbial-contamination-and-food-degradation/grumezescu/978-0-12-811262-5 (accessed on 20 May 2018).
- Hill, A. Traditional methods of detection and identification of brewery spoilage organisms. In Brewing Microbiology: Managing Microbes, Ensuring Quality and Valorising Waste; Series in Food Science, Technology and Nutrition; Woodhead: London, UK, 2015; Volume 289, 506p. [Google Scholar]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Quality and Composition of Airén Wines Fermented by Sequential Inoculation of Lachancea thermotolerans and Saccharomyces cerevisiae. Food Technol. Biotechnol. 2016, 54, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; House, J.F.; Joseph, C.M.L.; Bisson, L.F.; Bamforth, C.W. Lachancea thermotolerans as an alternative yeast for the production of beer. J. Inst. Brew. 2016, 122, 599–604. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The Combined Use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the Anthocyanin Wine Composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, M.W.; Rodriguez, S.B.; Honey, N.K.; Thornton, R.J. Purification, and characterization of urease from Schizosaccharomyces pombe. Can. J. Microbiol. 1996, 42, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Reduction of volatile acidity of wines by selected yeast strains. Appl. Microbiol. Biotechnol. 2008, 80, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela-Moura, A.; Schuller, D.; Falco, V.; Mendes-Faia, A.; Côrte-Real, M. Effect of refermentation conditions and micro-oxygenation on the reduction of volatile acidity by commercial S. cerevisiae strains and their impact on the aromatic profile of wines. Int. J. Food Microbiol. 2010, 141, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Effects of acetic acid, ethanol and SO2 on the removal of volatile acidity from acidic wines by two Saccharomyces cerevisiae commercial strains. Appl. Microbiol. Biotechnol. 2010, 87, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüller, H.J. Transcriptional control of non-fermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 2003, 43, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.J.; Rodrigues, F.; Côrte-Real, M.; Leão, C. Mechanisms underlying the transport and intracellular metabolism of acetic acid in the presence of glucose in the yeast Zygosaccharomyces bailii. Microbiology 1998, 144, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A.; Amaral, C.; Schuller, D.; Mendes-Faia, A.; Corte-Real, M. Combined use of Wallerstein and Zygosaccharomyces bailii modified differential media to isolate yeasts for the controlled reduction of volatile acidity of grape musts and wines. J. Biotech Res. 2015, 6, 43–53. [Google Scholar]
- Schuller, D.; Côrte-Real, M.; Leão, C. A differential medium for the enumeration of the spoilage yeast Zygosaccharomyces bailii in wine. J. Food Prot. 2000, 63, 1570–1575. [Google Scholar] [CrossRef] [PubMed]
- Van Uden, N. Transport-limited fermentation, and growth of Saccharomyces cerevisiae and its competitive inhibition. Arch. Mikrobiol. 1967, 58, 155–168. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Glucose (13%, w/v) Ethanol (4%, v/v) | Glucose (3.3%, w/v) Ethanol (10%, v/v) | |||
|---|---|---|---|---|
| Aerobic Conditions | Limited Aerobic Conditions | Aerobic Conditions | Limited Aerobic Conditions | |
| Yeast strains | Acetic acid Glucose | Acetic acid Glucose | Acetic acid Glucose | Acetic acid Glucose |
| Z. bailii ISA 1307 | 94.8 ± 3.30 c 52.4 ± 2.62 c | 40.9 ± 9.80 a 38.8 ± 6.36 b | 71.2 ± 3.02 b 23.1 ± 5.60 a | 41.6 ± 2.64 a 39.4 ± 2.10 b |
| L. thermotolerans 44C | 94.6 ± 4.79 d 58.5 ± 8.60 c | 15.25 ± 3.30 a 31.0 ± 5.69 b | 28.1 ± 1.70 c 16.4 ± 1.76 a | 17.4 ± 7.16 b 30.4 ± 5.79 b |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation 2018, 4, 56. https://doi.org/10.3390/fermentation4030056
Vilela A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation. 2018; 4(3):56. https://doi.org/10.3390/fermentation4030056
Chicago/Turabian StyleVilela, Alice. 2018. "Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines" Fermentation 4, no. 3: 56. https://doi.org/10.3390/fermentation4030056
APA StyleVilela, A. (2018). Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation, 4(3), 56. https://doi.org/10.3390/fermentation4030056