Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Impact of YNB Concentration on β-Carotene Production of SM14
2.2. Impact of Water Sources on β-Carotene Production of SM14
2.3. Impact of NaCl on β-Carotene Production of SM14
2.4. Impact of pH on β-Carotene Production of SM14
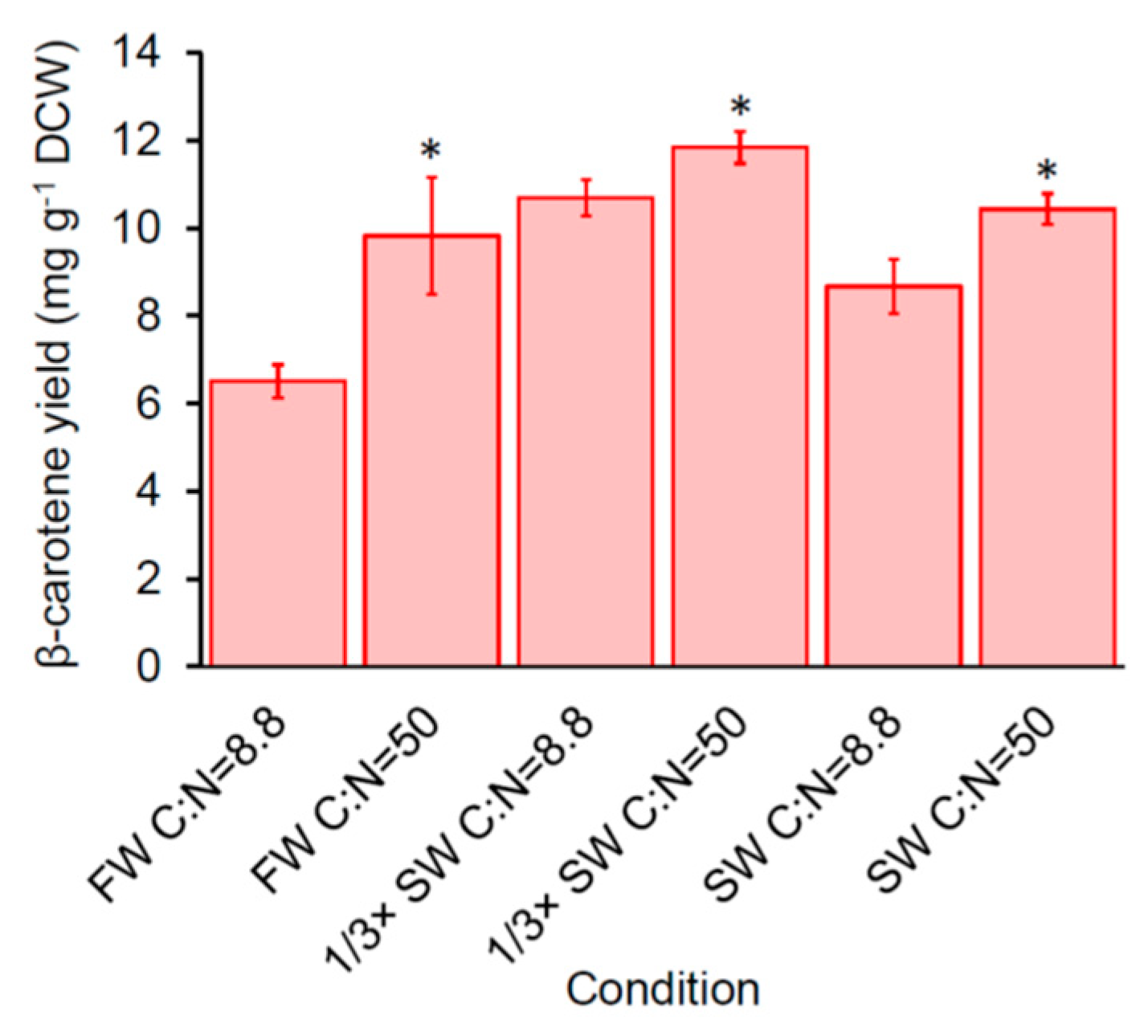
2.5. Impact of Carbon-to-Nitrogen Ratio (C:N) on β-Carotene Production of SM14
2.6. Lipid Production and Composition
2.7. The Impact of Seawater on β-Carotene Production of Other Evolved Hyper-Producers
3. Conclusions
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Carotenoid Quantification
4.3. Lipid Quantification
Supplementary Materials
 ) and seawater (
) and seawater (  ) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, (
) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, (  ) initial pH 5.2, (
) initial pH 5.2, (  ) initial pH 6.5, (
) initial pH 6.5, (  ) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) ( ) C:N = 8.8 in freshwater, (
) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) ( ) C:N = 8.8 in freshwater, (  ) C:N = 50 in freshwater, (
) C:N = 50 in freshwater, (  ) C:N = 8.8 in freshwater with 0.05 M NaCl, ( ) C:N = 50 in freshwater with 0.05 M NaCl; (b) ( ) C:N = 8.8 in 1/3× seawater, ( ) C:N = 50 in 1/3× seawater, ( ) C:N = 8.8 in seawater, ( ) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater ( ) or 1/3× seawater ( ). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid composition
) C:N = 8.8 in freshwater with 0.05 M NaCl, ( ) C:N = 50 in freshwater with 0.05 M NaCl; (b) ( ) C:N = 8.8 in 1/3× seawater, ( ) C:N = 50 in 1/3× seawater, ( ) C:N = 8.8 in seawater, ( ) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater ( ) or 1/3× seawater ( ). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid compositionAuthor Contributions
Funding
Conflicts of Interest
References
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Fong, N.J.; Burgess, M.L.; Barrow, K.D.; Glenn, D.R. Carotenoid accumulation in the psychrotrophic bacterium Arthrobacter agilis in response to thermal and salt stress. Appl. Microbiol. Biotechnol. 2001, 56, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Distributions, biosyntheses and functions of carotenoids in algae. Agro Food Ind. Hi Tech 2013, 24, 55–58. [Google Scholar]
- Avalos, J.; Limon, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Kirti, K.; Amita, S.; Priti, S.; Mukesh Kumar, A.; Jyoti, S. Colorful world of microbes: Carotenoids and their applications. Adv. Biol. 2014, 2014, 13. [Google Scholar] [CrossRef]
- Nigam, P.S.; Luke, J.S. Food additives: Production of microbial pigments and their antioxidant properties. Curr. Opin. Food Sci. 2016, 7, 93–100. [Google Scholar] [CrossRef]
- Manimala, M.; Murugesan, R. In vitro antioxidant and antimicrobial activity of carotenoid pigment extracted from Sporobolomyces sp. Isolated from natural source. J. Appl. Nat. Sci. 2014, 6, 649–653. [Google Scholar] [CrossRef]
- Anunciato, T.P.; da Rocha, P.A. Carotenoids and polyphenols in nutricosmetics, nutraceuticals, and cosmeceuticals. J. Cosmet. Dermatol. 2012, 11, 51–54. [Google Scholar] [CrossRef]
- Vachali, P.; Bhosale, P.; Bernstein, P.S. Microbial carotenoids. In Microbial Carotenoids from Fungi: Methods and Protocols; Barredo, J.-L., Ed.; Humana Press: Totowa, NJ, USA, 2012; pp. 41–59. [Google Scholar]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving beta-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef]
- Reyes, L.H.; Gomez, J.M.; Kao, K.C. Improving carotenoids production in yeast via adaptive laboratory evolution. Metab. Eng. 2014, 21, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kao, K.C.; Sherlock, G. Molecular characterization of clonal interference during adaptive evolution in asexual populations of Saccharomyces cerevisiae. Nat. Genet. 2008, 40, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.L.; Johnson, J.; Carswell, W.F.; Reyes, L.H.; Senger, R.S.; Kao, K.C. Characterization of an evolved carotenoids hyper-producer of Saccharomyces cerevisiae through bioreactor parameter optimization and raman spectroscopy. J. Ind. Microbiol. Biotechnol. 2016, 43, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Gerbens-Leenes, P.W.; Lienden, A.R.V.; Hoekstra, A.Y.; van der Meer, T.H. Biofuel scenarios in a water perspective: The global blue and green water footprint of road transport in 2030. Glob. Environ. Chang. 2012, 22, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Zaky, A.S. Marine fermentation, the sustainable approach for bioethanol production. EC Microbiol. 2017, ECO.01, 25–27. [Google Scholar]
- Goncalves, F.A.; dos Santos, E.S.; de Macedo, G.R. Alcoholic fermentation of Saccharomyces cerevisiae, Pichia stipitis and Zymomonas mobilis in the presence of inhibitory compounds and seawater. J. Basic Microbiol. 2015, 55, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Gadre, R.V. Production of beta-carotene by a Rhodotorula glutinis mutant in sea water medium. Bioresour. Technol. 2001, 76, 53–55. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Breierova, E.; Koci, R. Production of carotenoid-/ergosterol-supplemented biomass by red yeast Rhodotorula glutinis grown under external stress. Food Technol. Biotechnol. 2010, 48, 56–61. [Google Scholar]
- Farhat, N.; Rabhi, M.; Falleh, H.; Jouini, J.; Abdelly, C.; Smaoui, A. Optimization of salt concentrations for a higher carotenoid production in Dunaliella salina (chlorophyceae). J. Phycol. 2011, 47, 1072–1077. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Certik, M.; Dvorakova, T.; Haronikova, A. Use of several waste substrates for carotenoid-rich yeast biomass production. J. Environ. Manag. 2012, 95, S338–S342. [Google Scholar] [CrossRef]
- Kanzy, H.M.; Nasr, N.; El-Shazly, H.A.; Barakat, O.S. Optimization of carotenoids production by yeast strains of Rhodotorula using salted cheese whey. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 456–469. [Google Scholar]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachaal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef]
- Kuge, S.; Jones, N. YAP1 dependent activation of TRX2 is essential for the response of Saccharomyces cerevisiae to oxidative stress by hydroperoxides. Embo J. 1994, 13, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Godon, C.; Lagniel, G.; Spector, D.; Garin, J.; Labarre, J.; Toledano, M.B. Yap1 and skn7 control two specialized oxidative stress response regulons in yeast. J. Biol. Chem. 1999, 274, 16040–16046. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.; Ideker, T. Genome-wide fitness and expression profiling implicate Mga2 in adaptation to hydrogen peroxide. PLoS Genet. 2009, 5, e1000488. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Leroy, C.; Isnard, A.D.; Labarre, J.; Boy-Marcotte, E.; Toledano, M.B. The control of the yeast H2O2 response by the Msn2/4 transcription factors. Mol. Microbiol. 2002, 45, 233–241. [Google Scholar] [CrossRef]
- Berry, D.B.; Gasch, A.P. Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol. Biol. Cell 2008, 19, 4580–4587. [Google Scholar] [CrossRef]
- Święciło, A. Cross-stress resistance in Saccharomyces cerevisiae yeast—New insight into an old phenomenon. Cell Stress Chaperones 2016, 21, 1–14. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Sestric, R.; Ignatia, L.; Levin, D.; German, J.B.; Gillies, L.A.; Almada, L.A.; Boundy-Mills, K.L. Manipulation of culture conditions alters lipid content and fatty acid profiles of a wide variety of known and new oleaginous yeast species. Bioresour. Technol. 2013, 144, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Turk, M.; Mejanelle, L.; Sentjurc, M.; Grimalt, J.O.; Gunde-Cimerman, N.; Plemenitas, A. Salt-induced changes in lipid composition and membrane fluidity of halophilic yeast-like melanized fungi. Extremophiles 2004, 8, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lv, X.; Ye, L.; Zhou, P.; Yu, H. Construction of lycopene-overproducing Saccharomyces cerevisiae by combining directed evolution and metabolic engineering. Metab. Eng. 2015, 30, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Qin, X.; Cheng, Y.; Laskar, D.; Qiao, W.; Sun, S.; Reyes, L.H.; Wang, X.; Dai, S.Y.; Sattler, S.E.; et al. Simultaneous conversion of all cell wall components by an oleaginous fungus without chemi-physical pretreatment. Green Chem. 2015, 17, 1657–1667. [Google Scholar] [CrossRef] [Green Version]
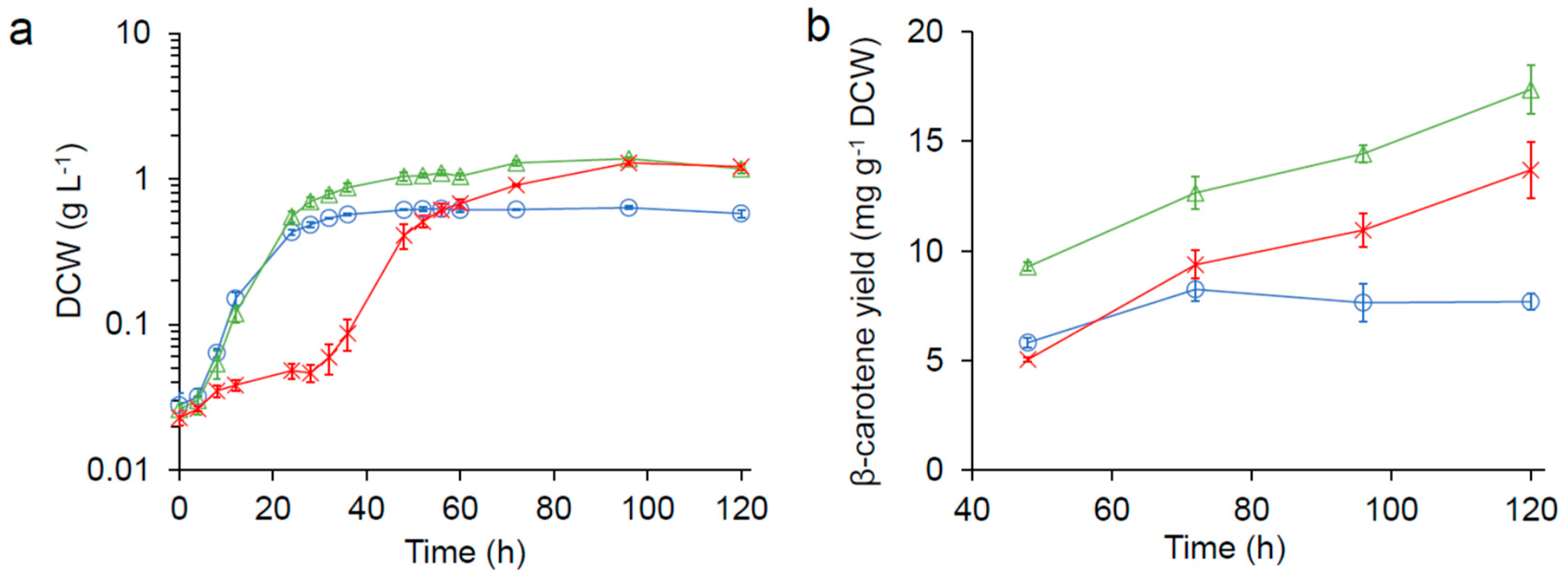
), 1/3× seawater ( ), and seawater (  ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
), 1/3× seawater ( ), and seawater ( ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
), 1/3× seawater ( ), and seawater ( ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
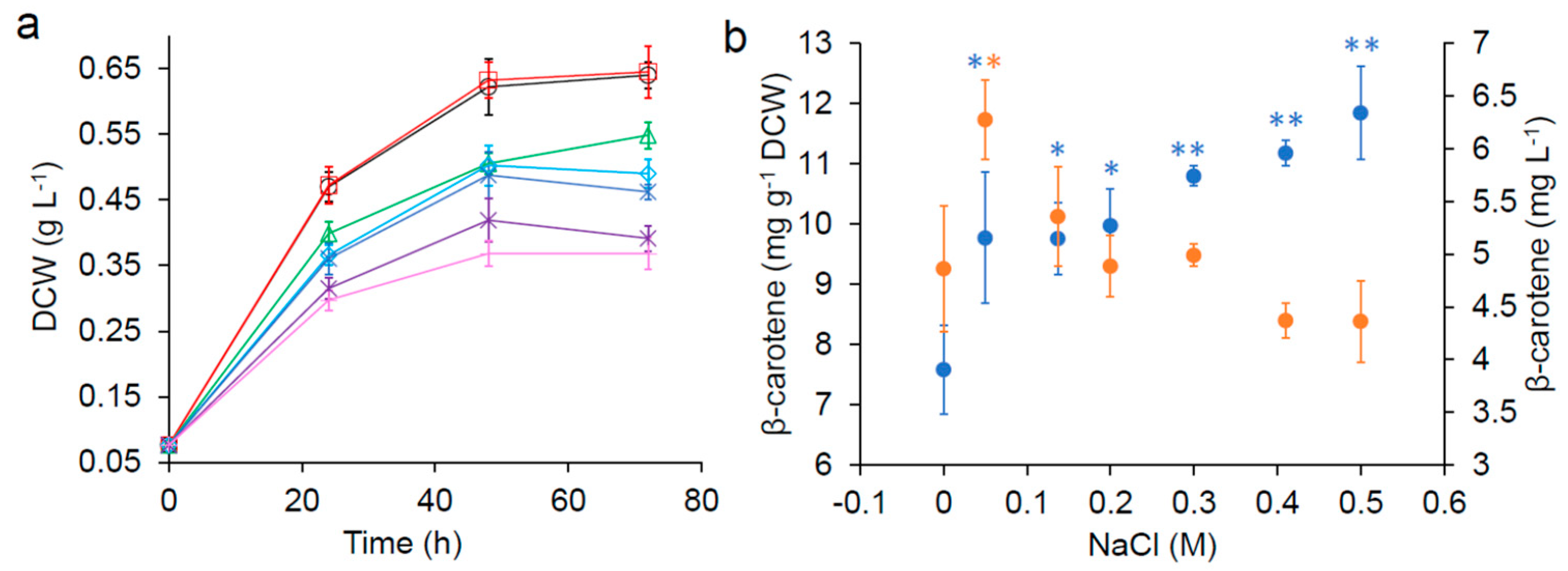
 ), 0.05 M ( ), 0.137 M ( ), 0.2 M (
), 0.05 M ( ), 0.137 M ( ), 0.2 M (  ), 0.3 M (
), 0.3 M (  ), 0.41 M (
), 0.41 M (  ) and 0.5 M (
) and 0.5 M (  ) NaCl. (b) β-carotene yield (
) NaCl. (b) β-carotene yield (  ) and concentration (
) and concentration (  ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
), 0.05 M ( ), 0.137 M ( ), 0.2 M ( ), 0.3 M ( ), 0.41 M ( ) and 0.5 M ( ) NaCl. (b) β-carotene yield ( ) and concentration ( ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
), 0.05 M ( ), 0.137 M ( ), 0.2 M ( ), 0.3 M ( ), 0.41 M ( ) and 0.5 M ( ) NaCl. (b) β-carotene yield ( ) and concentration ( ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
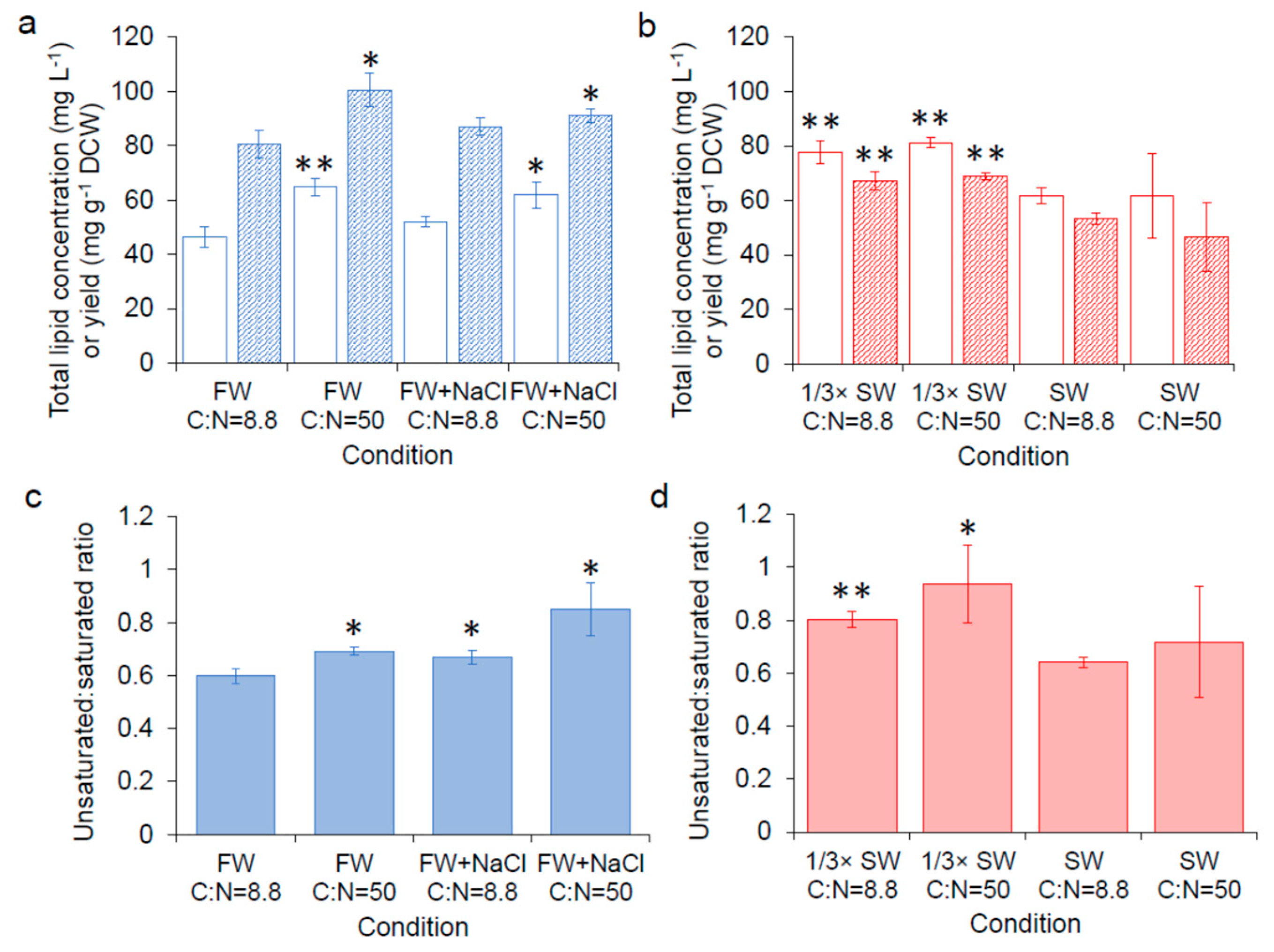
 ) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Xie, S.; Yuan, J.S.; Kao, K.C. Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae. Fermentation 2019, 5, 6. https://doi.org/10.3390/fermentation5010006
Guo Y, Xie S, Yuan JS, Kao KC. Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae. Fermentation. 2019; 5(1):6. https://doi.org/10.3390/fermentation5010006
Chicago/Turabian StyleGuo, Yuqi, Shangxian Xie, Joshua S. Yuan, and Katy C. Kao. 2019. "Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae" Fermentation 5, no. 1: 6. https://doi.org/10.3390/fermentation5010006
APA StyleGuo, Y., Xie, S., Yuan, J. S., & Kao, K. C. (2019). Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae. Fermentation, 5(1), 6. https://doi.org/10.3390/fermentation5010006