Production of Proteins prM/M and E of Dengue Virus-3 in Pichia pastoris: Simplified Purification and Evaluation of Their Use as Antigens in Serological Diagnosis of Dengue

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Plasmid
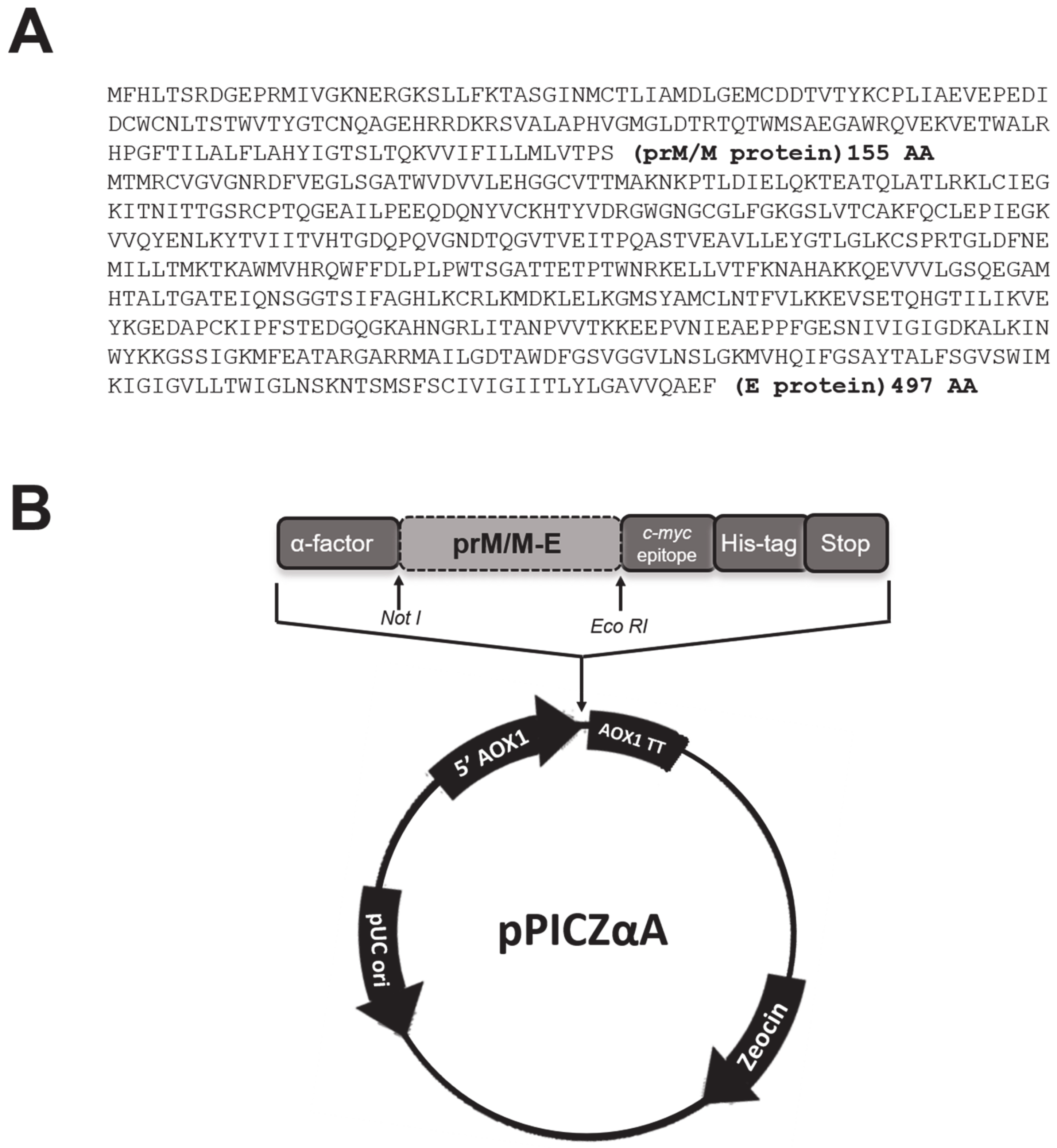
2.2. Cloning the Sequences of the prM/M and E Proteins in the Expression Vector pPICZαA
2.3. Expression of Recombinant Proteins
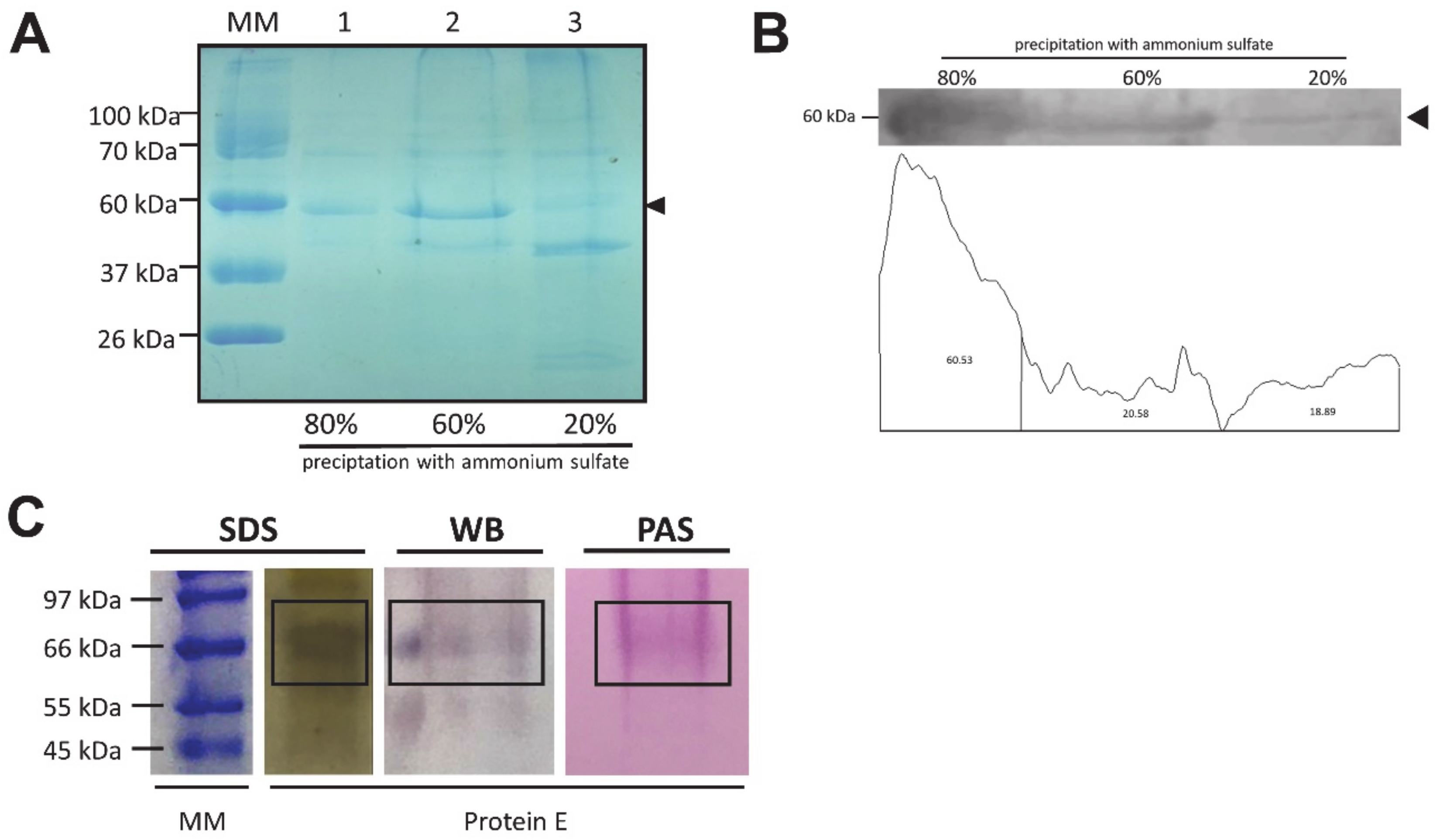
2.4. Recombinant Protein Characterization
2.5. Samples of Sera
2.6. Indirect Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of IgM and IgG
2.7. Statistical Analysis
3. Results
3.1. Construction of Recombinant Pichia pastoris Clones Expressing the prM and E Proteins of DENV3
3.2. Expression of Recombinant Proteins
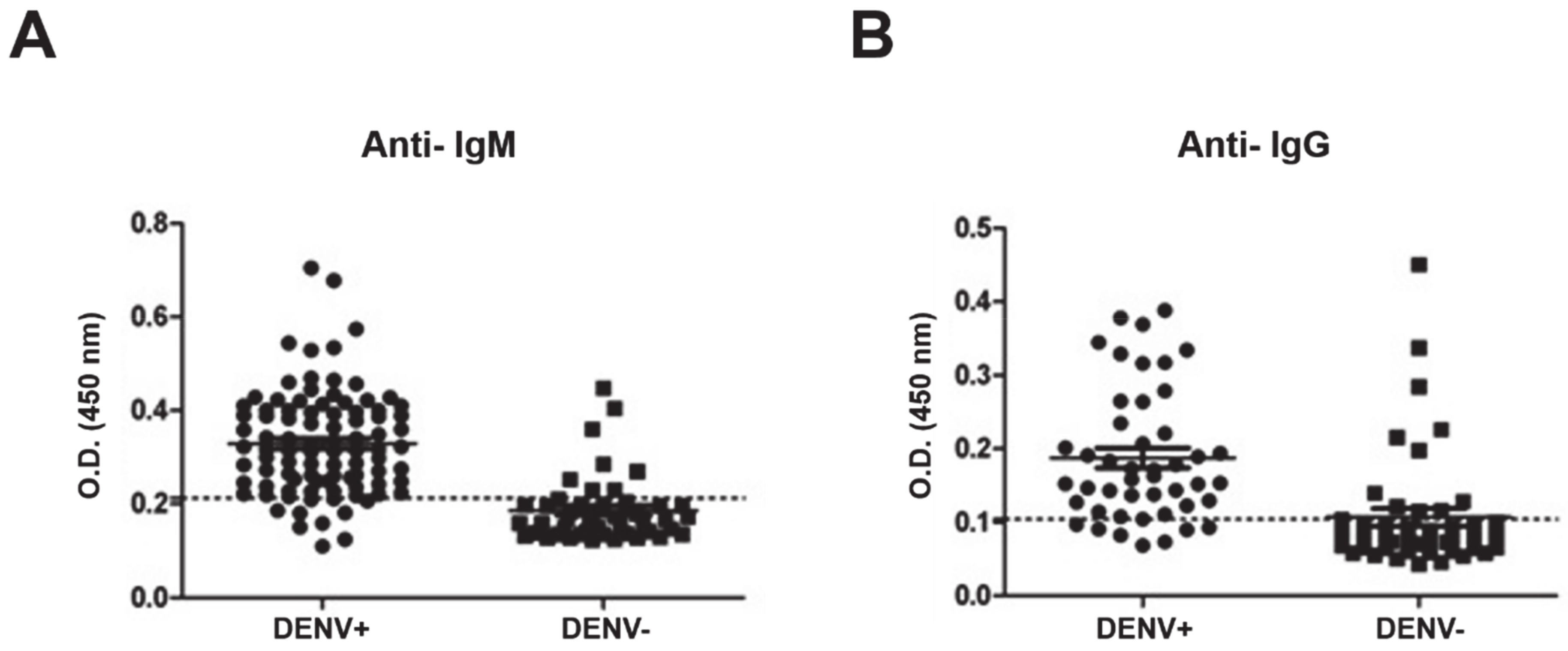
3.3. ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Dengue and Severe Dengue. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 10 September 2020).
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Gubler, D.J.; Kuno, G.; Markoff, L. Flaviviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer and Lippincott Williams: Philadelphia, PA, USA, 2007; pp. 1153–1252. [Google Scholar]
- Perera, R.; Kuhn, R.J. Structural proteomics of dengue virus. Curr. Opin. Microbiol. 2008, 11, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, S.S.; Blaney, J.E.; Durbin, A.P.; Murphy, B.R. Prospects for a dengue virus vaccine. Nat. Rev. Genet. 2007, 5, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martinez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Genet. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, F.X.; Stiasny, K. Flaviviruses and flavivirus vaccines. Vaccine 2012, 30, 4301–4306. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.T.T.; Bhuvanakantham, R.; Li, J.; Howe, J.; Ng, M.-L. Tyrosine 78 of premembrane protein is essential for assembly of West Nile virus. J. Gen. Virol. 2009, 90, 1081–1092. [Google Scholar] [CrossRef]
- Zheng, A.; Umashankar, M.; Kielian, M. In Vitro and In Vivo Studies Identify Important Features of Dengue Virus pr-E Protein Interactions. PLoS Pathog. 2010, 6, e1001157. [Google Scholar] [CrossRef]
- Rey, F.A. Dengue virus envelope glycoprotein structure: New insight into its interactions during viral entry. Proc. Natl. Acad. Sci. USA 2003, 100, 6899–6901. [Google Scholar] [CrossRef] [Green Version]
- De Paula, S.O.; Fonseca, B.A.L. Dengue: A review of the laboratory tests a clinican must know to achieve a correct diagnosis. Braz. J. Infect. Dis. 2004, 8, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Le, R.D.; Melanson, S.E.F. What’s New in Point-of-Care Testing? Point Care J. Near-Patient Test. Technol. 2014, 13, 152–155. [Google Scholar] [CrossRef]
- Holmes, D.A.; Purdy, D.E.; Chao, D.-Y.; Noga, A.J.; Chang, G.-J.J. Comparative Analysis of Immunoglobulin M (IgM) Capture Enzyme-Linked Immunosorbent Assay Using Virus-Like Particles or Virus-Infected Mouse Brain Antigens To Detect IgM Antibody in Sera from Patients with Evident Flaviviral Infections. J. Clin. Microbiol. 2005, 43, 3227–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paula, S.O.; Dutra, N.R.; De Paula, M.B.; De Oliveira, M.D.; De Oliveira, L.L. The laboratorial diagnosis of dengue: Applications and implications. J. Glob. Infect. Dis. 2009, 1, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Batra, G.; Gurramkonda, C.; Nemani, S.K.; Jain, S.K.; Swaminathan, S.; Khanna, N. Optimization of conditions for secretion of dengue virus type 2 envelope domain III using Pichia pastoris. J. Biosci. Bioeng. 2010, 110, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jiang, H.; Zhou, J.; Yang, X.; Tang, Y.; Fang, D.; Jiang, L. Recombinant dengue virus-like particles from Pichiapastoris: Efficient production and immunological properties. Virus Genes 2010, 40, 53–59. [Google Scholar] [CrossRef]
- Rocha, E.S.O.; Oliveira, J.G.; Santos, J.R.; Rodrigues, G.O.L.; Figueiredo, L.B.; Pessanha, E.M.; Proietti, F.A.; Fonseca, F.G.; Bonjardim, C.A.; Ferreira, P.C.P.; et al. Recombinant envelope protein-based enzyme immunoassay for IgG antibodies is comparable to netralization tests for epidemiological studies of dengue infection. J. Virol. Methods 2013, 187, 114–120. [Google Scholar] [CrossRef]
- Álvarez-Rodríguez, L.M.; Ramos-Ligonio, A.; Rosales-Encina, J.L.; Martínez-Cázares, M.T.; Parissi-Crivelli, A.; López-Monteoni, A. Expression, Purification, and evaluation of Diagnostic Potential and Immunogenicity of Recombinant NS3 Protein from All Serotypes of Degue Virus. J. Trop. Med. 2012. [Google Scholar] [CrossRef]
- Cardoso, S.A.; Paixão, V.F.; Oliveira, M.D.; Honda, E.R.; Oliveira, L.L.; da Silva, C.C.; De Paula, S.O. Dengue-1 envelope protein domain III produced in Pichiapastoris: potential use for serological diagnosis. Protein Expr. Purif. 2013, 92, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Daly, R.; Hearn, M.T.W. Expression of heterologous proteins in Pichiapastoris: a useful experimental tool in protein engineering and production. J. Mol. Recognit. 2005, 18, 119–138. [Google Scholar] [CrossRef]
- Krainer, F.W.; Dietzsch, C.; Hajek, T.; Herwig, C.; Spadiut, O.; Glieder, A. Recombinant protein expression in Pichiapastoris strains with an engineered methanol utilization pathway. Microb. Cell Factories 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J. Dengue and Dengue Hemorrhagic Fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [Green Version]
- Bavia, L.; Melanda, F.N.; Arruda, T.B.; Mosimann, A.L.P.; Silveira, G.F.; Aoki, M.N.; Kuczera, D.; Sarzi, M.L., Jr.; Conchon-Costa, I.; Pavanelli, W.R.; et al. Epidemiological study on dengue in southern Brazil under the perspective of climate and poverty. Sci. Rep. 2020, 10, 2127. [Google Scholar] [CrossRef] [PubMed]
- Sugrue, R.J.; Cui, T.; Xu, Q.; Fu, J.; Chan, Y.C. The production of recombinant dengue vírus E protein using Escherichia coli and Pichiapastoris. J. Virol. Methods 1997, 69, 159–169. [Google Scholar] [CrossRef]
- Blacksell, S.D.; Jarman, R.G.; Gibbons, R.V.; Tanganuchitcharnchai, A.; Mammen, M.P.; Nisalak, A.; Kalayanarooj, S.; Bailey, M.; Premaratna, R.; De Silva, H.J.; et al. Comparison of Seven Commercial Antigen and Antibody Enzyme-Linked Immunosorbent Assays for Detection of Acute Dengue Infection. Clin. Vaccine Immunol. 2012, 19, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Cuzzubbo, A.J.; Vaughn, D.W.; Nisalak, A.; Solomon, T.; Kalayanarooj, S.; Aaskov, J.; Dung, N.M.; Devine, P.L. Comparison of PanBio Dengue Duo Enzyme-Linked Immunosorbent Assay (ELISA) and MRL Dengue Fever Virus Immunoglobulin M Capture ELISA for Diagnosis of Dengue Virus Infections in Southeast Asia. Clin. Diagn. Lab. Immunol. 1999, 6, 705–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koraka, P.; Zeller, H.; Niedrig, M.; Osterhaus, A.D.; Groen, J. Reactivity of serum 473 samples from patients with a flavivirus infection measured by immunofluores- 474 cence assay and ELISA. Microbes Infect. 2002, 4, 1209–1215. [Google Scholar] [CrossRef]
- Fry, S.R.; Meyer, M.; Semple, M.G.; Simmons, C.P.; Sekaran, S.D.; Huang, J.X.; McElnea, C.; Huang, C.; Valks, A.; Cooper, M.A. The diagnostic sensivity of dengue rapid test assays is significantly enhanced by using a combined antigen and antibody testing approach. PLoS Negl. Trop. Dis. 2011, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Positive | Negative | Test | |
|---|---|---|---|
| Anti-IgM | 92 | 46 | Dengue Duo IgM and IgG ELISA Capture Kits + MAC-ELISA IgM |
| Anti-IgG | 46 | 46 | Dengue Duo IgM and IgG ELISA Capture Kits |
| Ammonium sulfate | Densitometry (Area) | [] µg/mL | Yield |
|---|---|---|---|
| 80% | 46,031 | 131.213 | 53.52 % |
| 60% | 36,070 | 102.7731 | 41.92 % |
| 20% | 3997 | 11.20171 | 4.56 % |
| % Sensitivity | % Specificity | Cut-Off | |
|---|---|---|---|
| Anti-IgM | 82.61 | 89.25 | 0.2138 |
| Anti-IgG | 76.09 | 84.78 | 0.1043 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, M.D.O.; Dias, R.S.; Prates, J.W.O.; Monteiro, J.M.C.; Xisto, M.F.; da Silva, C.C.; De Paula, S.O. Production of Proteins prM/M and E of Dengue Virus-3 in Pichia pastoris: Simplified Purification and Evaluation of Their Use as Antigens in Serological Diagnosis of Dengue. Fermentation 2020, 6, 88. https://doi.org/10.3390/fermentation6030088
Teixeira MDO, Dias RS, Prates JWO, Monteiro JMC, Xisto MF, da Silva CC, De Paula SO. Production of Proteins prM/M and E of Dengue Virus-3 in Pichia pastoris: Simplified Purification and Evaluation of Their Use as Antigens in Serological Diagnosis of Dengue. Fermentation. 2020; 6(3):88. https://doi.org/10.3390/fermentation6030088
Chicago/Turabian StyleTeixeira, Michelle D. O., Roberto S. Dias, John W. O. Prates, Juliana M. C. Monteiro, Mariana F. Xisto, Cynthia C. da Silva, and Sérgio O. De Paula. 2020. "Production of Proteins prM/M and E of Dengue Virus-3 in Pichia pastoris: Simplified Purification and Evaluation of Their Use as Antigens in Serological Diagnosis of Dengue" Fermentation 6, no. 3: 88. https://doi.org/10.3390/fermentation6030088
APA StyleTeixeira, M. D. O., Dias, R. S., Prates, J. W. O., Monteiro, J. M. C., Xisto, M. F., da Silva, C. C., & De Paula, S. O. (2020). Production of Proteins prM/M and E of Dengue Virus-3 in Pichia pastoris: Simplified Purification and Evaluation of Their Use as Antigens in Serological Diagnosis of Dengue. Fermentation, 6(3), 88. https://doi.org/10.3390/fermentation6030088