Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
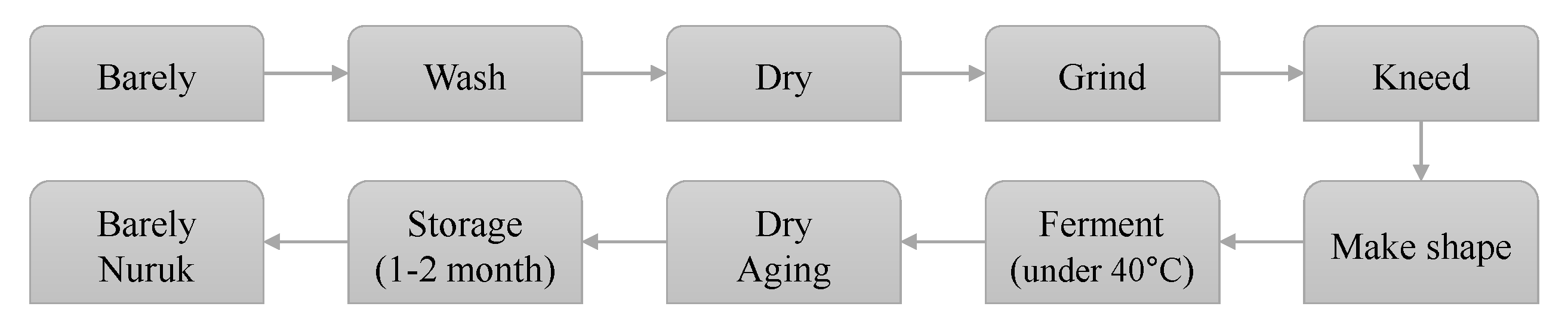
2.1. Preparation of Extract
2.2. Preparation of Ethyl Acetate (EtOAc) Fraction
2.3. PCR Amplification and Sequencing
2.4. Cell Culture
2.5. Measurement of Cell Viability
2.6. Measurement of Nitric Oxide Production
2.7. Measurement of Cytokine Production
2.8. Western Blot Assay
3. Results
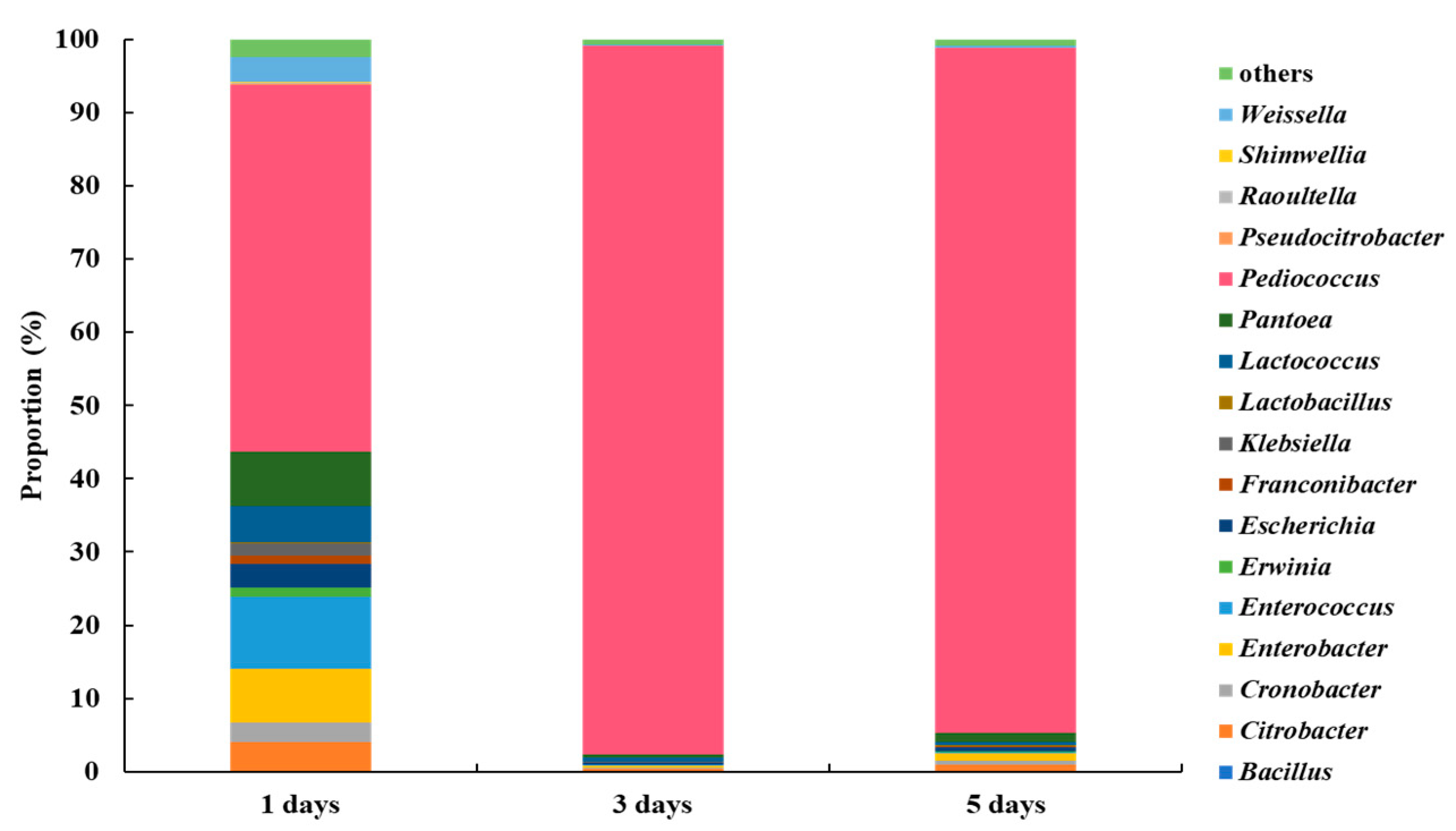
3.1. Microbial Community Analysis
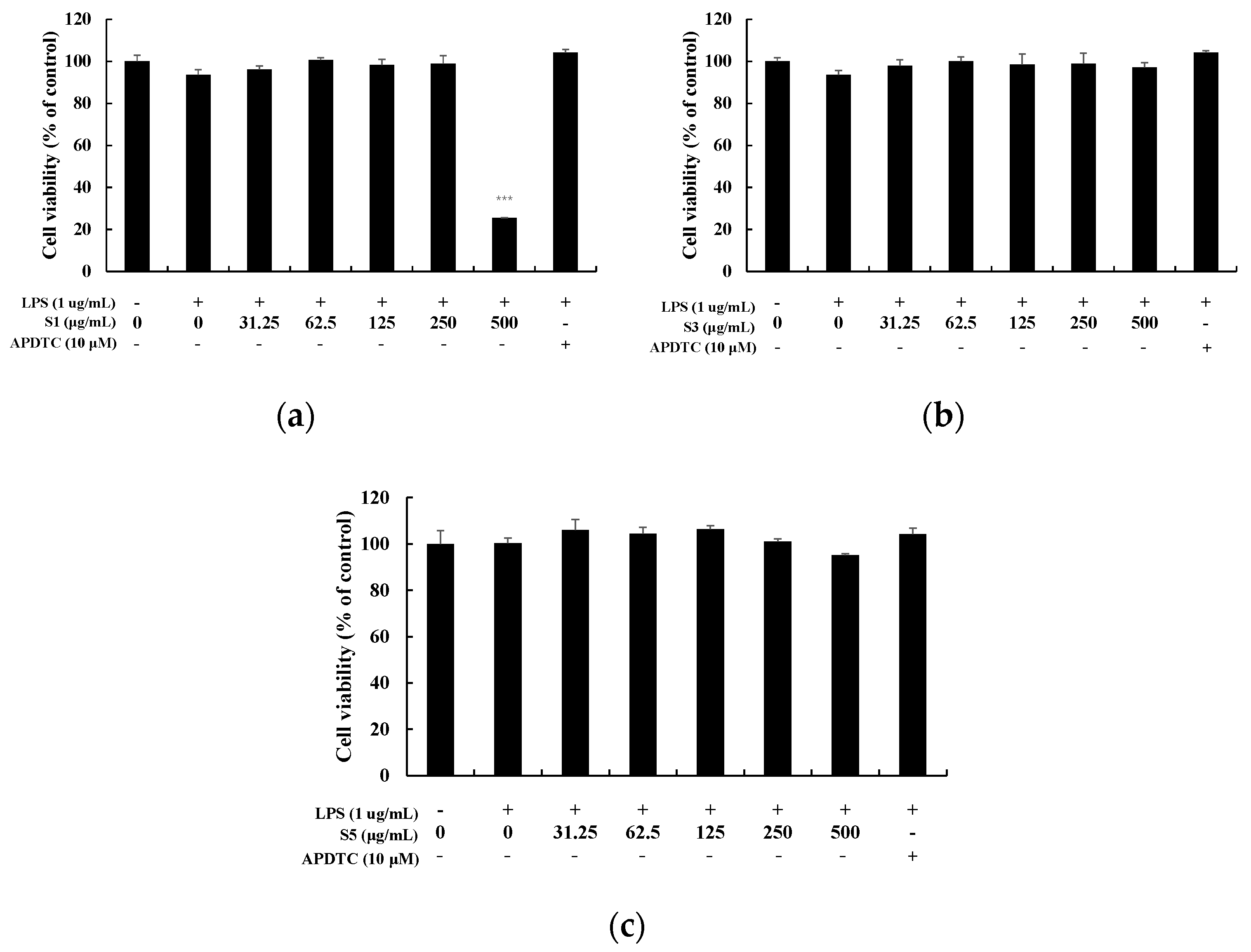
3.2. Measurement of Cell Viability
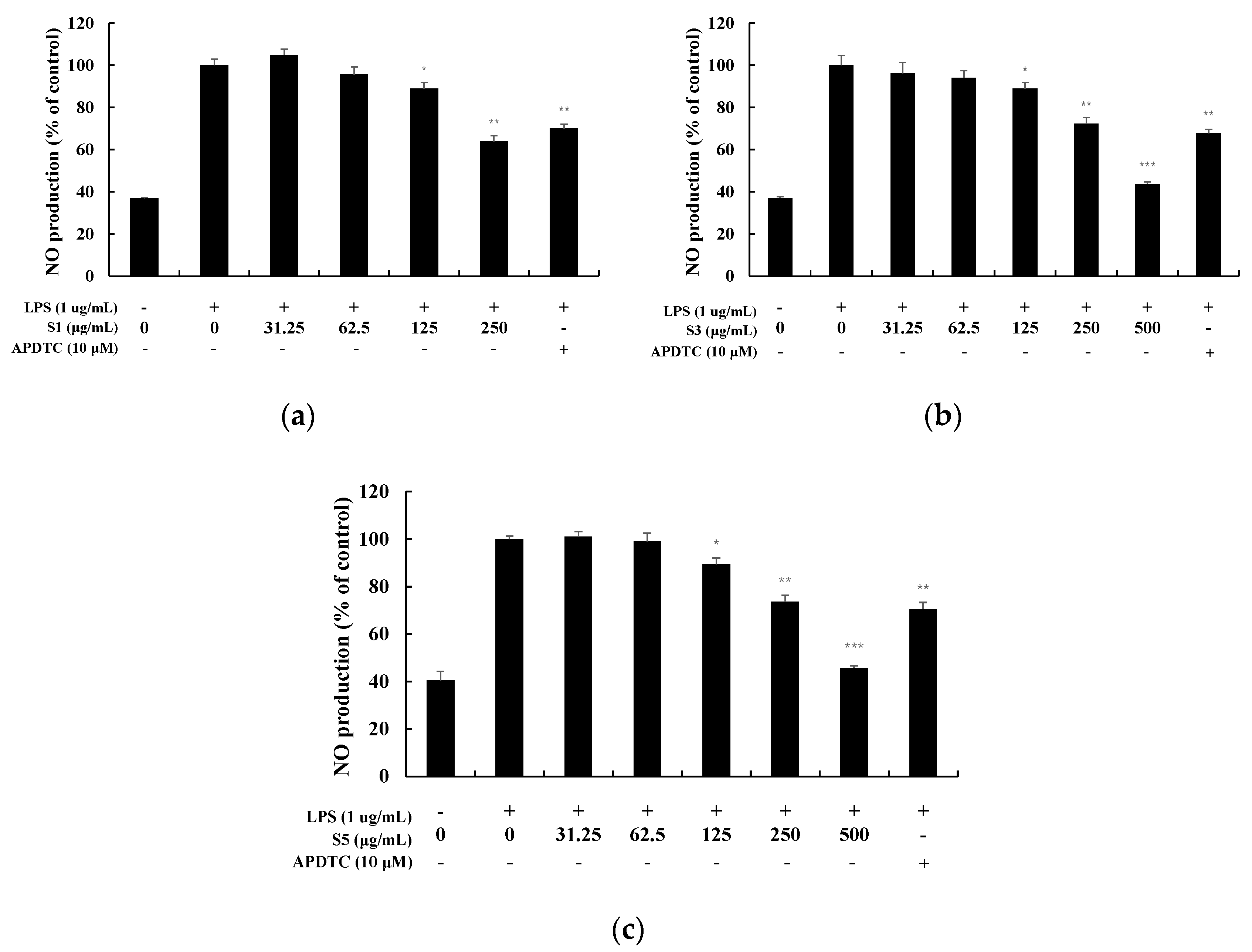
3.3. Measurement of Nitric Oxide Production
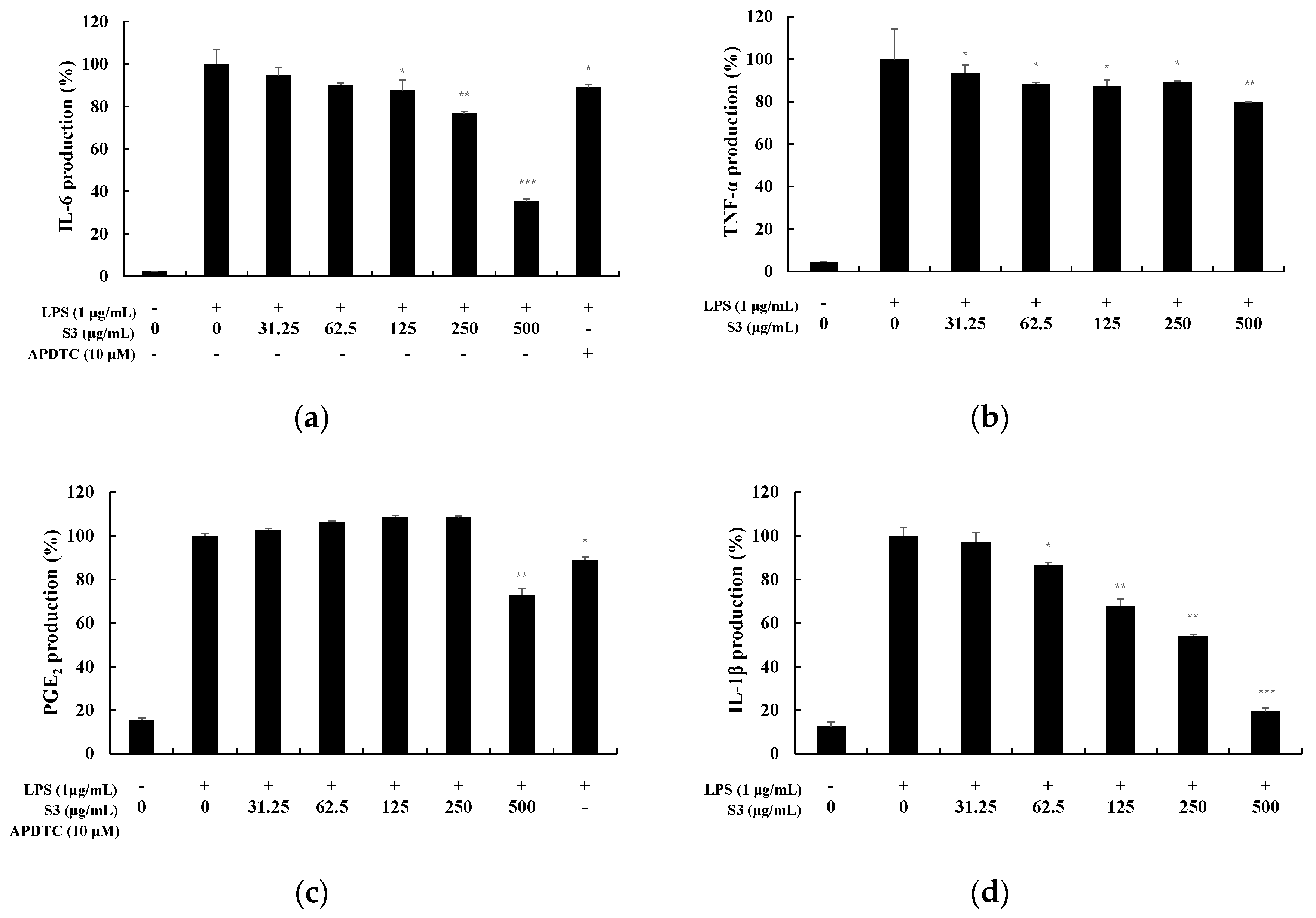
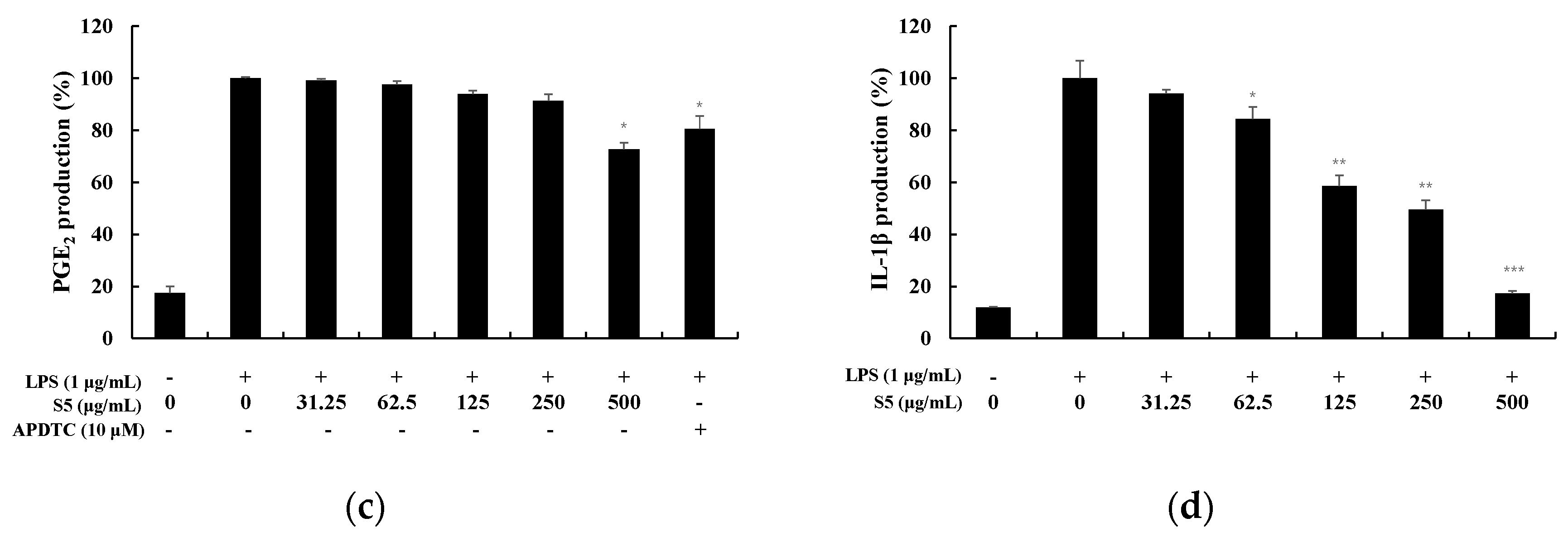
3.4. Measurement of Cytokine Production
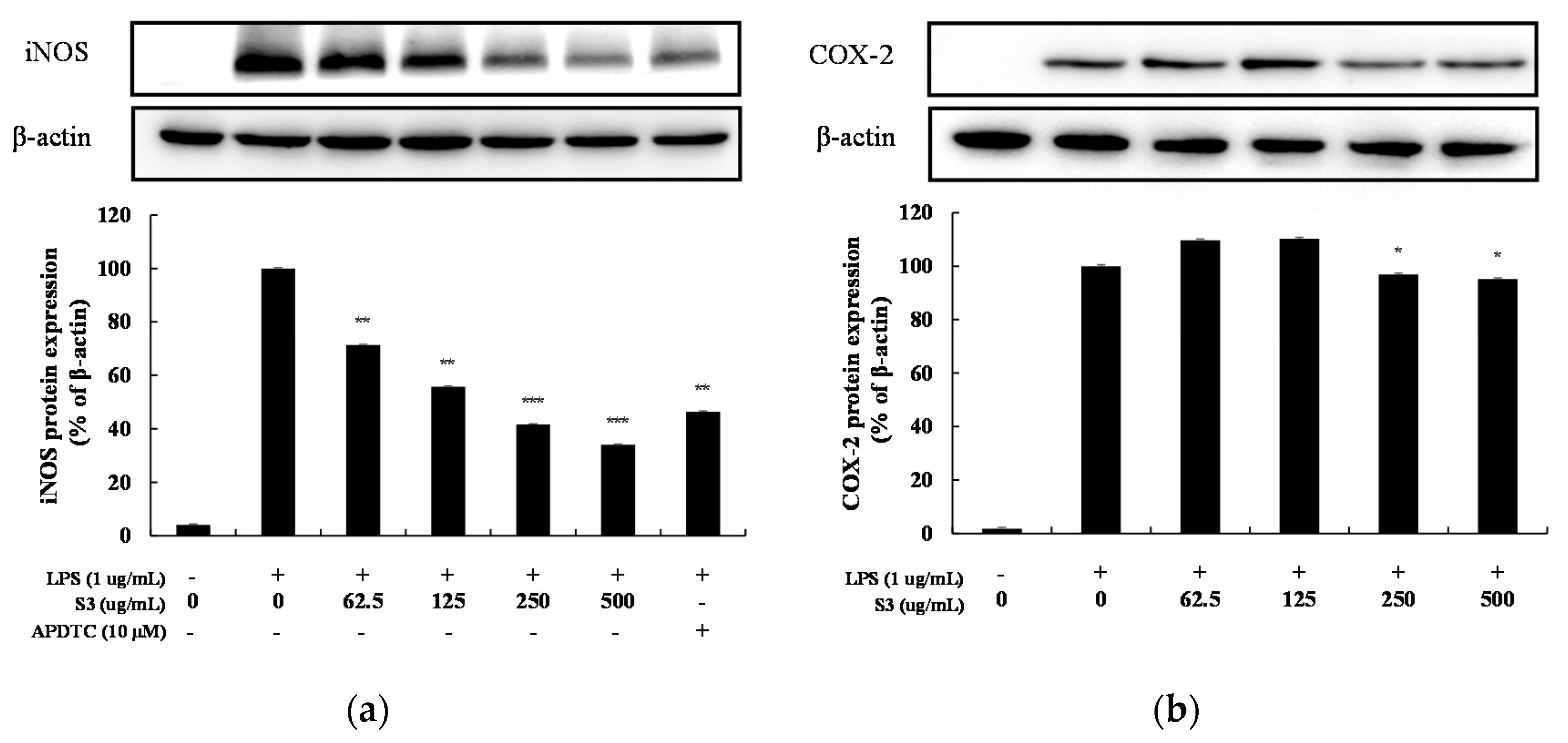
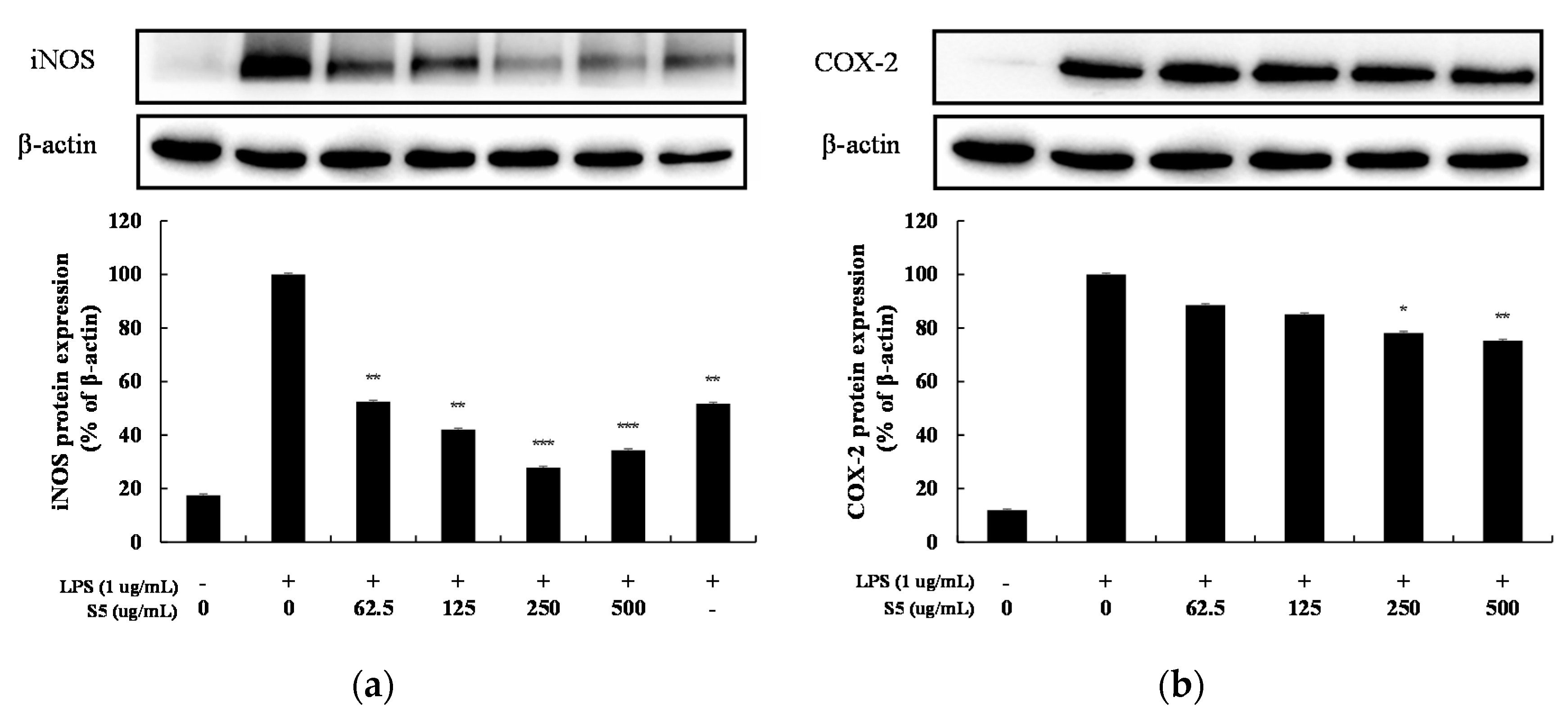
3.5. iNOS, COX-2 Expression in LPS-Induced RAW264.7
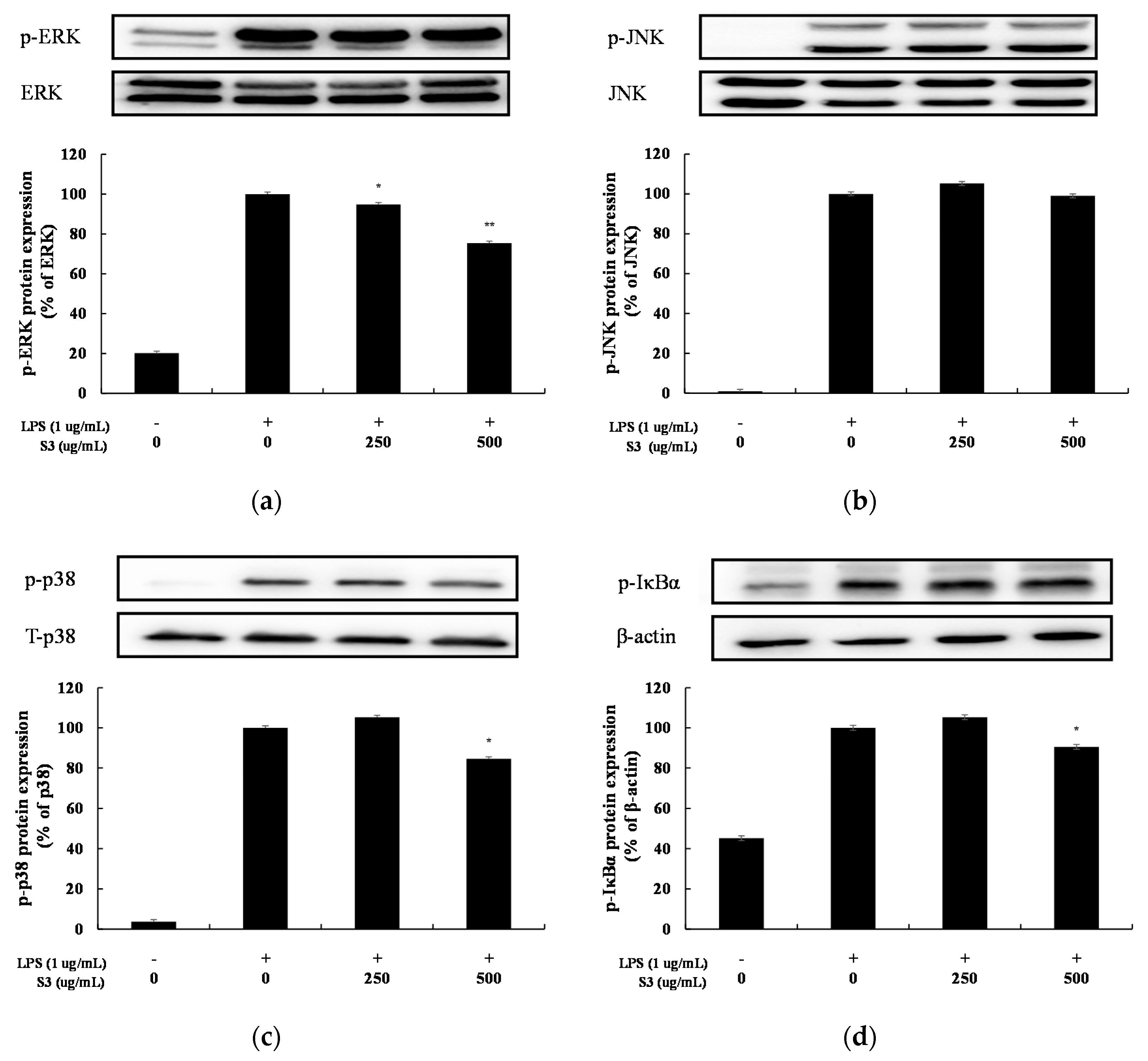
3.6. MAPK and NF-κB Signaling Activation in LPS-Induced RAW264.7
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J. Research of the Food Culture Comparison between the Tamra & Mongolia. Trans Humanit. 2012, 5, 211–243. [Google Scholar]
- Ahn, S.L.; Yoon, J.H. Development of Jeju Local Food Tour Package and Economic Value Estimation based on Contingent Valuation Method. J. East Asian Soc. Diet. Life 2016, 26, 346–358. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, E.J. Fermentation Characteristics of Shindari Added with Carrot. Korean J. Food Cook Sci. 2015, 3, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Applications of Next Generation Sequencing Technologies as Applied to Food-Related Microbiome Studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar] [CrossRef]
- Sarkar, D.; Fisher, P.B. Molecular Mechanisms of Aging-associated Inflammation. Cancer Lett. 2006, 236, 13–23. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kang, B.K.; Kim, K.B.W.R.; Kim, M.J.; Ahn, D.H. Anti-inflammatory Activity of Sargassum micracanthum Water Extract. J. Appl. Biol. Chem. 2014, 57, 227–234. [Google Scholar] [CrossRef]
- Choi, S.B.; Bae, G.S.; Jo, I.J.; Park, K.C.; Seo, S.H.; Kim, D.G.; Shin, J.Y.; Gwak, T.S.; Lee, J.H.; Lee, G.S.; et al. The Anti-inflammatory Effect of Lithospermum erythrorhizon on Lipopolysaccharide-induced Inflammatory Response in RAW 264.7 Cells. Korea J. Herbol. 2013, 28, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.H.; Sim, G.S.; Choi, H.Y.; Ha, W.K. A Study on the Natural Preservative(Lactobacillus-fermented antimicrobial solution), Fermented with Plant Originated Lactic Acid Bacteria. Food Sci. Ind. 2011, 44, 45–51. [Google Scholar]
- Moon, Y.L.; You, J.W.; Shim, H.J. Arthroscopic Management of The Septic Gonitis. Knee Surg. Relat. Res. 2001, 13, 114–116. [Google Scholar]
- Seo, S.T.; Lee, J.S.; Park, J.H.; Han, K.S.; Jang, H.I. Investigation of Antibiotic Susceptibility of Some Plant Pathogenic Bacteria. Korean J. Hort. Sci. Technol. 2005, 23, 495–498. [Google Scholar]
- Jeong, I.; Heo, E.Y.; Lee, J.S.; Kim, H.R.; Lee, E.Y.; Kim, Y.W.; Yang, S.C. Streptococcus salivarius Pneumonia with Pulmonary Nocardiosis in a Rheumatoid Arthritis Patient Treated with Immunosuppressants. Korean J. Med. 2009, 77, 128–132. [Google Scholar]
- Lee, J.Y.; Yoo, D.H.; Joo, D.H.; Kim, S.R.; Jo, H.S.; Joo, S.H.; Chae, J.W. Anti-inflammatory Effects of Amelanchier asiatica Fruits Ethanol Extract. J. Soc. Cosmet. Sci. Korea 2017, 43, 19–26. [Google Scholar]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Shinmura, K.; Tang, X.L.; Kodani, E.; Xuan, Y.T.; Guo, Y.; Dawn, B. Discovery of a New Function of Cyclooxygenase (COX)-2: COX-2 is a Cardioprotective Protein that Alleviates Ischemia/Reperfusion Injury and Mediates the Late Phase of Preconditioning. Cardiovasc. Res. 2002, 55, 506–519. [Google Scholar] [CrossRef]
- Patrono, C. Cardiovascular Effects of Cyclooxygenase-2 Inhibitors: A Mechanistic and Clinical Perspective. Br. J. Clin. Pharmacol. 2016, 82, 957–964. [Google Scholar] [CrossRef]
- Jang, H.H. A Study on the Popularization Process of Chinese Tea. J. Chin. Hum. 2018, 8, 509–535. [Google Scholar]
- Kim, H.S.; Ham, J.S. Antioxidative Ability of Lactic Acid Bacteria. Korean Food Sci. Anim. Resour. 2003, 23, 186–192. [Google Scholar]
- Kobayashi, T.; Kimura, B.; Fujii, T. Strictly Anaerobic Halophiles Isolated from Canned Swedish Fermented Herrings (Surströmming). Int. J. Food Microbiol. 2000, 54, 81–89. [Google Scholar] [CrossRef]
- Barrangou, R.; Yoon, S.S.; Breidt, F., Jr.; Fleming, H.P.; Klaenhammer, T.R. Characterization of Six Leuconostoc fallax Bacteriophages Isolated from an Industrial Sauerkraut Fermentation. Appl. Environ. Microbiol. 2002, 68, 5452–5458. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, W.; Aoyagi, A. History of Tempeh; Soyinfo Center: Lafayette, CA, USA, 2007; p. 85. [Google Scholar]
- Feng, C.H. The Tale of Sushi: History and Regulations. Compr. Rev. Food Sci. Food Saf. 2012, 11, 205–220. [Google Scholar] [CrossRef]
- Choi, W.S.; Kwon, H.S.; No, R.H.; Choi, G.P.; Lee, H.Y. Enhancement of Anti-inflammatory Activities of Fermented Scutellaria baicalensis Extracts using Lactobacillus rhamnosus. J. Soc. Cosmet. Sci. Korea 2013, 39, 303–311. [Google Scholar]
- Song, H.S.; Eom, S.H.; Kang, Y.M.; Choi, J.D.; Kim, Y.M. Enhancement of the Antioxidant and Anti-inflammatory Activity of Hizikia fusiforme Water Extract by Lactic Acid Bacteria Fermentation. Korean J. Fish Aquat. Sci. 2011, 44, 111–117. [Google Scholar]
- Lim, H.W.; Lee, Y.J.; Huang, Y.H.; Yoon, J.Y.; Lee, S.H.; Kim, K.H.; Lim, C.J. Enhancement of Skin Antioxidant and Anti-Inflammatory Potentials of Agastache rugosa Leaf Extract by Probiotic Bacterial Fermentation in Human Epidermal Keratinocytes. Microbiol. Biotechnol. Lett. 2017, 45, 35–42. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, Y.K. Ethanol Extract of Forsythiae fructus Inhibits the Production of Inflammatory Mediators in LPS-stimulated BV-2 Microglial Cells. Korea J. Herbol. 2008, 23, 93–102. [Google Scholar]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyun, S.B.; Hyun, C.-G. Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage. Fermentation 2020, 6, 87. https://doi.org/10.3390/fermentation6030087
Hyun SB, Hyun C-G. Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage. Fermentation. 2020; 6(3):87. https://doi.org/10.3390/fermentation6030087
Chicago/Turabian StyleHyun, Su Bin, and Chang-Gu Hyun. 2020. "Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage" Fermentation 6, no. 3: 87. https://doi.org/10.3390/fermentation6030087
APA StyleHyun, S. B., & Hyun, C. -G. (2020). Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage. Fermentation, 6(3), 87. https://doi.org/10.3390/fermentation6030087