Snails as Taxis for a Large Yeast Biodiversity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Yeast Isolation
2.3. Molecular Typing of Yeast Isolates
2.4. Whole Genome Shotgun Sequencing
2.5. Fermentation Conditions
2.6. Analytical Methods
3. Results
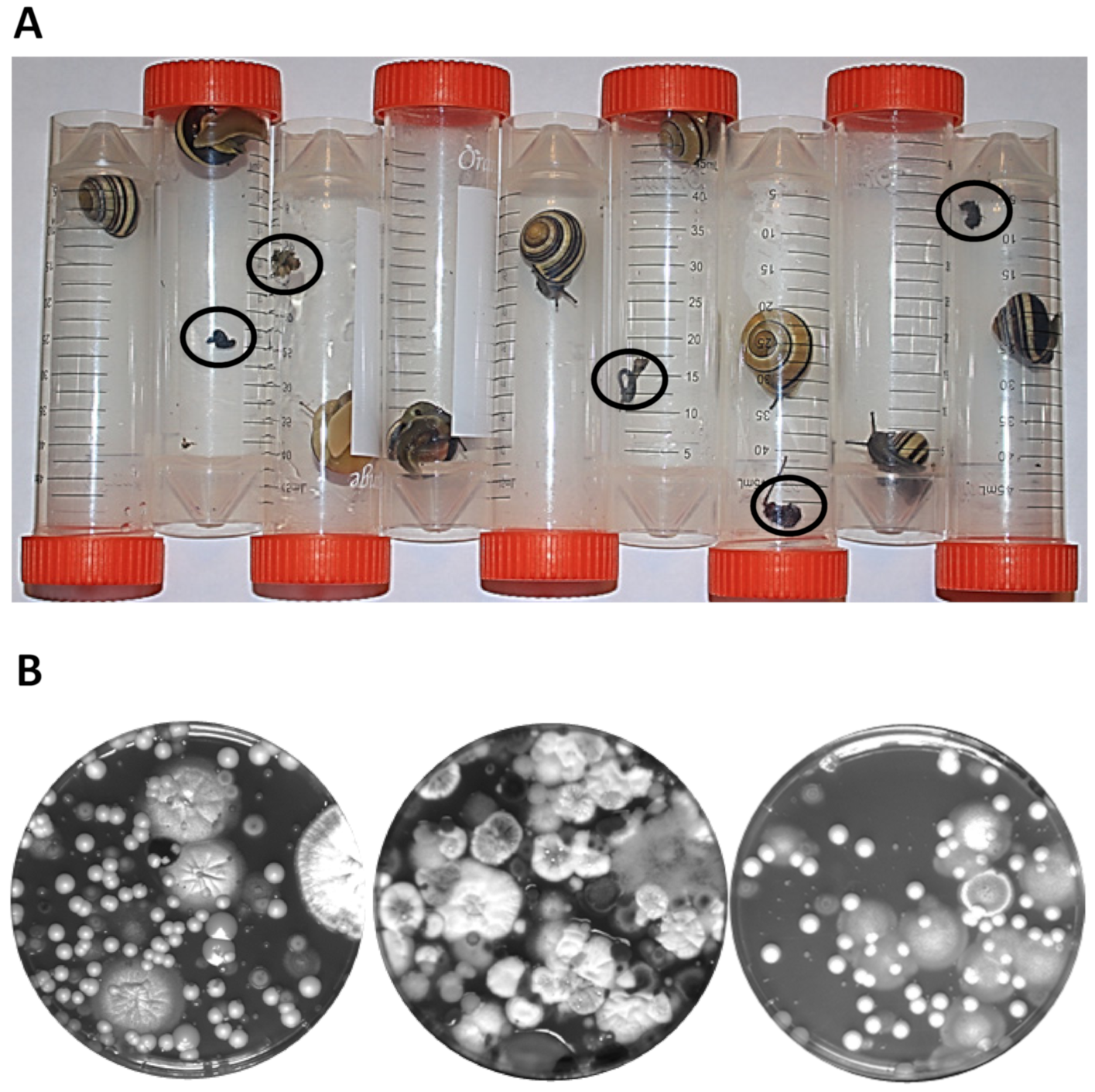
3.1. Isolating Yeast Strains from Snails
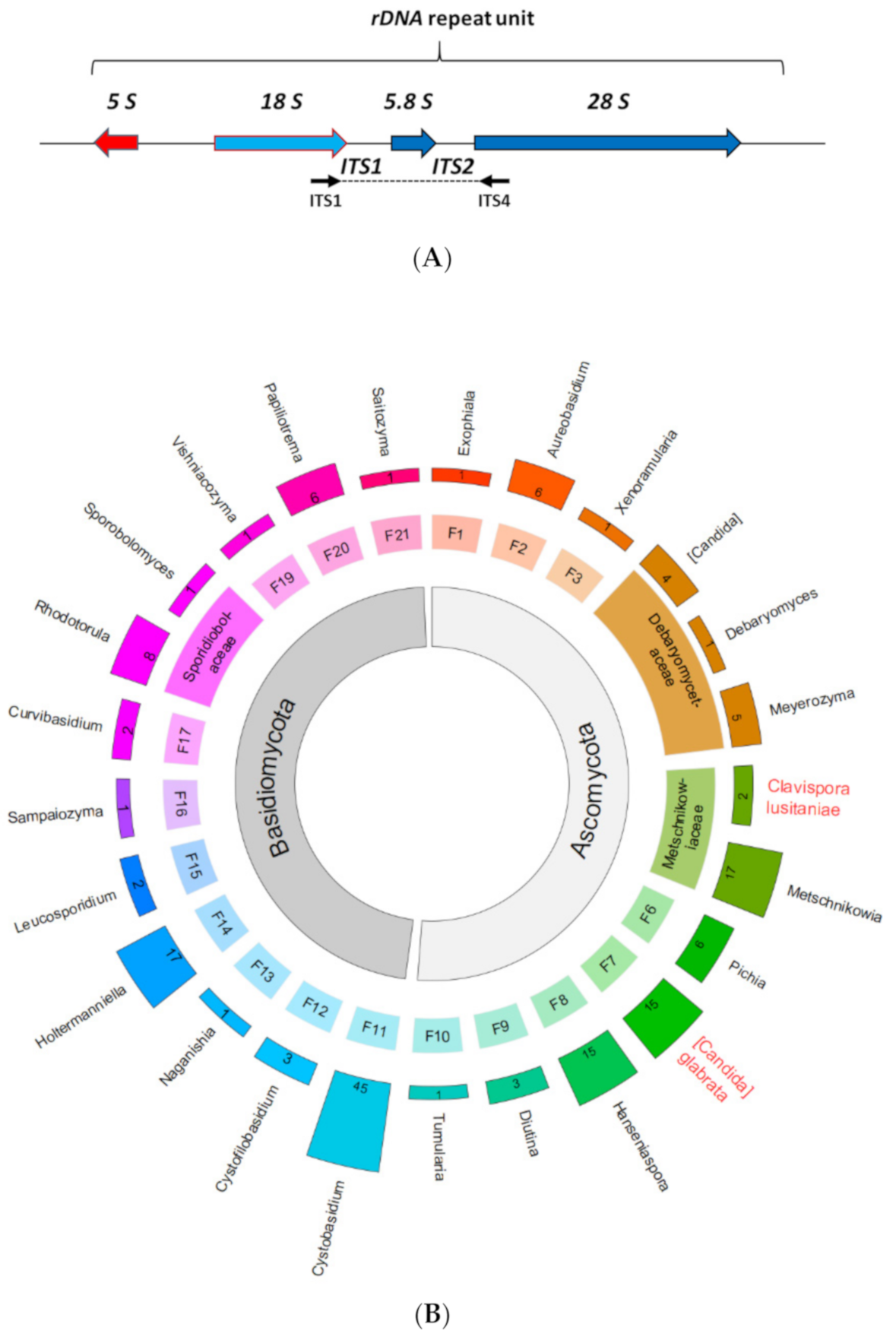
3.2. Typing of Yeast Isolates
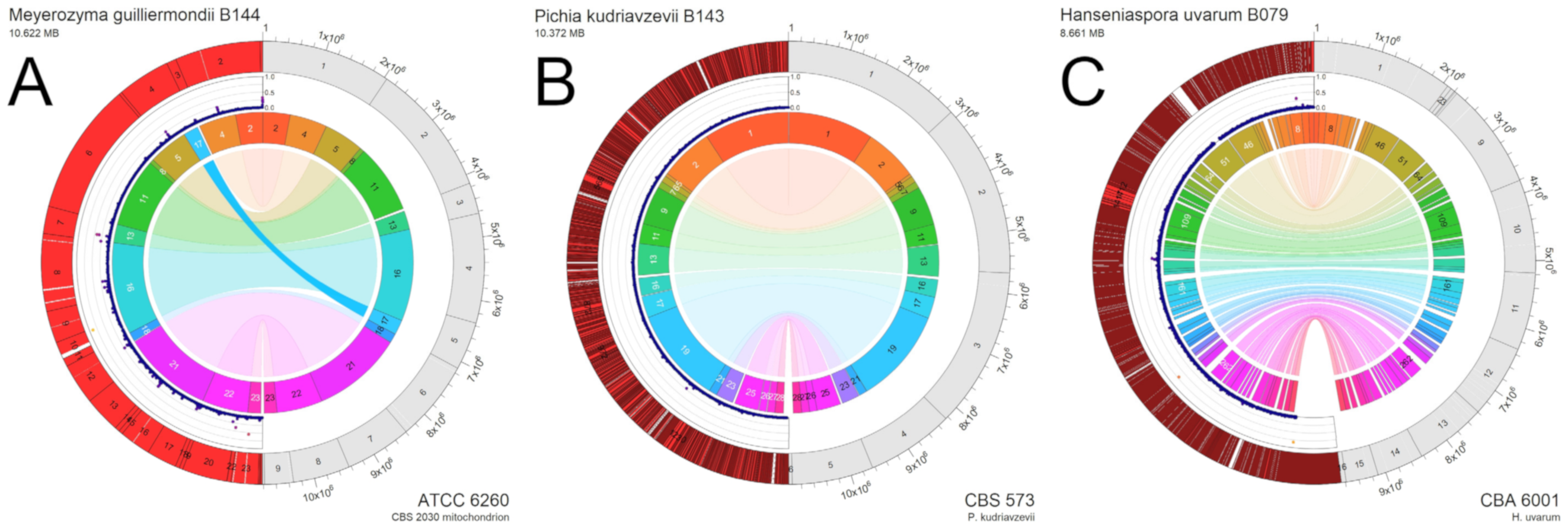
3.3. Genome Sequencing
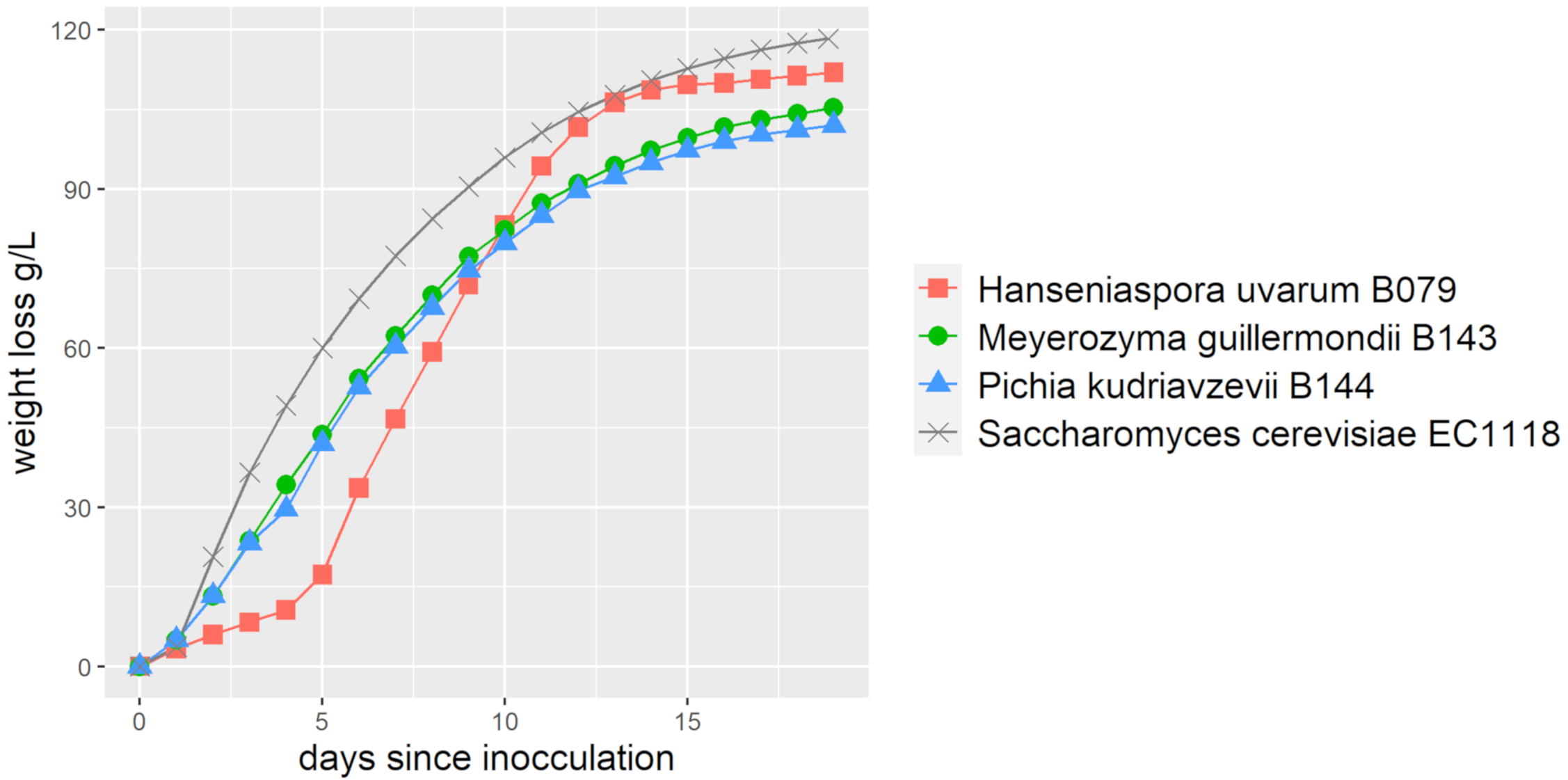
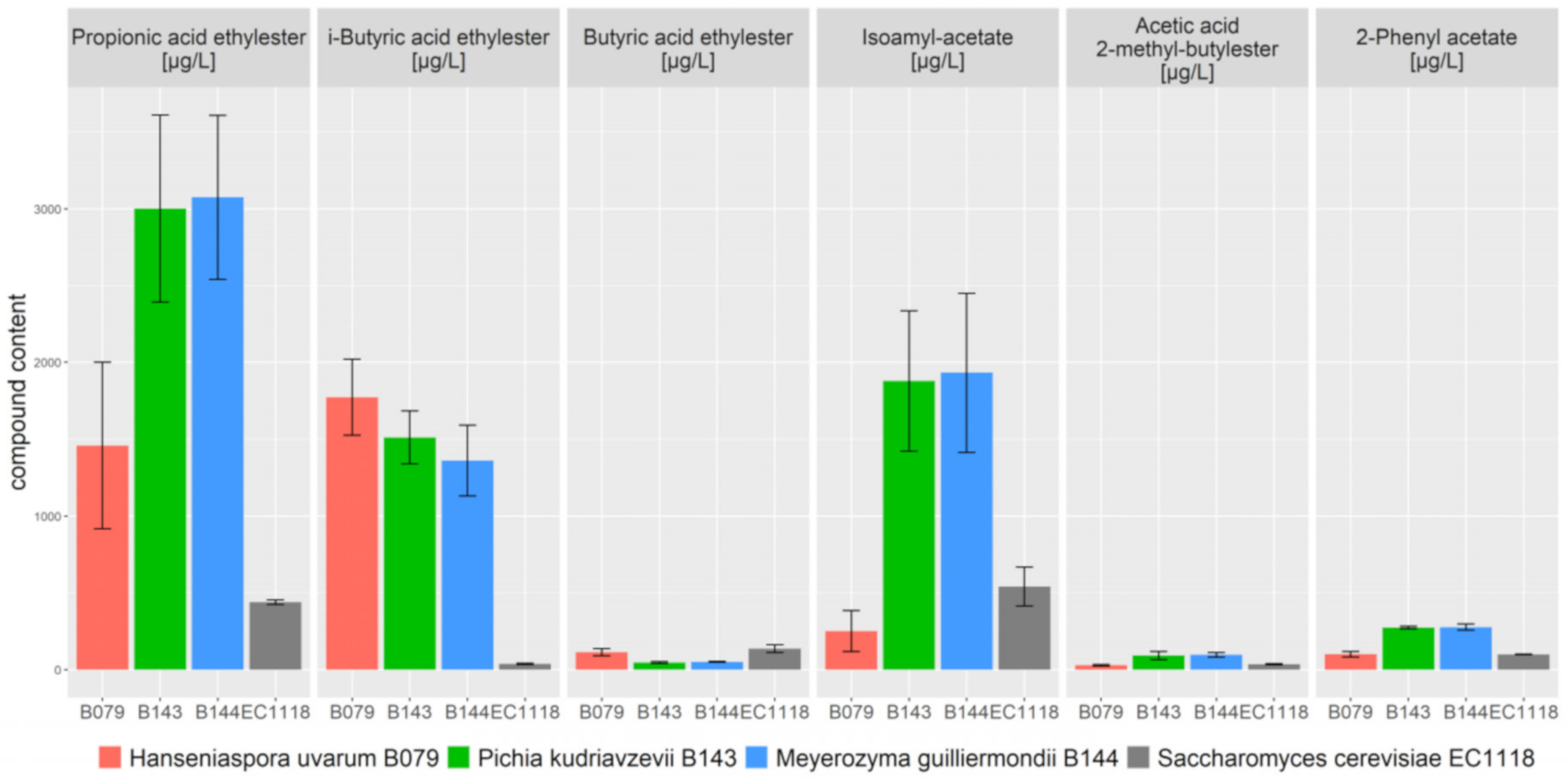
3.4. Fermentation Characteristics of Non-Conventional Yeasts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Botstein, D.; Fink, G.R. Yeast: An experimental organism for 21st Century biology. Genetics 2011, 189, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Starmer, W.T.; Lachance, M.A. Yeast ecology. In The Yeasts: A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier Science: London, UK; Burlington, MA, USA, 2011; pp. 65–83. [Google Scholar]
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition assays and physiological experiments of soil and phyllosphere yeasts identify Candida subhashii as a novel antagonist of filamentous fungi. BMC Microbiol. 2017, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Chappell, C.R.; Fukami, T. Nectar yeasts: A natural microcosm for ecology. Yeast 2018, 35, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Ganter, P.F. Yeast and invertebrate associations. In Biodiversity and Ecophysiology of Yeasts; Rosa, C.A., Peter, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 303–370. [Google Scholar]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect-yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [Green Version]
- Klaps, J.; Lievens, B.; Alvarez-Perez, S. Towards a better understanding of the role of nectar-inhabiting yeasts in plant-animal interactions. Fungal. Biol. Biotechnol. 2020, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Dapporto, L.; Legras, J.L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [Green Version]
- Palanca, L.; Gaskett, A.C.; Gunther, C.S.; Newcomb, R.D.; Goddard, M.R. Quantifying variation in the ability of yeasts to attract Drosophila melanogaster. PLoS ONE 2013, 8, e75332. [Google Scholar] [CrossRef] [Green Version]
- Murgier, J.; Everaerts, C.; Farine, J.P.; Ferveur, J.F. Live yeast in juvenile diet induces species-specific effects on Drosophila adult behaviour and fitness. Sci. Rep. 2019, 9, 8873. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Dapporto, L.; Berna, L.; Polsinelli, M.; Turillazzi, S.; Cavalieri, D. Social wasps are a Saccharomyces mating nest. Proc. Natl. Acad. Sci. USA 2016, 113, 2247–2251. [Google Scholar] [CrossRef] [Green Version]
- Christiaens, J.F.; Franco, L.M.; Cools, T.L.; De Meester, L.; Michiels, J.; Wenseleers, T.; Hassan, B.A.; Yaksi, E.; Verstrepen, K.J. The fungal aroma gene ATF1 promotes dispersal of yeast cells through insect vectors. Cell Rep. 2014, 9, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Proffit, M.; Rozpedowska, E.; Becher, P.G.; Andreadis, S.; Coracini, M.; Lindblom, T.U.; Ream, L.J.; Hagman, A.; Bengtsson, M.; et al. “This is not an apple”-yeast mutualism in codling moth. J. Chem. Ecol. 2012, 38, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Tatum, E.L. Nutritional requirements of drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1939, 25, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.O.; Noda, H.; Blackwell, M. Insect symbiosis: Derivation of yeast-like endosymbionts within an entomopathogenic filamentous lineage. Mol. Biol. Evol. 2001, 18, 995–1000. [Google Scholar] [CrossRef]
- Walther, A.; Wendland, J. Yap1-dependent oxidative stress response provides a link to riboflavin production in Ashbya gossypii. Fungal Genet. Biol. 2012, 49, 697–707. [Google Scholar] [CrossRef]
- Dietrich, F.S.; Voegeli, S.; Kuo, S.; Philippsen, P. Genomes of Ashbya fungi isolated from insects reveal four mating-type loci, numerous translocations, lack of transposons, and distinct gene duplications. G3 (Bethesda) 2013, 3, 1225–1239. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, T.Q.; Silva, R.; Domingues, L. Ashbya gossypii beyond industrial riboflavin production: A historical perspective and emerging biotechnological applications. Biotechnol. Adv. 2015, 33, 1774–1786. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, J.L.; Ledesma-Amaro, R.; Lozano-Martinez, P.; Diaz-Fernandez, D.; Buey, R.M.; Jimenez, A. Bioproduction of riboflavin: A bright yellow history. J. Ind. Microbiol. Biotechnol. 2017, 44, 659–665. [Google Scholar] [CrossRef]
- Wendland, J. Special issue: Non-conventional yeasts: Genomics and biotechnology. Microorganisms 2019, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Alper, H.S. Non-conventional hosts for the production of fuels and chemicals. Curr. Opin. Chem. Biol. 2020, 59, 15–22. [Google Scholar] [CrossRef]
- Reis, E.J.C.; Buscariolo, F.; Siqueira, J.P.Z.; Castilho, E.M.; Almeida, M.T.G. Agapornis sp. pet birds: Source of dissemination of azole-resistant yeasts. Med. Mycol. 2019, 57, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A.; Williamson, D.A. Method of isolating Protoplasts from Yeast. Nature 1957, 179, 1252–1253. [Google Scholar] [CrossRef]
- Johnston, J.R.; Mortimer, R.K. Use of snail digestive juice in isolation of yeast spore tetrads. J. Bacteriol. 1959, 78, 292. [Google Scholar] [CrossRef] [Green Version]
- Millbank, J.W.; Macrae, R.M. Degradation of yeast cell wall by fractionated snail gut enzyme. Nature 1964, 201, 1347. [Google Scholar] [CrossRef]
- Anderson, F.B.; Millbank, J.W. Protoplast formation and yeast cell-wall structure. The action of the enzymes of the snail, Helix pomatia. Biochem. J. 1966, 99, 682–687. [Google Scholar] [CrossRef] [Green Version]
- Cilia, G.; Fratini, F. Antimicrobial properties of terrestrial snail and slug mucus. J. Complement. Integr. Med. 2018, 15. [Google Scholar] [CrossRef]
- Whitson, M. Cepaea nemoralis (Gastropoda, Helicidae): The invited invader. J. Ky. Acad. Sci. 2005, 66, 82–88. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Kwiecinski, Z.; Lesicki, A.; Skorka, P.; Kobak, J.; Szymanska, A.; Osiejuk, T.S.; Kaluski, T.; Jaskulska, M.; Tryjanowski, P. Shell colour, temperature, (micro)habitat structure and predator pressure affect the behaviour of Cepaea nemoralis. Naturwissenschaften 2018, 105, 35. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Rissman, A.I.; Mau, B.; Biehl, B.S.; Darling, A.E.; Glasner, J.D.; Perna, N.T. Reordering contigs of draft genomes using the Mauve aligner. Bioinformatics 2009, 25, 2071–2073. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize Implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matti, K.; Bernardi, B.; Brezina, S.; Semmler, H.; von Wallbrunn, C.; Rauhut, D.; Wendland, J. Characterization of old wine yeasts kept for decades under a zero-emission maintenance regime. Fermentation 2020, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Surmacki, A.; Ozarowska-Nowicka, A.; Rosin, Z.M. Color polymorphism in a land snail Cepaea nemoralis (Pulmonata: Helicidae) as viewed by potential avian predators. Naturwissenschaften 2013, 100, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.; Jackson, H.J.; Murphy, E.W.; Reader, T. Discrete or indiscrete? Redefining the colour polymorphism of the land snail Cepaea nemoralis. Heredity 2019, 123, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An emerging pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465717. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, A.A.; Harrison, L.H.; Farley, M.M.; Hollick, R.; Stein, B.; Chiller, T.M.; Lockhart, S.R.; Park, B.J. Declining incidence of candidemia and the shifting epidemiology of Candida resistance in two US metropolitan areas, 2008–2013: Results from population-based surveillance. PLoS ONE 2015, 10, e0120452. [Google Scholar] [CrossRef] [Green Version]
- Setati, M.E.; Jacobson, D.; Andong, U.C.; Bauer, F.F. The vineyard yeast microbiome, a mixed model microbial map. PLoS ONE 2012, 7, e52609. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genoves, S.; Valles, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Brysch-Herzberg, M. Ecology of yeasts in plant-bumblebee mutualism in Central Europe. FEMS Microbiol. Ecol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Guzman, B.; Lachance, M.A.; Herrera, C.M. Phylogenetic analysis of the angiosperm-floricolous insect-yeast association: Have yeast and angiosperm lineages co-diversified? Mol. Phylogenet. Evol. 2013, 68, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, M. Made for Each Other: Ascomycete Yeasts and Insects. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Keebaugh, E.S.; Yamada, R.; Ja, W.W. The nutritional environment influences the impact of microbes on drosophila melanogaster life span. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [Green Version]
- Gabaldon, T.; Fairhead, C. Genomes shed light on the secret life of Candida glabrata: Not so asexual, not so commensal. Curr. Genet. 2019, 65, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic budding yeasts isolated outside of clinical settings. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef]
- Al-Yasiri, M.H.; Normand, A.C.; L’Ollivier, C.; Lachaud, L.; Bourgeois, N.; Rebaudet, S.; Piarroux, R.; Mauffrey, J.F.; Ranque, S. Opportunistic fungal pathogen Candida glabrata circulates between humans and yellow-legged gulls. Sci. Rep. 2016, 6, 36157. [Google Scholar] [CrossRef] [Green Version]
- Chenot, C.; Briffoz, L.; Lomartire, A.; Collin, S. Occurrence of ehrlich-derived and varietal polyfunctional thiols in belgian white wines made from chardonnay and solaris grapes. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of hanseniaspora uvarum in simultaneous and sequential co-fermentation with saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Bora, S.S.; Keot, J.; Das, S.; Sarma, K.; Barooah, M. Metagenomics analysis of microbial communities associated with a traditional rice wine starter culture (Xaj-pitha) of Assam, India. 3 Biotech. 2016, 6, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.X.; Wang, G.Q.; Aihaiti, A. Combined indigenous yeast strains produced local wine from over ripen Cabernet Sauvignon grape in Xinjiang. World J. Microbiol. Biotechnol. 2020, 36, 122. [Google Scholar] [CrossRef] [PubMed]
- Bekatorou, A.; Psarianos, C.; Koutinas, A.A. Production of food grade yeasts. Food Technol. Biotechnol. 2006, 44, 407–415. [Google Scholar]
- Douglass, A.P.; Offei, B.; Braun-Galleani, S.; Coughlan, A.Y.; Martos, A.A.R.; Ortiz-Merino, R.A.; Byrne, K.P.; Wolfe, K.H. Population genomics shows no distinction between pathogenic Candida krusei and environmental Pichia kudriavzevii: One species, four names. PLoS Pathog. 2018, 14, e1007138. [Google Scholar] [CrossRef]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; BŁażejak, S.; Gientka, I.; Kurcz, A. Biotechnological use of Candida yeasts in the food industry: A review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and emerged pathogenic Candida species: Beyond the Candida albicans paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef] [Green Version]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Cadez, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLoS Biol. 2019, 17, e3000255. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucose | Fructose | Total Sugars | Tartaric Acid | Malate | Shikimic Acid | Lactate | Acetate | Citric Acid | Ethanol | Ethanol | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| strain | [g/L] | [g/L] | [g/L] | [g/L] | [g/L] | [mg/L] | [g/L] | [g/L] | [g/L] | [g/L] | [%] |
| B079 (H. uvarum) | <1 | <1 | 0 | 4.1 ± 0.1 | 2.4 ± 0.1 | 28.3 ± 0.6 | 0.2 ± 0 | 0.2 ± 0.1 | 0.2 ± 0 | 79.7 ± 0.4 | 10.1 ± 0.1 |
| B143 (P. kudriavzevii) | <1 | 16.1 ± 2.1 | 16.1 ± 2.1 | 4.2 ± 0.1 | 1.7 ± 0 | 32 ± 0 | 0.2 ± 0 | 0.8 ± 0.1 | 0.2 ± 0 | 66.7 ± 0.4 | 8.4 ± 0.1 |
| B144 (M. guilliermondii) | 1 | 18.5 ± 6.5 | 18.8 ± 7.1 | 4.2 ± 0 | 1.8 ± 0.1 | 32 ± 0 | 0.2 ± 0 | 0.7 ± 0.1 | 0.2 ± 0 | 66.2 ± 3.4 | 8.4 ± 0.5 |
| EC1118 (S. cerevisiae) | 1.6 | 18.6 | 20.2 | 3.7 | 2.7 | 31 | 0.12 | 0.39 | 0.13 | 77.7 | 9.9 |
| Strain | Acetic Acid Ethylester [mg/L] | i-Butanol [mg/L] | Propionic Acid Ethylester [µg/L] | Isoamyl-Alcohol [mg/L] | 2-Methyl-Butanol [mg/L] | i-Butyric Acid Ethylester [µg/L] | Butyric Acid Ethylester [µg/L] | i-Valeric Acid [µg/L] | Hexanol [µg/L] | Isoamyl-Acetate [µg/L] | Acetic Acid 2-Methyl-Butylester [µg/L] | 2-Phenyl-Ethanol [mg/L] | 2-Phenyl Acetate [µg/L] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B079 (H. uvarum) | 425 ± 51 | 251 ± 61 | 1459 ± 541 | 375 ± 60 | 89 ± 19 | 1773 ± 247 | 114 ± 24 | 2074 ± 15 | 884 ± 93 | 251 ± 132 | 28 ± 6 | 48 ± 12 | 100 ± 18 |
| B143 (P. kudriavzevii) | 868 ± 213 | 494 ± 64 | 3002 ± 609 | 361 ± 47 | 104 ± 11 | 1512 ± 173 | 46 ± 7 | 2479 ± 195 | 436 ± 39 | 1878 ± 457 | 92 ± 27 | 58 ± 7 | 275 ± 9 |
| B144 (M. guilliermondii) | 883 ± 68 | 497 ± 62 | 3075 ± 534 | 355 ± 32 | 104 ± 8 | 1361 ± 231 | 51 ± 3 | 2090 ± 90 | 438 ± 0 | 1932 ± 519 | 97 ± 15 | 56 ± 3 | 278 ± 21 |
| EC1118 (S. cerevisiae) | 36 ± 4 | 60 ± 5 | 438 ± 15 | 402 ± 35 | 71 ± 4 | 38 ± 4 | 137 ± 25 | 1668 ± 103 | 1542 ± 29 | 540 ± 127 | 36 ± 4 | 57 ± 4 | 98 ± 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akan, M.; Michling, F.; Matti, K.; Krause, S.; Muno-Bender, J.; Wendland, J. Snails as Taxis for a Large Yeast Biodiversity. Fermentation 2020, 6, 90. https://doi.org/10.3390/fermentation6030090
Akan M, Michling F, Matti K, Krause S, Muno-Bender J, Wendland J. Snails as Taxis for a Large Yeast Biodiversity. Fermentation. 2020; 6(3):90. https://doi.org/10.3390/fermentation6030090
Chicago/Turabian StyleAkan, Madina, Florian Michling, Katrin Matti, Sinje Krause, Judith Muno-Bender, and Jürgen Wendland. 2020. "Snails as Taxis for a Large Yeast Biodiversity" Fermentation 6, no. 3: 90. https://doi.org/10.3390/fermentation6030090
APA StyleAkan, M., Michling, F., Matti, K., Krause, S., Muno-Bender, J., & Wendland, J. (2020). Snails as Taxis for a Large Yeast Biodiversity. Fermentation, 6(3), 90. https://doi.org/10.3390/fermentation6030090