A Comprehensive Understanding of Electro-Fermentation


 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
1.1. The Origin of Bioelectrochemical Systems and Electrofermentaion
1.2. Electro-Fermentation and its Principles
1.3. Anodic Fermentation
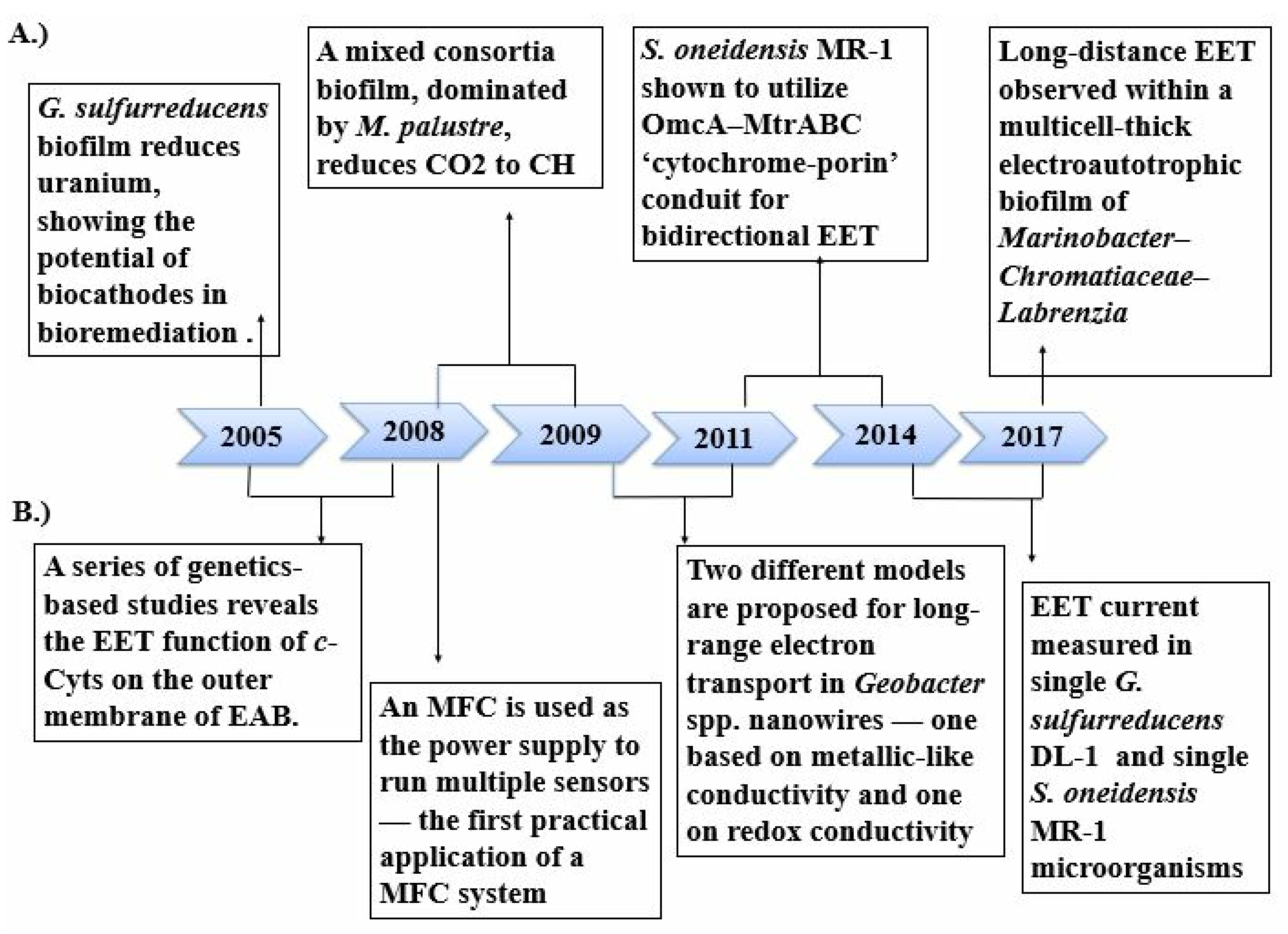
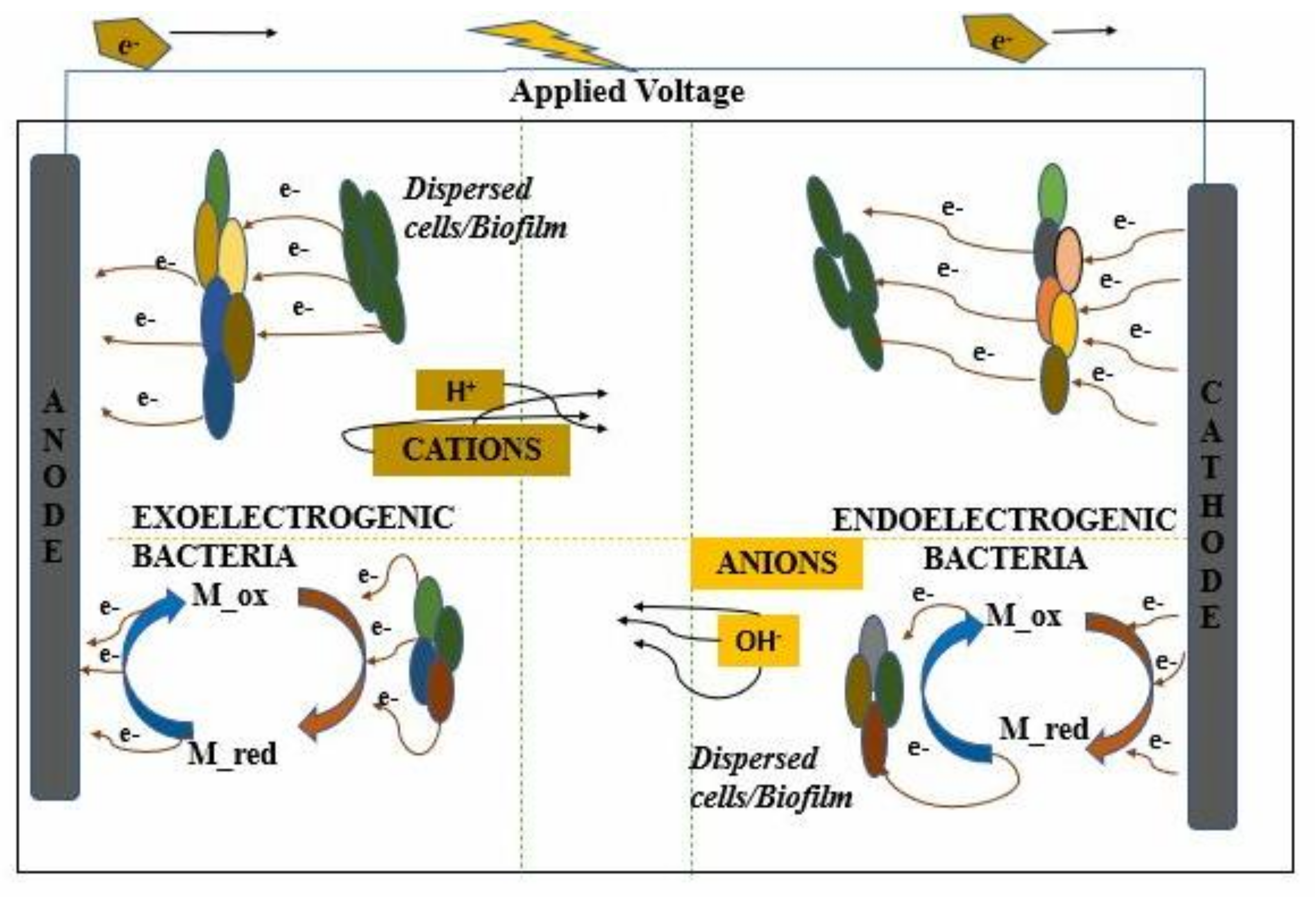
2. Microbe–Electrode Interaction
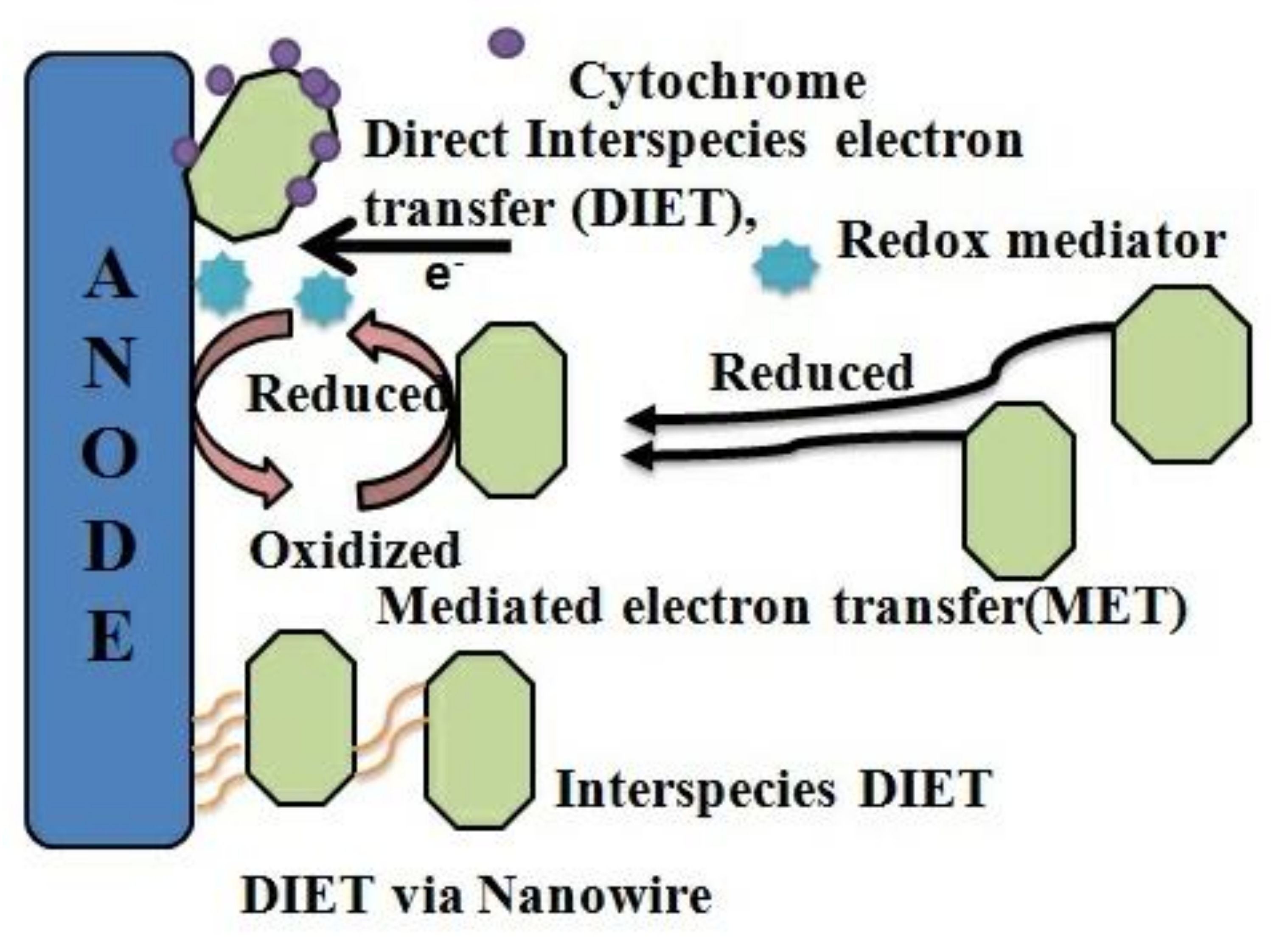
2.1. Electron Transfer Mechanism
2.1.1. Anode Electron Transfer
2.1.2. Cathode Electron Transfer
2.1.3. Applied Voltage as a Power Source
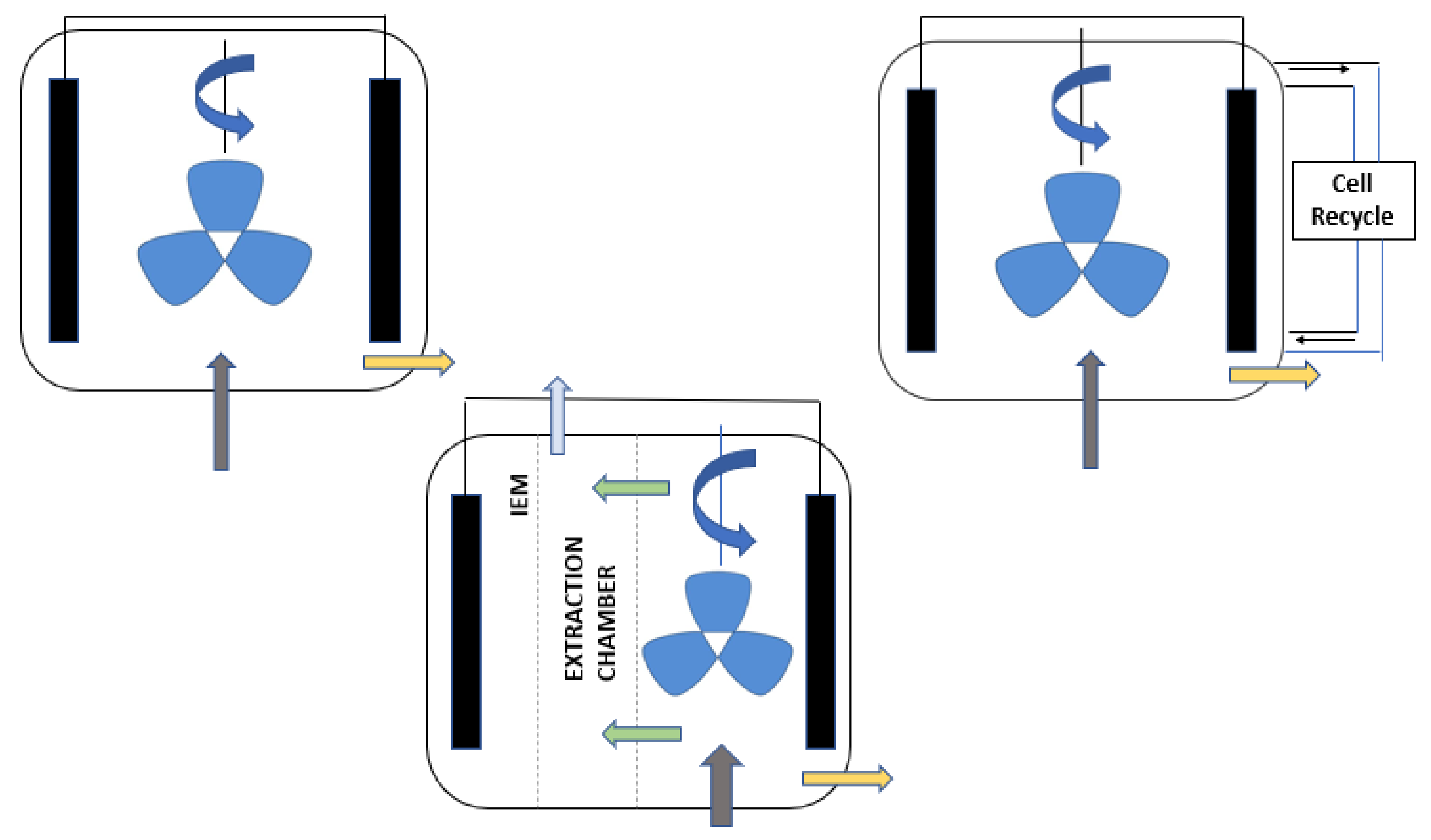
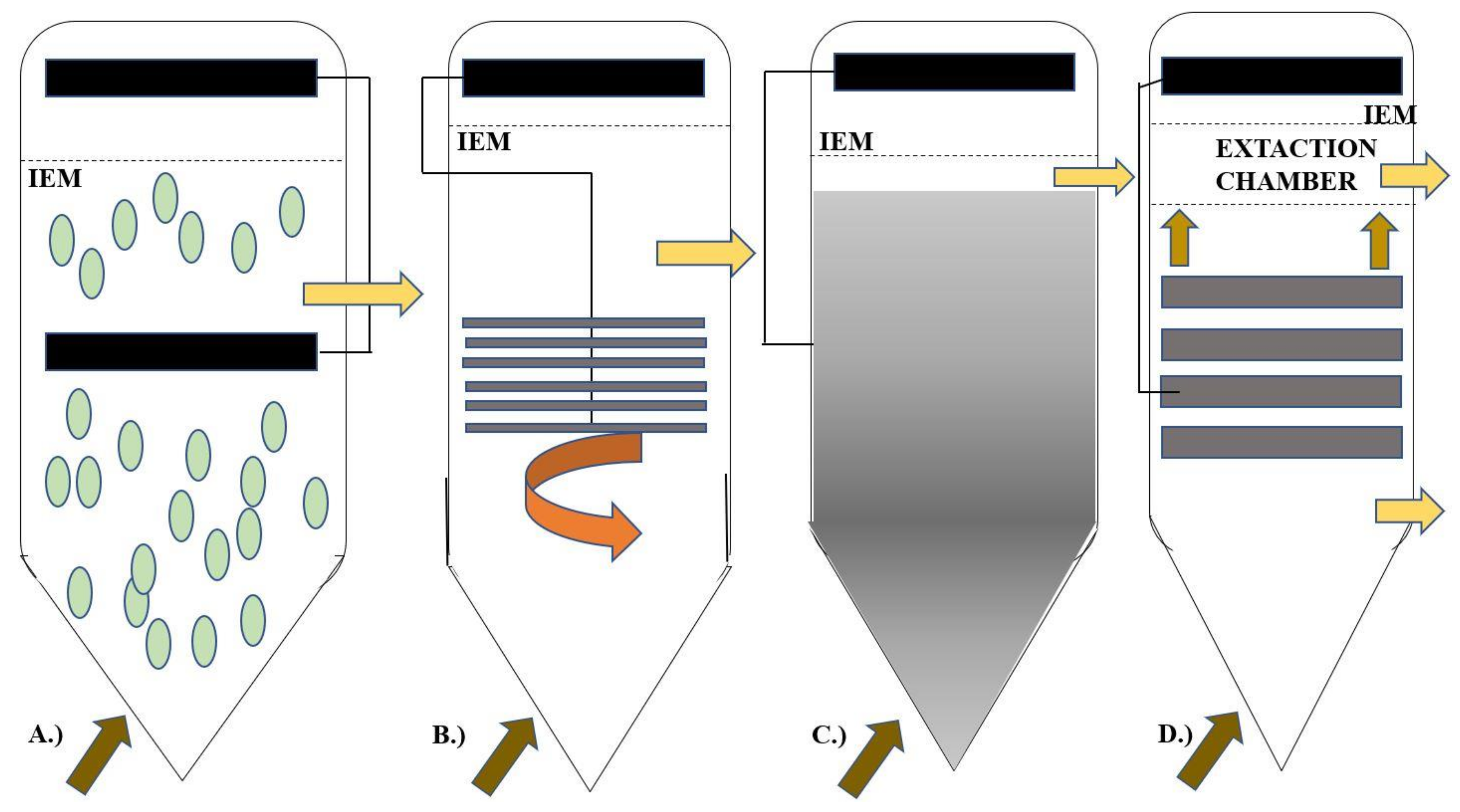
3. Electro-Fermenter Design
4. Mixed Microbial Cultures Used in Electro Fermentation Processes in Industrial Biomass Refineries
5. Industrial Fermentations of Pure Cultures Using an Electro Fermentation Approach
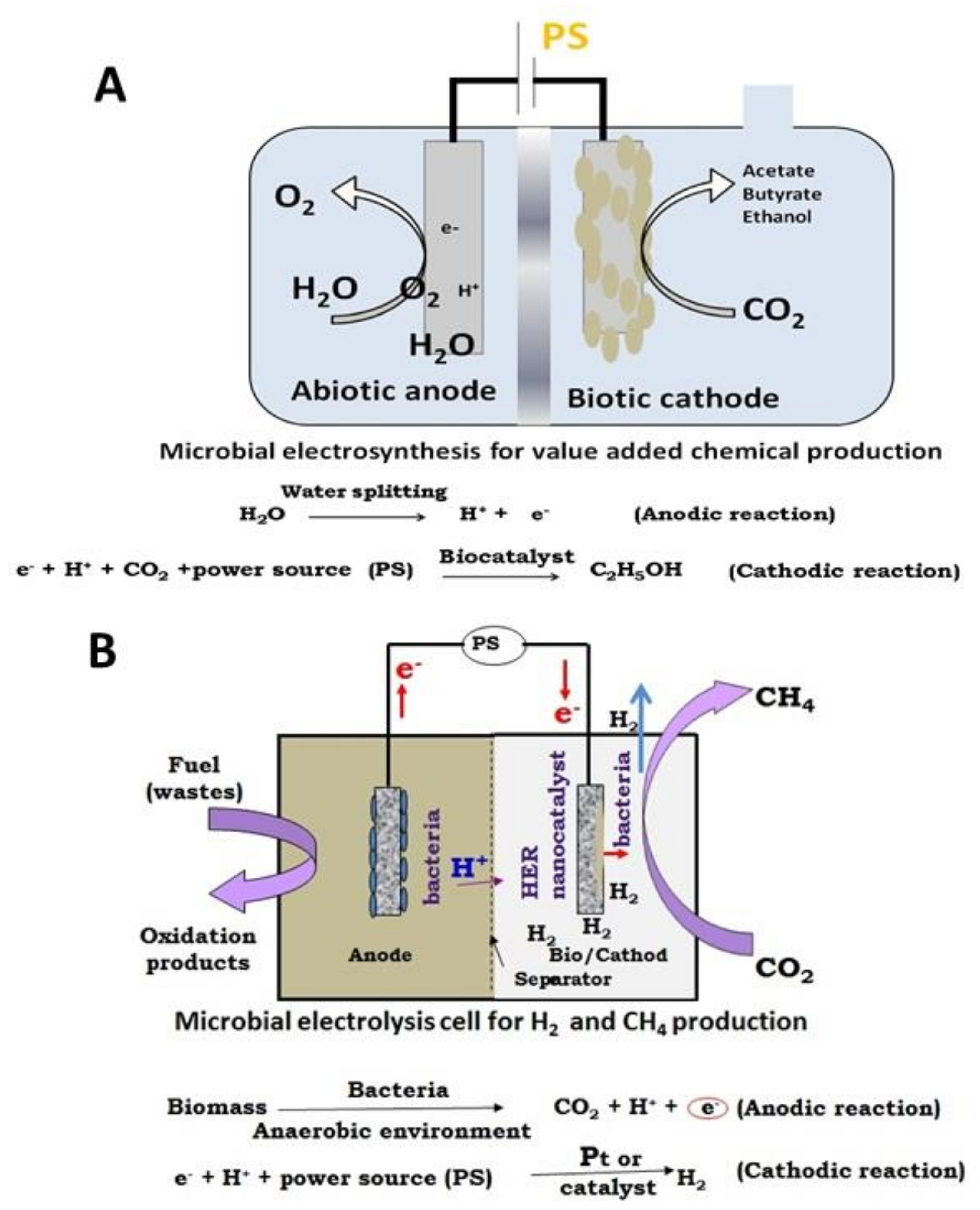
6. Microbial Electrosynthesis for Value-Added Chemicals-Reactors Design and Recent Development
6.1. Integrating MEC-Microbial Fuel Cell (MFC)
6.2. Submersible Microbial Electrolysis Cell
6.3. Integration of Pyrolysis-MEC
6.4. Integrating MEC and Process of Lignocellulosic Ethanol Biorefinery
7. Bio-Electro Products
7.1. Short-Chain Fatty Acids (SCFAs)
7.2. Medium-Chain Fatty Acids (MCFA)
7.3. Bio Alcohols
7.4. Biohydrogen
7.5. Biomethane
7.6. CO2 to Value-Added Products
8. Microbial Electrolysis for Hydrogen-Reactors Design and Recent Development
9. Setbacks in Lactic Acid Fermentation
10. Current Challenges and Impediments to Scaling-Up the MEC
11. Future Perspectives
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EF | Electro-fermentation |
| PHA | Polyhydroxyalkanoates |
| PHB | Polyhydroxybutyrate |
| BES | Bioelectrochemical systems |
| AF | Anaerobic fermentation |
| ATP | Adenosine triphosphate |
| Moxd | Medox |
| MRed | Med RED |
| MET | Microbial electron transfer |
| EET | Extracellular electron transfer |
| NAD | Nicotinamide adenine dinucleotide |
| MFC | Microbial fuel cells |
| MES | Microbial Electrosynthesis |
| MHC | Major histocompatibility complex |
| MWCNT | Multi walled carbon nanotubes |
| NADH | Nicotinamide adenine dinucleotide hydrogen |
| EAB | Electroactive Bacteria |
| MEC | Microbial electrolysis cell |
| IEM | Ion exchange membrane |
| PEM | Proton exchange membrane |
| ORR | Oxygen reduction reaction |
| VFA | Volatile fatty acid |
| SCFA | Short-chain fatty acids |
| MCFA | Medium-chain fatty acids |
| BES | Bioelectrochemical systems |
| SHE | Standard hydrogen electrode |
| MET | Microbial electrosynthesis technology |
| PBS | Phosphate buffer saline/solution |
| σ | Ionic conductor |
| U | Cell voltage |
| An. | Anodic faradic yield |
| γG | Energy yield |
| γH | Thermal yield |
References
- Flynn, J.M.; Ross, D.E.; Hunt, K.A.; Bond, D.R.; Gralnick, J.A. Enabling Unbalanced Fermentations by Using Engineered Electrode-Interfaced Bacteria. mBio 2010, 1. [Google Scholar] [CrossRef] [Green Version]
- Soccol, C.R.; Pandey, A.; Larroche, C.; Pandey, A.; Larroche, C. Fermentation Processes Engineering in the Food Industry; CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-0-429-11200-3. [Google Scholar]
- Humphrey, A.E.; Lee, S.E. Industrial Fermentation: Principles, Processes, and Products. In Riegel’s Handbook of Industrial Chemistry; Springer: Dordrecht, The Netherlands, 1992; pp. 916–986. [Google Scholar]
- Albuquerque, M.G.E.; Eiroa, M.; Torres, C.; Nunes, B.R.; Reis, M.A.M. Strategies for the development of a side stream process for polyhydroxyalkanoate (PHA) production from sugar cane molasses. J. Biotechnol. 2007, 130, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Kaushik, N.; Biswas, S. Bioprocess and Bioproducts: Technology Trends and Opportunities; Asiatech Publishers: New Delhi, India, 2009; ISBN 8187680202. [Google Scholar]
- Schievano, A.; PepéSciarria, T.; Vanbroekhoven, K.; De Wever, H.; Puig, S.; Andersen, S.J.; Rabaey, K.; Pant, D. Electro-Fermentation—Merging Electrochemistry with Fermentation in Industrial Applications. Trends Biotechnol. 2016, 34, 866–878. [Google Scholar] [CrossRef]
- Angenent, L.T.; Karim, K.; Al-Dahhan, M.H.; Wrenn, B.A.; Domíguez-Espinosa, R. Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol. 2004, 22, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Vanbroekhoven, K.; Buisman, C.J.N.; Pant, D.; Strik, D.P.B.T.B. Application of gas diffusion biocathode in microbial electrosynthesis from carbon dioxide. Environ. Sci. Pollut. Res. 2016, 23, 22292–22308. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-P.; Zheng, P.; Sun, Z.-H.; Ni, Y.; Dong, J.-J.; Wei, P. Strategies of pH control and glucose-fed batch fermentation for production of succinic acid by Actinobacillus succinogenes CGMCC1593. J. Chem. Technol. Biotechnol. 2008, 83, 722–729. [Google Scholar] [CrossRef]
- Roy, S.; Schievano, A.; Pant, D. Electro-stimulated microbial factory for value added product synthesis. Bioresour. Technol. 2016, 213, 129–139. [Google Scholar] [CrossRef]
- Mohan, S.V.; Butti, S.K.; Amulya, K.; Dahiya, S.; Modestra, J.A. Waste Biorefinery: A New Paradigm for a Sustainable Bioelectro Economy. Trends Biotechnol. 2016, 34, 852–855. [Google Scholar] [CrossRef]
- Rabaey, K.; Angenent, L.; Schroder, U.; Keller, J. Bioelectrochemical Systems; IWA Publishing: London, UK, 2009; ISBN 978-1-84339-233-0. [Google Scholar]
- Thrash, J.C.; Coates, J.D. Review: Direct and Indirect Electrical Stimulation of Microbial Metabolism. Environ. Sci. Technol. 2008, 42, 3921–3931. [Google Scholar] [CrossRef]
- Lovley, D.R. Powering microbes with electricity: Direct electron transfer from electrodes to microbes. Environ. Microbiol. Rep. 2011, 3, 27–35. [Google Scholar] [CrossRef]
- Lee, H.-S.; Rittmann, B.E. Significance of Biological Hydrogen Oxidation in a Continuous Single-Chamber Microbial Electrolysis Cell. Environ. Sci. Technol. 2010, 44, 948–954. [Google Scholar] [CrossRef]
- Choi, O.; Sang, B.-I. Extracellular electron transfer from cathode to microbes: Application for biofuel production. Biotechnol. Biofuels 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikhil, G.N.; Venkata Subhash, G.; Yeruva, D.K.; Venkata Mohan, S. Synergistic yield of dual energy forms through biocatalyzedelectrofermentation of waste: Stoichiometric analysis of electron and carbon distribution. Energy 2015, 88, 281–291. [Google Scholar] [CrossRef]
- Logan, B.E. Scaling up microbial fuel cells and other bioelectrochemical systems. Appl. Microbiol. Biotechnol. 2010, 85, 1665–1671. [Google Scholar] [CrossRef]
- Rosenbaum, M.A.; Franks, A.E. Microbial catalysis in bioelectrochemical technologies: Status quo, challenges and perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, Z.J. A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol. Adv. 2013, 31, 1796–1807. [Google Scholar] [CrossRef]
- Leech, D.; Kavanagh, P.; Schuhmann, W. Enzymatic fuel cells: Recent progress. Electrochim. Acta 2012, 84, 223–234. [Google Scholar] [CrossRef]
- Habermüller, K.; Mosbach, M.; Schuhmann, W. Electron-transfer mechanisms in amperometric biosensors. Fresenius J. Anal. Chem. 2000, 366, 560–568. [Google Scholar] [CrossRef]
- Ghindilis, A.L.; Atanasov, P.; Wilkins, E. Enzyme-catalyzed direct electron transfer: Fundamentals and analytical applications. Electroanalysis 1997, 9, 661–674. [Google Scholar] [CrossRef]
- Kumar, A.; Hsu, L.H.-H.; Kavanagh, P.; Barrière, F.; Lens, P.N.L.; Lapinsonnière, L.; Lienhard V, J.H.; Schröder, U.; Jiang, X.; Leech, D. The ins and outs of microorganism–electrode electron transfer reactions. Nat. Rev. Chem. 2017, 1, 1–13. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Dipasquale, L.; Vella, F.M.; Romano, I.; Gambacorta, A.; Cutignano, A.; Fontana, A. Hydrogen metabolism in the extreme thermophile Thermotoganeapolitana. Int. J. Hydrog. Energy 2010, 35, 2290–2295. [Google Scholar] [CrossRef]
- Manzini, E.; Scaglia, B.; Schievano, A.; Adani, F. Dark fermentation effectiveness as a key step for waste biomass refineries: Influence of organic matter macromolecular composition and bioavailability. Int. J. Energy Res. 2015, 39, 1519–1527. [Google Scholar] [CrossRef]
- Agler, M.T.; Wrenn, B.A.; Zinder, S.H.; Angenent, L.T. Waste to bioproduct conversion with undefined mixed cultures: The carboxylate platform. Trends Biotechnol. 2011, 29, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Aryal, N.; Sarma, P.M.; Vanbroekhoven, K.; Lal, B.; Benetton, X.D.; Pant, D. Bioelectrocatalyzed reduction of acetic and butyric acids via direct electron transfer using a mixed culture of sulfate-reducers drives electrosynthesis of alcohols and acetone. Chem. Commun. Camb. Engl. 2013, 49, 6495–6497. [Google Scholar] [CrossRef]
- Metabolic and Practical Considerations on Microbial Electrosynthesis.—Abstract—Europe PMC. Available online: https://europepmc.org/article/med/21353525 (accessed on 20 June 2020).
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation—The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Effect of Applied Voltage, Initial Concentration, and Natural Organic Matter on Sequential Reduction/Oxidation of Nitrobenzene by Graphite Electrodes—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/22571797/ (accessed on 19 June 2020).
- Lovley, D.R. Syntrophy Goes Electric: Direct Interspecies Electron Transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Speers, A.M.; Young, J.M.; Reguera, G. Fermentation of Glycerol into Ethanol in a Microbial Electrolysis Cell Driven by a Customized Consortium. Environ. Sci. Technol. 2014, 48, 6350–6358. [Google Scholar] [CrossRef]
- Moscoviz, R.; Toledo-Alarcón, J.; Trably, E.; Bernet, N. Electro-Fermentation: How to Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef]
- Logan, B.E.; Call, D.; Cheng, S.; Hamelers, H.V.M.; Sleutels, T.H.J.A.; Jeremiasse, A.W.; Rozendal, R.A. Microbial Electrolysis Cells for High Yield Hydrogen Gas Production from Organic Matter. Environ. Sci. Technol. 2008, 42, 8630–8640. [Google Scholar] [CrossRef] [PubMed]
- Rago, L.; Ruiz, Y.; Baeza, J.A.; Guisasola, A.; Cortés, P. Microbial community analysis in a long-term membrane-less microbial electrolysis cell with hydrogen and methane production. Bioelectrochemistry 2015, 106, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Park, D.H.; Laivenieks, M.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Microbial Utilization of Electrically Reduced Neutral Red as the Sole Electron Donor for Growth and Metabolite Production. Appl. Environ. Microbiol. 1999, 65, 2912–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.; Onaka, Y.; Kobayashi, H.; Fu, Q.; Kawaguchi, H.; Vilcaez, J.; Sato, K. Mechanism of Electromethanogenic Reduction of CO2 by a Thermophilic Methanogen. Energy Procedia 2013, 37, 7021–7028. [Google Scholar] [CrossRef] [Green Version]
- Bader, J.; Mast-Gerlach, E.; Popović, M.K.; Bajpai, R.; Stahl, U. Relevance of microbial coculture fermentations in biotechnology. J. Appl. Microbiol. 2010, 109, 371–387. [Google Scholar] [CrossRef]
- Lenin Babu, M.; Venkata Subhash, G.; Sarma, P.N.; Venkata Mohan, S. Bio-electrolytic conversion of acidogenic effluents to biohydrogen: An integration strategy for higher substrate conversion and product recovery. Bioresour. Technol. 2013, 133, 322–331. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically conductive bacterial nanowires produced by Shewanellaoneidensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef] [Green Version]
- Rahimnejad, M.; Adhami, A.; Darvari, S.; Zirepour, A.; Oh, S.-E. Microbial fuel cell as new technology for bioelectricity generation: A review. Alex. Eng. J. 2015, 54, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Venkata Mohan, S.; Velvizhi, G.; Vamshi Krishna, K.; Lenin Babu, M. Microbial catalyzed electrochemical systems: A bio-factory with multi-facet applications. Bioresour. Technol. 2014, 165, 355–364. [Google Scholar] [CrossRef]
- Optimizing Treatment Performance of Microbial Fuel Cells by Reactor Staging Industrial & Engineering Chemistry Research. Available online: https://pubs.acs.org/doi/abs/10.1021/ie1004898 (accessed on 19 June 2020).
- Yu, J.; Song, X.; Wang, J.; Chen, J.; Liu, Q. BioelectrocatalyticDechlorination of Trichloroacetic Acid by Hemoglobin Modified Graphite Electrode in Aqueous Solutions of Different pH and Temperature. Int. J. Electrochem. Sci. 2012, 7, 11. [Google Scholar]
- Patil, S.A.; Hägerhäll, C.; Gorton, L. Electron transfer mechanisms between microorganisms and electrodes in bioelectrochemical systems. Bioanal. Rev. 2012, 4, 159–192. [Google Scholar] [CrossRef]
- Cao, Y.; Li, X.; Li, F.; Song, H. CRISPRi–sRNA: Transcriptional–Translational Regulation of Extracellular Electron Transfer in Shewanellaoneidensis. ACS Synth. Biol. 2017, 6, 1679–1690. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Dhar, B.R.; An, J.; Rittmann, B.E.; Ryu, H.; Santo Domingo, J.W.; Ren, H.; Chae, J. The Roles of Biofilm Conductivity and Donor Substrate Kinetics in a Mixed-Culture Biofilm Anode. Environ. Sci. Technol. 2016, 50, 12799–12807. [Google Scholar] [CrossRef] [PubMed]
- Croese, E.; Pereira, M.A.; Euverink, G.-J.W.; Stams, A.J.M.; Geelhoed, J.S. Analysis of the microbial community of the biocathode of a hydrogen-producing microbial electrolysis cell. Appl. Microbiol. Biotechnol. 2011, 92, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Squier, T.C.; Zachara, J.M.; Fredrickson, J.K. Respiration of metal (hydr)oxides by Shewanella and Geobacter: A key role for multihaem c-type cytochromes. Mol. Microbiol. 2007, 65, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Gregory, K.B.; Lovley, D.R. Remediation and Recovery of Uranium from Contaminated Subsurface Environments with Electrodes. Environ. Sci. Technol. 2005, 39, 8943–8947. [Google Scholar] [CrossRef]
- Zuo, Y.; Xing, D.; Regan, J.M.; Logan, B.E. Isolation of the exoelectrogenic bacterium Ochrobactrumanthropi YZ-1 by using a U-tube microbial fuel cell. Appl. Environ. Microbiol. 2008, 74, 3130–3137. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.E.; Newman, D.K. Extracellular electron transfer. Cell. Mol. Life Sci. CMLS 2001, 58, 1562–1571. [Google Scholar] [CrossRef]
- Rabaey, K.; Boon, N.; Höfte, M.; Verstraete, W. Microbial phenazine production enhances electron transfer in biofuel cells. Environ. Sci. Technol. 2005, 39, 3401–3408. [Google Scholar] [CrossRef]
- Kadier, A.; Simayi, Y.; Abdeshahian, P.; Azman, N.F.; Chandrasekhar, K.; Kalil, M.S. A comprehensive review of microbial electrolysis cells (MEC) reactor designs and configurations for sustainable hydrogen gas production. Alex. Eng. J. 2016, 55, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Zhao, Q.; Zhang, J.; Jiang, J.; Wan, C.; Du, M.; Zhao, S. A graphite-granule membrane-less tubular air-cathode microbial fuel cell for power generation under continuously operational conditions. J. Power Sources 2007, 173, 172–177. [Google Scholar] [CrossRef]
- Sun, R.; Zhou, A.; Jia, J.; Liang, Q.; Liu, Q.; Xing, D.; Ren, N. Characterization of methane production and microbial community shifts during waste activated sludge degradation in microbial electrolysis cells. Bioresour. Technol. 2014, 175, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Sugnaux, M.; Happe, M.; Cachelin, C.P.; Gloriod, O.; Huguenin, G.; Blatter, M.; Fischer, F. Two stage bioethanol refining with multi litre stacked microbial fuel cell and microbial electrolysis cell. Bioresour. Technol. 2016, 221, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Gil-Carrera, L.; Escapa, A.; Carracedo, B.; Morán, A.; Gómez, X. Performance of a semi-pilot tubular microbial electrolysis cell (MEC) under several hydraulic retention times and applied voltages. Bioresour. Technol. 2013, 146, 63–69. [Google Scholar] [CrossRef]
- Clauwaert, P.; Verstraete, W. Methanogenesis in membraneless microbial electrolysis cells. Appl. Microbiol. Biotechnol. 2009, 82, 829–836. [Google Scholar] [CrossRef]
- Feng, Q.; Song, Y.-C. Surface Modification of a Graphite Fiber Fabric Anode for Enhanced Bioelectrochemical Methane Production. Energy Fuels 2016, 30, 6467–6474. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Quan, X.; Chen, S. Enhanced anaerobic digestion of waste activated sludge digestion by the addition of zero valent iron. Water Res. 2013, 52, 242–250. [Google Scholar] [CrossRef]
- Asztalos, J.R.; Kim, Y. Enhanced digestion of waste activated sludge using microbial electrolysis cells at ambient temperature. Water Res. 2015, 87, 503–512. [Google Scholar] [CrossRef]
- Blasco-Gómez, R.; Batlle-Vilanova, P.; Villano, M.; Balaguer, M.D.; Colprim, J.; Puig, S. On the Edge of Research and Technological Application: A Critical Review of Electromethanogenesis. Int. J. Mol. Sci. 2017, 18, 874. [Google Scholar] [CrossRef]
- Kadier, A.; Simayi, Y.; Kalil, M.S.; Abdeshahian, P.; Hamid, A.A. A review of the substrates used in microbial electrolysis cells (MECs) for producing sustainable and clean hydrogen gas. Renew. Energy 2014, 71, 466–472. [Google Scholar] [CrossRef]
- Hu, K.; Chen, W.; Jia, S.; Wang, W.; Han, F. Enhanced Degradation of Waste Activated Sludge in Microbial Electrolysis Cell by Ultrasonic Treatment. Front. Microbiol. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Gajaraj, S.; Huang, Y.; Zheng, P.; Hu, Z. Methane production improvement and associated methanogenic assemblages in bioelectrochemically assisted anaerobic digestion. Biochem. Eng. J. 2017, 117, 105–112. [Google Scholar] [CrossRef]
- Siegert, M.; Yates, M.D.; Call, D.F.; Zhu, X.; Spormann, A.; Logan, B.E. Comparison of Nonprecious Metal Cathode Materials for Methane Production by Electromethanogenesis. ACS Sustain. Chem. Eng. 2014, 2, 910–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, B.; Tian, D.; Jun, H. Bioelectrochemical enhancement of methane production from highly concentrated food waste in a combined anaerobic digester and microbial electrolysis cell. Bioresour. Technol. 2018, 247, 226–233. [Google Scholar] [CrossRef]
- Su, M.; Wei, L.; Qiu, Z.; Jia, Q.; Shen, J. A graphene modified biocathode for enhancing hydrogen production. RSC Adv. 2015, 5, 32609–32614. [Google Scholar] [CrossRef]
- Yan, H.; Catania, C.; Bazan, G.C. Membrane-intercalating conjugated oligoelectrolytes: Impact on bioelectrochemical systems. Adv. Mater. 2015, 27, 2958–2973. [Google Scholar] [CrossRef]
- Jourdin, L.; Grieger, T.; Monetti, J.; Flexer, V.; Freguia, S.; Lu, Y.; Chen, J.; Romano, M.; Wallace, G.G.; Keller, J. High Acetic Acid Production Rate Obtained by Microbial Electrosynthesis from Carbon Dioxide. Environ. Sci. Technol. 2015, 49, 13566–13574. [Google Scholar] [CrossRef]
- Jourdin, L.; Freguia, S.; Donose, B.C.; Chen, J.; Wallace, G.G.; Keller, J.; Flexer, V. A novel carbon nanotube modified scaffold as an efficient biocathode material for improved microbial electrosynthesis. J. Mater. Chem. A 2014, 2, 13093–13102. [Google Scholar] [CrossRef] [Green Version]
- Choi, O.; Kim, T.; Woo, H.M.; Um, Y. Electricity-driven metabolic shift through direct electron uptake by electroactive heterotroph Clostridium pasteurianum. Sci. Rep. 2014, 4, 6961. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.R.; Myers, J.M. Localization of cytochromes to the outer membrane of anaerobically grown Shewanella putrefaciens MR-1. J. Bacteriol. 1992, 174, 3429–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Shi, L.; Brown, R.N.; Xiong, Y.; Fredrickson, J.K.; Romine, M.F.; Marshall, M.J.; Lipton, M.S.; Beyenal, H. Extracellular polymeric substances from Shewanella sp. HRCR-1 biofilms: Characterization by infrared spectroscopy and proteomics. Environ. Microbiol. 2011, 13, 1018–1031. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Yang, Y.; Sun, G.; Wu, D. Impact of applied voltage on methane generation and microbial activities in an anaerobic microbial electrolysis cell (MEC). Chem. Eng. J. 2016, 283, 260–265. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, B.; Yin, C.; Zhang, C.; Dai, X.; Yuan, H.; Zhu, N. Biostimulation by direct voltage to enhance anaerobic digestion of waste activated sludge. RSC Adv. 2015, 6, 1581–1588. [Google Scholar] [CrossRef]
- Zhen, G.; Kobayashi, T.; Lu, X.; Kumar, G.; Xu, K. Biomethane recovery from Egeria densa in a microbial electrolysis cell-assisted anaerobic system: Performance and stability assessment. Chemosphere 2016, 149, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gildemyn, S.; Verbeeck, K.; Slabbinck, R.; Andersen, S.J.; Prévoteau, A.; Rabaey, K. Integrated Production, Extraction, and Concentration of Acetic Acid from CO2 through Microbial Electrosynthesis. Environ. Sci. Technol. Lett. 2015, 2, 325–328. [Google Scholar] [CrossRef]
- Lai, B.; Yu, S.; Bernhardt, P.V.; Rabaey, K.; Virdis, B.; Krömer, J.O. Anoxic metabolism and biochemical production in Pseudomonas putida F1 driven by a bioelectrochemical system. Biotechnol. Biofuels 2016, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Freguia, S.; Dennis, P.G.; Keller, J.; Rabaey, K. Development of bioelectrocatalytic activity stimulates mixed-culture reduction of glycerol in a bioelectrochemical system. Microb. Biotechnol. 2015, 8, 483–489. [Google Scholar] [CrossRef]
- ElMekawy, A.; Hegab, H.M.; Losic, D.; Saint, C.P.; Pant, D. Applications of graphene in microbial fuel cells: The gap between promise and reality. Renew. Sustain. Energy Rev. 2017, 72, 1389–1403. [Google Scholar] [CrossRef]
- Moscoviz, R.; Trably, E.; Bernet, N. Electro-fermentation triggering population selection in mixed-culture glycerol fermentation. Microb. Biotechnol. 2017, 11, 74–83. [Google Scholar] [CrossRef]
- Farid, M.A.; El-Enshasy, H.A.; El-Deen, A.M.N. Alcohol production from starch by mixed cultures of Aspergillus awamori and immobilized Saccharomyces cerevisiae at different agitation speeds. J. Basic Microbiol. 2002, 42, 162–171. [Google Scholar] [CrossRef]
- Palanisamy, K. State of the art and future concept of food waste fermentation to bioenergy. Renew. Sustain. Energy Rev. 2016, 53, 547–557. [Google Scholar]
- Zhao, Z.; Zhang, Y.; Wang, L.; Quan, X. Potential for direct interspecies electron transfer in an electric-anaerobic system to increase methane production from sludge digestion. Sci. Rep. 2015, 5, 11094. [Google Scholar] [CrossRef] [PubMed]
- Hafez, H.M. Method and System for Electro-assisted Hydrogen Production from Organic Material. U.S. Patent 9,458,474, 4 October 2016. [Google Scholar]
- Selembo, P.A.; Perez, J.M.; Lloyd, W.A.; Logan, B.E. High hydrogen production from glycerol or glucose by electrohydrogenesis using microbial electrolysis cells. Int. J. Hydrogen Energy 2009, 34, 5373–5381. [Google Scholar] [CrossRef]
- Butanol Production from Food Waste: A Novel Process for Producing Sustainable Energy and Reducing Environmental Pollution Biotechnology for Biofuels Full Text. Available online: https://biotechnologyforbiofuels.biomedcentral.com/articles/10.1186/s13068-015-0332-x (accessed on 19 June 2020).
- Lee, J.W.; Kim, H.U.; Choi, S.; Yi, J.; Lee, S.Y. Microbial production of building block chemicals and polymers. Curr. Opin. Biotechnol. 2011, 22, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Ammu, B.; Binod, P.; Deepthi, S.K.; Ramachandran, K.B.; Soccol, C.R.; Pandey, A. Production and characterization of poly-3-hydroxybutyrate from crude glycerol by Bacillus sphaericus NII 0838 and improving its thermal properties by blending with other polymers. Braz. Arch. Biol. Technol. 2011, 54, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Cok, B.; Tsiropoulos, I.; Roes, A.L.; Patel, M.K. Succinic acid production derived from carbohydrates: An energy and greenhouse gas assessment of a platform chemical toward a bio-based economy. Biofuels Bioprod. Biorefin. 2014, 8, 16–29. [Google Scholar] [CrossRef]
- Choi, O.; Um, Y.; Sang, B.-I. Butyrate production enhancement by Clostridium tyrobutyricum using electron mediators and a cathodic electron donor. Biotechnol. Bioeng. 2012, 109, 2494–2502. [Google Scholar] [CrossRef]
- Park, D.H.; Zeikus, J.G. Utilization of Electrically Reduced Neutral Red by Actinobacillus succinogenes: Physiological Function of Neutral Red in Membrane-Driven Fumarate Reduction and Energy Conservation. J. Bacteriol. 1999, 181, 2403–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.S.; Kim, B.H. Electron flow shift inClostridiumacetobutylicum fermentation by electrochemically introduced reducing equivalent. Biotechnol. Lett. 1988, 10, 123–128. [Google Scholar] [CrossRef]
- Schmitz, S.; Nies, S.; Wierckx, N.; Blank, L.M.; Rosenbaum, M.A. Engineering mediator-based electroactivity in the obligate aerobic bacterium Pseudomonas putida KT2440. Front. Microbiol. 2015, 6, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, A.V.; Mahadevan, R. In silico characterization of microbial electrosynthesis for metabolic engineering of biochemicals. Microb. Cell Factories 2011, 10, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, A.S.; Wang, J.; Luo, J.; Yau, S.-T. Enhanced ethanol production via electrostatically accelerated fermentation of glucose using Saccharomyces cerevisiae. Sci. Rep. 2015, 5, 15713. [Google Scholar] [CrossRef] [Green Version]
- Deval, A.S.; Parikh, H.A.; Kadier, A.; Chandrasekhar, K.; Bhagwat, A.M.; Dikshit, A.K. Sequential microbial activities mediated bioelectricity production from distillery wastewater using bio-electrochemical system with simultaneous waste remediation. Int. J. Hydrogen Energy 2017, 42, 1130–1141. [Google Scholar] [CrossRef]
- Thygesen, A.; Thomsen, A.B.; Possemiers, S.; Verstraete, W. Integration of microbial electrolysis cells (MECs) in the biorefinery for production of ethanol, H2 and phenolics. Waste Biomass Valoriz. 2010, 1, 9–20. [Google Scholar] [CrossRef]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Schaap, J.D.; Kampman, C.; Buisman, C.J.N. Bioelectrochemical ethanol production through mediated acetate reduction by mixed cultures. Environ. Sci. Technol. 2010, 44, 513–517. [Google Scholar] [CrossRef]
- Lee, W.S.; Chua, A.S.M.; Yeoh, H.K.; Ngoh, G.C. A review of the production and applications of waste-derived volatile fatty acids. Chem. Eng. J. 2013, 235, 83–99. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Mohanakrishna, G.; Goud, R.K.; Sarma, P.N. Acidogenic fermentation of vegetable based market waste to harness biohydrogen with simultaneous stabilization. Bioresour. Technol. 2009, 100, 3061–3068. [Google Scholar] [CrossRef]
- Sarkar, O.; Venkata Mohan, S. Deciphering acidogenic process towards biohydrogen, biohythane, and short chain fatty acids production: Multi-output optimization strategy. Biofuel Res. J. 2016, 3, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yan, B.; Wong, J.W.C.; Zhang, Y. Enhanced volatile fatty acids production from anaerobic fermentation of food waste: A mini-review focusing on acidogenic metabolic pathways. Bioresour. Technol. 2018, 248, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Gildemyn, S.; Molitor, B.; Usack, J.G.; Nguyen, M.; Rabaey, K.; Angenent, L.T. Upgrading syngas fermentation effluent using Clostridium kluyveri in a continuous fermentation. Biotechnol. Biofuels 2017, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eerten-Jansen, M.C.A.A.; Ter Heijne, A.; Grootscholten, T.I.M.; Steinbusch, K.J.J.; Sleutels, T.H.J.A.; Hamelers, H.V.M.; Buisman, C.J.N. Bioelectrochemical Production of Caproate and Caprylate from Acetate by Mixed Cultures. ACS Sustain. Chem. Eng. 2013, 1, 513–518. [Google Scholar] [CrossRef]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Plugge, C.M.; Buisman, C.J.N. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass. Energy Environ. Sci. 2010, 4, 216–224. [Google Scholar] [CrossRef]
- Kucek, L.A.; Xu, J.; Nguyen, M.; Angenent, L.T. Waste Conversion into n-Caprylate and n-Caproate: Resource Recovery from Wine Lees Using Anaerobic Reactor Microbiomes and In-line Extraction. Front. Microbiol. 2016, 7, 1892. [Google Scholar] [CrossRef] [Green Version]
- Grootscholten, T.I.M.; dal Borgo, F.K.; Hamelers, H.V.M.; Buisman, C.J.N. Promoting chain elongation in mixed culture acidification reactors by addition of ethanol. Biomass Bioenergy 2013, 48, 10–16. [Google Scholar] [CrossRef]
- Logan, B.E.; Rabaey, K. Conversion of wastes into bioelectricity and chemicals by using microbial electrochemical technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.W.-C.; Tyagi, R.D.; Pandey, A. Current Developments in Biotechnology and Bioengineering: Solid Waste Management; Elsevier Science: Amsterdam, The Netherlands, 2016; ISBN 978-0-444-63664-5. [Google Scholar]
- Montpart, N.; Rago, L.; Baeza, J.A.; Guisasola, A. Hydrogen production in single chamber microbial electrolysis cells with different complex substrates. Water Res. 2015, 68, 601–615. [Google Scholar] [CrossRef]
- Pasupuleti, S.B.; Sarkar, O.; Mohan, S.V. Upscaling of biohydrogen production process in semi-pilot scale biofilm reactor: Evaluation with food waste at variable organic loads. Int. J. Hydrogen Energy 2014, 39, 7587–7596. [Google Scholar] [CrossRef]
- Schröder, U.; Harnisch, F.; Angenent, L.T. Microbial electrochemistry and technology: Terminology and classification. Energy Environ. Sci. 2015, 8, 513–519. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, J.; Yu, Q.; Ma, W.; Quan, X.; Zhang, Y. Enhanced decomposition of waste activated sludge via anodic oxidation for methane production and bioenergy recovery. Int. Biodeterior. Biodegrad. 2016, 106, 161–169. [Google Scholar] [CrossRef]
- Van Steendam, C.; Smets, I.; Skerlos, S.; Raskin, L. Improving anaerobic digestion via direct interspecies electron transfer requires development of suitable characterization methods. Curr. Opin. Biotechnol. 2019, 57, 183–190. [Google Scholar] [CrossRef]
- Cheng, S.; Xing, D.; Call, D.F.; Logan, B.E. Direct biological conversion of electrical current into methane by electromethanogenesis. Environ. Sci. Technol. 2009, 43, 3953–3958. [Google Scholar] [CrossRef]
- Liu, H.; Grot, S.; Logan, B.E. Electrochemically Assisted Microbial Production of Hydrogen from Acetate. Environ. Sci. Technol. 2005, 39, 4317–4320. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.-E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef]
- Freguia, S.; Rabaey, K.; Yuan, Z.; Keller, J. Syntrophic Processes Drive the Conversion of Glucose in Microbial Fuel Cell Anodes. Environ. Sci. Technol. 2008, 42, 7937–7943. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Modestra, J.A.; Amulya, K.; Butti, S.K.; Velvizhi, G. A Circular Bioeconomy with Biobased Products from CO2 Sequestration. Trends Biotechnol. 2016, 34, 506–519. [Google Scholar] [CrossRef]
- Butti, S.K.; Mohan, S.V. Autotrophic biorefinery: Dawn of the gaseous carbon feedstock. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Modestra, J.A.; Babu, M.C.L.; Mohan, S.V. Electro-fermentation of real-field acidogenic spent wash effluents for additional biohydrogen production with simultaneous treatment in a microbial electrolysis cell. Sep. Purif. Technol. 2015. [Google Scholar] [CrossRef]
- Villano, M.; Aulenta, F.; Ciucci, C.; Ferri, T.; Giuliano, A.; Majone, M. Bioelectrochemical reduction of CO2 to CH4 via direct and indirect extracellular electron transfer by a hydrogenophilic methanogenic culture. Bioresour. Technol. 2010, 101, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Su, M. Production of Acetate from Carbon Dioxide in Bioelectrochemical Systems Based on Autotrophic Mixed Culture. J. Microbiol. Biotechnol. 2013, 23, 1140–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganigué, R.; Puig, S.; Batlle-Vilanova, P.; Balaguer, M.D.; Colprim, J. Microbial electrosynthesis of butyrate from carbon dioxide. Chem. Commun. Camb. Engl. 2015, 51, 3235–3238. [Google Scholar] [CrossRef] [PubMed]
- Kadier, A.; Simayi, Y.; Chandrasekhar, K.; Ismail, M.; Kalil, M.S. Hydrogen gas production with an electroformed Ni mesh cathode catalysts in a single-chamber microbial electrolysis cell (MEC). Int. J. Hydrogen Energy 2015, 40, 14095–14103. [Google Scholar] [CrossRef]
- Kumar, G.; Saratale, R.G.; Kadier, A.; Sivagurunathan, P.; Zhen, G.; Kim, S.H.; Saratale, G.D. A review on bio-electrochemical systems (BESs) for the syngas and value added biochemicals production. Chemosphere 2017, 177, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rozendal, R.A.; Hamelers, H.V.M.; Euverink, G.J.W.; Metz, S.J.; Buisman, C.J.N. Principle and perspectives of hydrogen production through biocatalyzed electrolysis. Int. J. Hydrogen Energy 2006, 31, 1632–1640. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Iversen, S.L.; Sørensen, K.I.; Johansen, E. The long and winding road from the research laboratory to industrial applications of lactic acid bacteria. FEMS Microbiol. Rev. 2005, 29, 611–624. [Google Scholar] [CrossRef]
- Lechardeur, D.; Cesselin, B.; Fernandez, A.; Lamberet, G.; Garrigues, C.; Pedersen, M.; Gaudu, P.; Gruss, A. Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 2011, 22, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.; Kashefi, K.; Blunt-Harris, E.L.; Lovley, D.R. Microbiological evidence for Fe(III) reduction on early Earth. Nature 1998, 395, 65–67. [Google Scholar] [CrossRef]
- Rozendal, R.A.; Jeremiasse, A.W.; Hamelers, H.V.M.; Buisman, C.J.N. Hydrogen Production with a Microbial Biocathode. Environ. Sci. Technol. 2008, 42, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Kadier, A.; Jain, P.; Lai, B.; Kalil, M.S.; Kondaveeti, S.; Alabbosh, K.F.S.; Abu-Reesh, I.M.; Mohanakrishna, G. Biorefinery perspectives of microbial electrolysis cells (MECs) for hydrogen and valuable chemicals production through wastewater treatment. Biofuel Res. J. 2020, 7, 1128–1142. [Google Scholar] [CrossRef] [Green Version]
- Venkata Mohan, S.; Mohanakrishna, G.; Sarma, P.N. Integration of acidogenic and methanogenic processes for simultaneous production of biohydrogen and methane from wastewater treatment. Int. J. Hydrogen Energy 2008, 33, 2156–2166. [Google Scholar] [CrossRef]
- Rathinam, N.K.; Bibra, M.; Salem, D.R.; Sani, R.K. Thermophiles for biohydrogen production in microbial electrolytic cells. Bioresour. Technol. 2019, 277, 171–178. [Google Scholar] [CrossRef]
- Shemfe, M.; Gadkari, S.; Yu, E.; Rasul, S.; Scott, K.; Head, I.M.; Gu, S.; Sadhukhan, J. Life cycle, techno-economic and dynamic simulation assessment of bioelectrochemical systems: A case of formic acid synthesis. Bioresour. Technol. 2018, 255, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Gil-Carrera, L.; Escapa, A.; Mehta, P.; Santoyo, G.; Guiot, S.R.; Morán, A.; Tartakovsky, B. Microbial electrolysis cell scale-up for combined wastewater treatment and hydrogen production. Bioresour. Technol. 2013, 130, 584–591. [Google Scholar] [CrossRef]
- Heidrich, E.S.; Edwards, S.R.; Dolfing, J.; Cotterill, S.E.; Curtis, T.P. Performance of a pilot scale microbial electrolysis cell fed on domestic wastewater at ambient temperatures for a 12month period. Bioresour. Technol. 2014, 173, 87–95. [Google Scholar] [CrossRef] [Green Version]
- San-Martín, M.I.; Sotres, A.; Alonso, R.M.; Díaz-Marcos, J.; Morán, A.; Escapa, A. Assessing anodic microbial populations and membrane ageing in a pilot microbial electrolysis cell. Int. J. Hydrogen Energy 2019, 44, 17304–17315. [Google Scholar] [CrossRef]
- Redwood, M.D.; Orozco, R.L.; Majewski, A.J.; Macaskie, L.E. Electro-extractive fermentation for efficient biohydrogen production. Bioresour. Technol. 2012, 107, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Rabaey, K.; Verstraete, W. Microbial fuel cells: Novel biotechnology for energy generation. Trends Biotechnol. 2005, 23, 291–298. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Kuppam, C.; Mudhoo, A.; Saratale, G.D.; Kadier, A.; Zhen, G.; Chatellard, L.; Trably, E.; Kumar, G. A comprehensive review on two-stage integrative schemes for the valorization of dark fermentative effluents. Crit. Rev. Biotechnol. 2018, 38, 868–882. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr.No. | Substrate | Applied Voltage (V) | Product Synthesized | Production Rate | Electrofermentation | Microbial Catalyst | References |
|---|---|---|---|---|---|---|---|
| 1 | Food waste | −0.6 V | Acetate, butyrate, andpropionate | 4595 mg/L d | Anodic | Mixed microbial culture | [84] |
| 2 | CO2 | −0.9 V Ecat | CH4 | 9.2 L/m2 d | Cathodic | Mixed enriched culture biocatalyst | [145] |
| 3 | Acetate | 0.5 V | H2 | 0.1 L/m2d | Anodic | Mixed enriched culture biocatalyst | [127] |
| 4 | CO2 | −0.9 to −1.1 | H2 and acetate | 0.38 mM d | Cathodic | Mixed culture domestic wastewater treatment plant sludge | [146] |
| 5 | CO2 | þ0.197 V/SHE | Butyrate, ethanol, and butanol | 1.82 MMC/d | Cathodic | Clostridium | [147] |
| Cathode Surface Area (cm2) | V (L) | σ(S·m−1) | Anolyte | Current Density (A·m−2) | U (V) | Gas Production (L·d−1·m−2) | Energy Yields (–) | Faradaic Yields (%) | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH4 | H2 | γH | γG | An. | Cath H2 | |||||||
| 3024 | 5 × 2 | 0.7 | Wastewater | 0.61 | 1.08 | yes | 1.0 | 0.22 | 0.19 | 19 | 23 | [146] |
| 3360 | 120 | 0.17 | Wastewater acetate | 0.3 max | 1.1 | 0.06 | 3.6 | NA | 0.69 | 16 | 55 | [147] |
| 840 | 2 × 2 | 0.05 | Wastewater | 0.24 | 1.0 | 0.23 | 1.06 | 1.1 | 0.5 | NA | 44 | [146] |
| 4466 | 33 | NA | Sewage sludge | 0.01 | 2.0 | 0.28 | 0 | 6.45 | 0 | NA | NA | [88] |
| 1800 | 16 | 2.6 | Pig slurry, acetate | 1.75 | 1.0 | <2% | 17.8 | 1.5 | 1.25 | 7–9 | 78 | [148] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhagchandanii, D.D.; Babu, R.P.; Sonawane, J.M.; Khanna, N.; Pandit, S.; Jadhav, D.A.; Khilari, S.; Prasad, R. A Comprehensive Understanding of Electro-Fermentation. Fermentation 2020, 6, 92. https://doi.org/10.3390/fermentation6030092
Bhagchandanii DD, Babu RP, Sonawane JM, Khanna N, Pandit S, Jadhav DA, Khilari S, Prasad R. A Comprehensive Understanding of Electro-Fermentation. Fermentation. 2020; 6(3):92. https://doi.org/10.3390/fermentation6030092
Chicago/Turabian StyleBhagchandanii, Drishti Dinesh, Rishi Pramod Babu, Jayesh M. Sonawane, Namita Khanna, Soumya Pandit, Dipak A. Jadhav, Santimoy Khilari, and Ram Prasad. 2020. "A Comprehensive Understanding of Electro-Fermentation" Fermentation 6, no. 3: 92. https://doi.org/10.3390/fermentation6030092
APA StyleBhagchandanii, D. D., Babu, R. P., Sonawane, J. M., Khanna, N., Pandit, S., Jadhav, D. A., Khilari, S., & Prasad, R. (2020). A Comprehensive Understanding of Electro-Fermentation. Fermentation, 6(3), 92. https://doi.org/10.3390/fermentation6030092