SSR-Marker Analysis—A Method for S. cerevisiae Strain Characterization and Its Application for Wineries

Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.1.1. Commercially Available Saccharomyces Cerevisiae Strains
2.1.2. Wine Samples
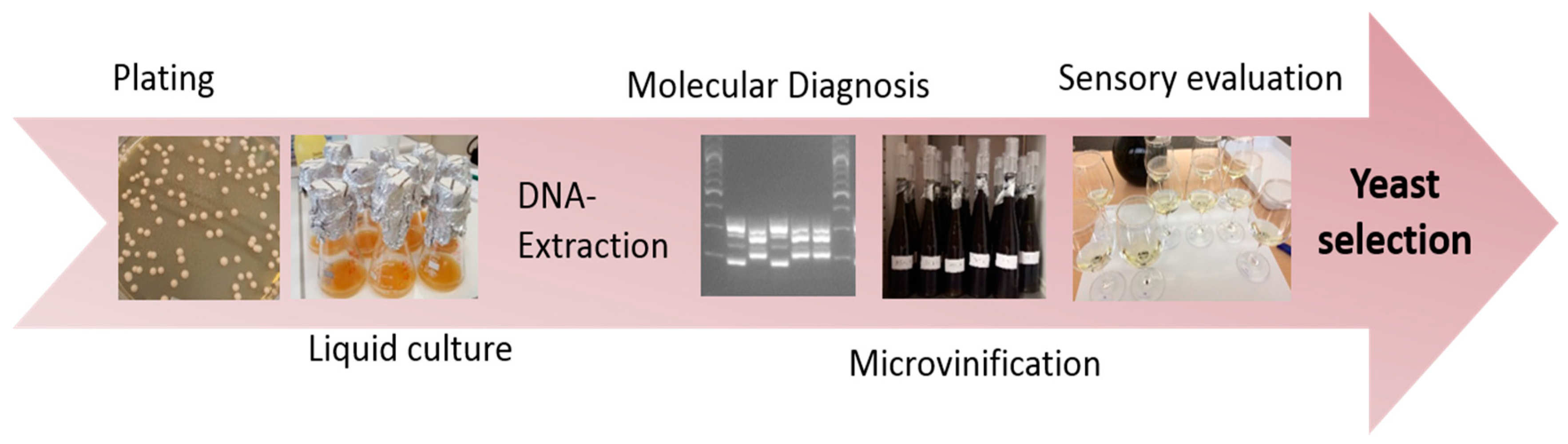
2.1.3. Yeast Isolation and Preservation
2.2. Methods:
2.2.1. DNA Extraction
2.2.2. SSR-PCR
2.2.3. Fragment Length Determination by Agarose Gel Electrophoresis
2.2.4. Capillary Sequencing and Method Validation
2.2.5. Fermentation and Sensory Tests
3. Results
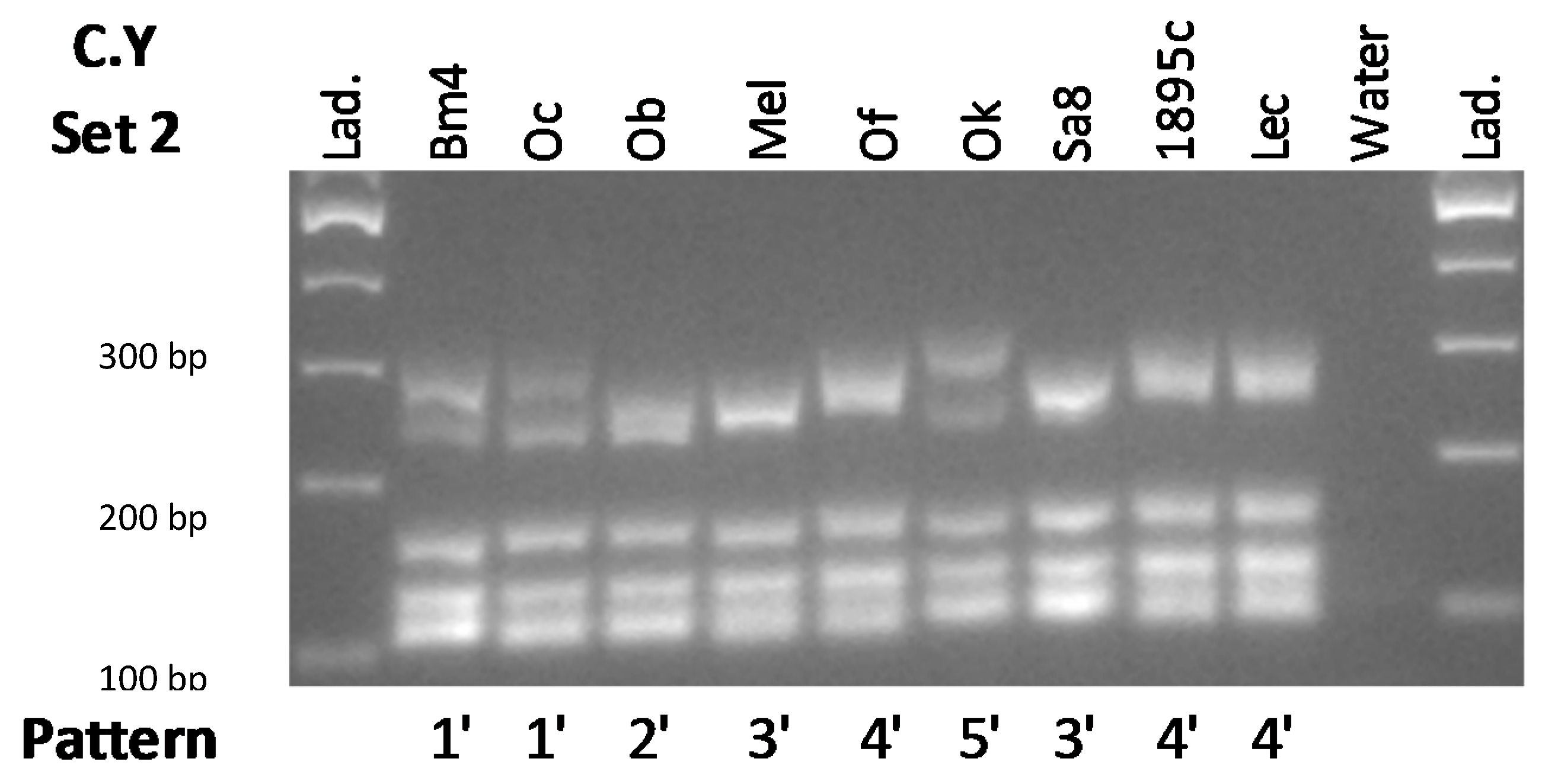
3.1. Primer Sets’ Composition and Method Validation
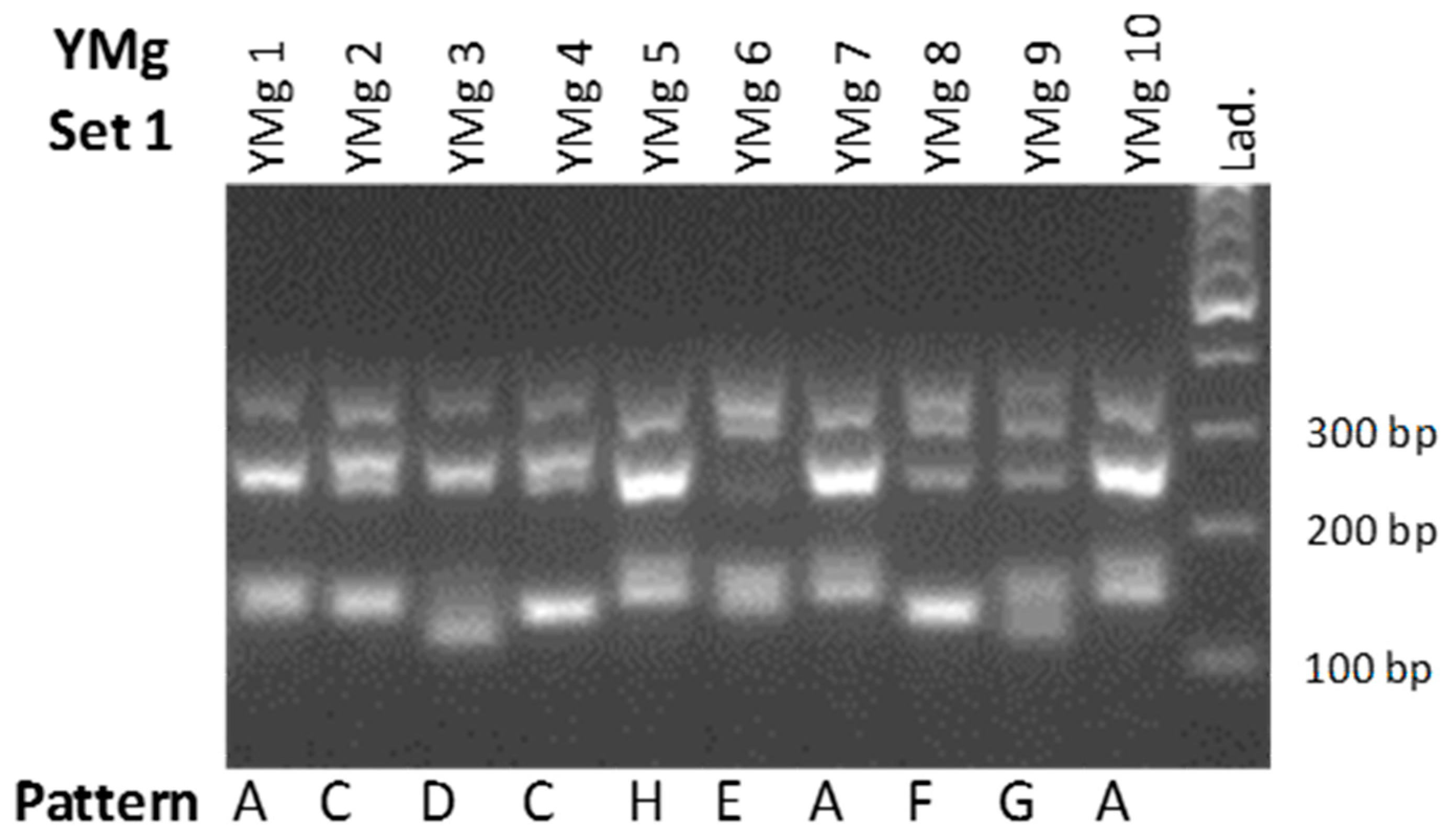
3.2. Yeast Differentiation
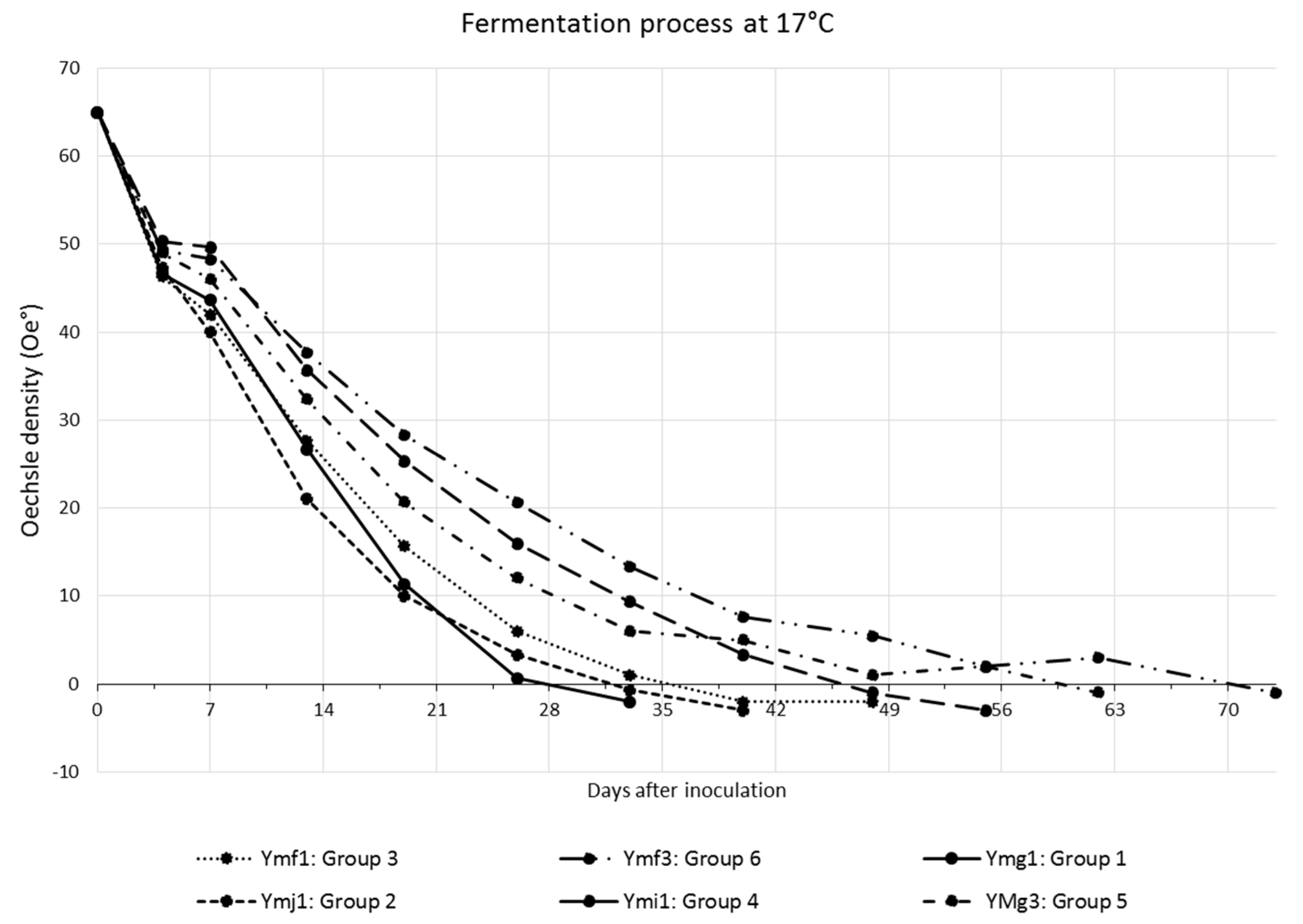
3.3. Fermentative Tests
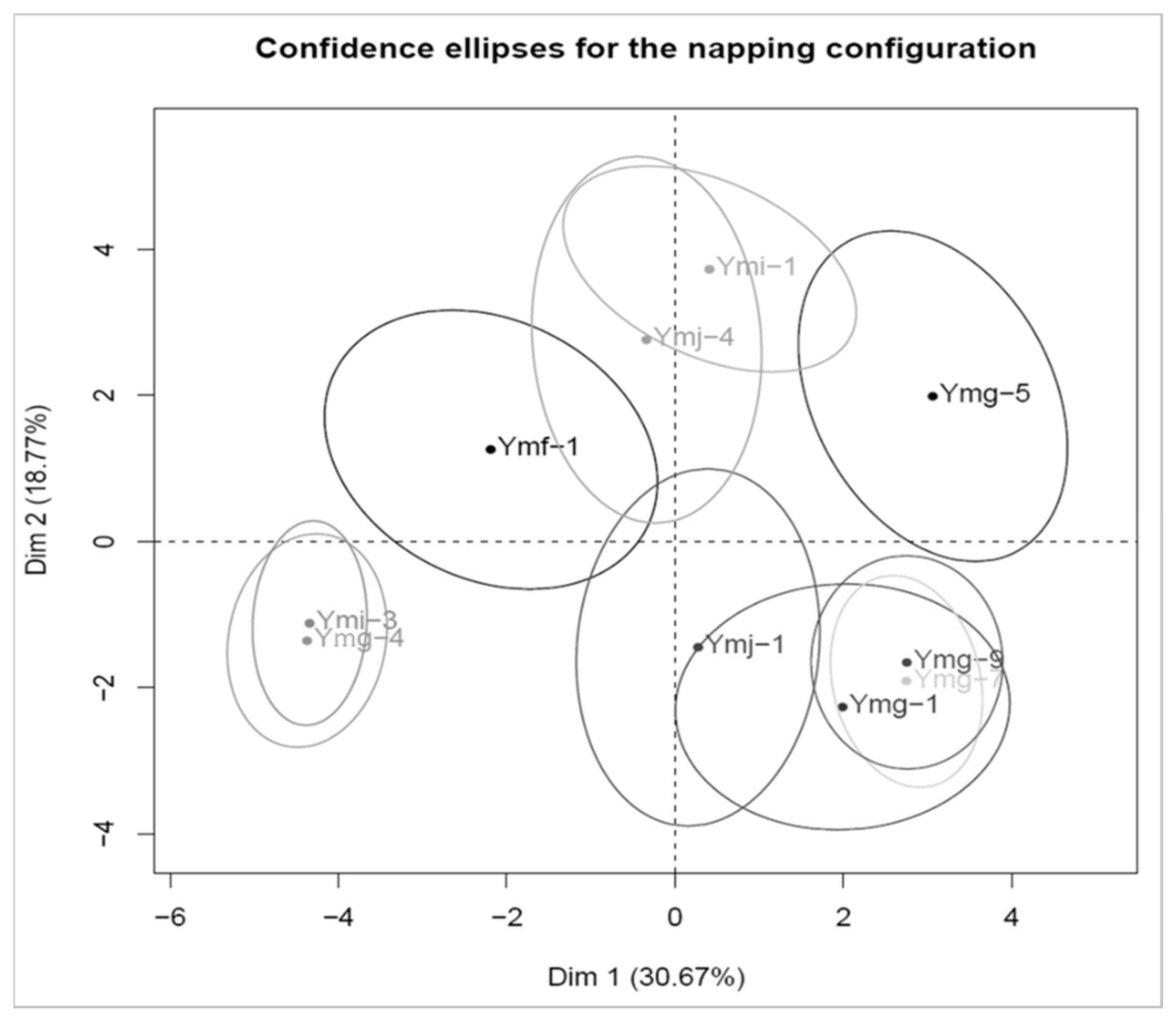
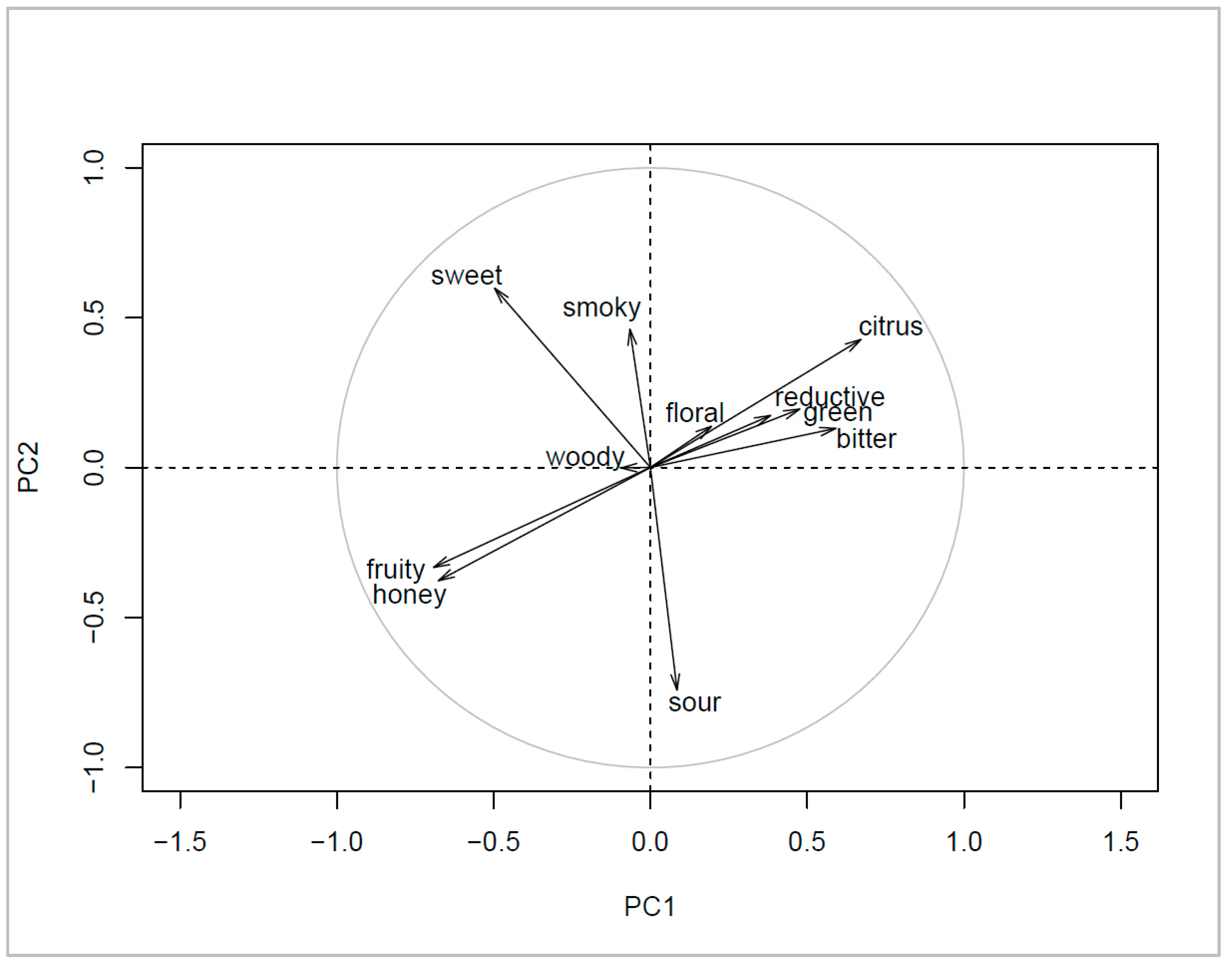
3.4. Sensory Differences
4. Discussion
4.1. Method Validation and Primer Set Composition
4.2. Winery’s Own Yeast Differentiation
4.3. Fermentative Tests
4.4. Sensory Differences
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chambers, A.H.; Evans, S.A.; Folta, K.M. Methyl Anthranilate and Gamma-Decalactone Inhibit Strawberry Pathogen Growth and Achene Germination. J. Agric. Food Chem. 2013, 61, 12625–12633. [Google Scholar] [CrossRef] [PubMed]
- Nykänen, L. Formation and Occurrence of Flavor Compounds in Wine and Distilled Alcoholic Beverages. Am. J. Enol. Vitic. 1986, 37, 84–96. [Google Scholar]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. S. Afr. J. Enol. Vitic. 2019, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- van Uden, N. Chapter 2—Ethanol Toxicity and Ethanol Tolerance in Yeasts. In Annual Reports on Fermentation Processes; Tsao, G.T., Ed.; Elsevier: Munich, Germany, 1985; pp. 11–58. [Google Scholar]
- D’Amore, T.; Panchal, C.J.; Russell, I.; Stewart, G.G. A study of ethanol tolerance in yeast. Crit. Rev. Biotechnol. 1989, 9, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Bandara, A.; Fraser, S.; Chambers, P.J.; Stanley, G.A. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J. Appl. Microbiol. 2010, 109, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Romano, P. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Techera, A.G.; Jubany, S.; Carrau, F.; Gaggero, C. Differentiation of industrial wine yeast strains using microsatellite markers. Lett. Appl. Microbiol. 2001, 33, 71–75. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Pretorius, I.S. Microbial formation and modification of flavor and off-flavor compounds in wine. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Heidelberg, Germany, 2009; pp. 209–231. [Google Scholar]
- Alexandre, H. Wine Yeast Terroir: Separating the Wheat from the Chaff—for an Open Debate. Microorganisms 2020, 8, 787. [Google Scholar] [CrossRef]
- Merle, A.; Piotrowski, M. Consommer des produits alimentaires locaux—Comment et pourquoi? Décisions Mark. 2012, 67, 37–48. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous Food Fermentations and Potential Risks for Human Health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Erusso, P.; Ecapozzi, V.; Espano, G.; Ecorbo, M.R.; Esinigaglia, M.; Bevilacqua, A. Metabolites of Microbial Origin with an Impact on Health: Ochratoxin A and Biogenic Amines. Front. Microbiol. 2016, 7, 482. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Russo, P. Microbiological Safety and the Management of Microbial Resources in Artisanal Foods and Beverages: The Need for a Transdisciplinary Assessment to Conciliate Actual Trends and Risks Avoidance. Microorganisms 2020, 8, 306. [Google Scholar] [CrossRef] [Green Version]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 Genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, P.; Jeffreys, A.; Wilson, V.; Blanchetot, A. Organization of the human myoglobin gene. EMBO J. 1984, 3, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Wills, C. Abundant microsatellite polymorphism in Saccharomyces cerevisiae, and the different distributions of microsatellites in eight prokaryotes and S. cerevisiae, result from strong mutation pressures and a variety of selective forces. Proc. Natl. Acad. Sci. USA 1998, 95, 1647–1652. [Google Scholar] [CrossRef] [Green Version]
- Jarne, P.; Lagoda, P.J. Microsatellites, from molecules to populations and back. Trends Ecol. Evol. 1996, 11, 424–429. [Google Scholar] [CrossRef]
- Albertin, W.; Panfili, A.; Miot-Sertier, C.; Goulielmakis, A.; Delcamp, A.; Salin, F.; Lonvaud-Funel, A.; Curtin, C.; Masneuf-Pomarede, I. Development of microsatellite markers for the rapid and reliable genotyping of Brettanomyces bruxellensis at strain level. Food Microbiol. 2014, 42, 188–195. [Google Scholar] [CrossRef]
- Ghislain, M.; Spooner, D.M.; Rodríguez, F.; Villamón, F.; Núñez, J.; Vásquez, C.; Waugh, R.; Bonierbale, M. Selection of highly informative and user-friendly microsatellites (SSRs) for genotyping of cultivated potato. Theor. Appl. Genet. 2003, 108, 881–890. [Google Scholar] [CrossRef]
- Garofalo, C.; Berbegal, C.; Grieco, F.; Tufariello, M.; Spano, G.; Capozzi, V. Selection of indigenous yeast strains for the production of sparkling wines from native Apulian grape varieties. Int. J. Food Microbiol. 2018, 285, 7–17. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The Biodiversity of Saccharomyces cerevisiae in Spontaneous Wine Fermentation: The Occurrence and Persistence of Winery-Strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Tufariello, M.; Maiorano, G.; Rampino, P.; Spano, G.; Grieco, F.; Perrotta, C.; Capozzi, V. Selection of an autochthonous yeast starter culture for industrial production of Primitivo “Gioia del Colle” PDO/DOC in Apulia (Southern Italy). LWT 2019, 99, 188–196. [Google Scholar] [CrossRef]
- Vaudano, E.; Garcia-Moruno, E. Discrimination of Saccharomyces cerevisiae wine strains using microsatellite multiplex PCR and band pattern analysis. Food Microbiol. 2008, 25, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.-L.; Ruh, O.; Merdinoglu, D.; Karst, F. Selection of hypervariable microsatellite loci for the characterization of Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2005, 102, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.L.; Masneuf-Pomarede, I. Cellar-Associated Saccharomyces cerevisiae Population Structure Revealed High-Level Diversity and Perennial Persistence at Sauternes Wine Estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- Pages, J. Recueil direct de distances sensorielles: Application à l’évaluation de dix vins blancs du Val-de-Loire. Sci. Aliment. 2003, 23, 679–688. [Google Scholar] [CrossRef]
- Pagès, J. Collection and analysis of perceived product inter-distances using multiple factor analysis: Application to the study of 10 white wines from the Loire Valley. Food Qual. Prefer. 2005, 16, 642–649. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: AnRPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hennequin, C.; Thierry, A.; Richard, G.F.; Lecointre, G.; Nguyen, H.V.; Gaillardin, C.; Dujon, B. Microsatellite Typing as a New Tool for Identification of Saccharomyces cerevisiae Strains. J. Clin. Microbiol. 2001, 39, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Sihlhavy, K.; Berger, S.U.; Mandl, K.A.; Hack, R.O.; Regner, F.E. Microsatellite analysis of commercial wine yeast strains. Mitt. Klosterneubg. 2006, 56, 140–146. [Google Scholar]
- Torija, M.J. Effects of fermentation temperature on the strain population of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 80, 47–53. [Google Scholar] [CrossRef]
- Iranzo, J.U.; Magaña, F.G.; Viñas, M.G. Evaluation of the formation of volatiles and sensory characteristics in the industrial production of white wines using different commercial strains of the genus Saccharomyces. Food Control 2000, 11, 143–147. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Commercial Name | Abbreviation | Origin |
|---|---|---|
| Lalvin BM4*4 | Bm4 | Lallemand, Fredericia, Denmark |
| Oenoferm Color | Oc | Erbslöh, Geisenheim, Germany |
| Oenoferm Bouquet | Ob | Erbslöh, Geisenheim, Germany |
| Oenoferm Klosterneuburg | Ok | Erbslöh, Geisenheim, Germany |
| Oenoferm Freddo | Of | Erbslöh, Geisenheim, Germany |
| Melody | Mel | Chr Hansen, Hoersholm, Denmark |
| Siha Activhefe 8 | Sa8 | Eaton Technologies, Langenlonsheim, Germany |
| 1895c | 1895c | Swiss Wineyeast GmbH, Meilen, Switzerland |
| Lalvin EC 1118 | Lec | Lallemand, Fredericia, Denmark |
| Primer | Motif | ORF Coordinates | Sequences |
|---|---|---|---|
| ScAAT2 | TAA | YBL084c | FW:CAGTCTTATTGCCTTGAACGA RV:GTCTCCATCCTCCAAACAGCC |
| ScAAt3 | TAA | YDR160w | FW:TGGGAGGAGGGAAATGGACAG RV:TTCAGTTACCCGCACAATCTA |
| C5 | GT | VI-210250/210414 | FW:TGACACAATAGCAATGGCCTTCA RV:GCAAGCGACTAGAACAACAATCACA |
| C3 | CAA | YGL139w | FW:CTTTTTATTTACGAGCGGGCCAT RV:AAATCTCATGCCTGTGAGGGGTAT |
| C8 | TAA | YGL014w | FW:CAGGTCGTTCTAACGTTGGTAAAATG RV:GCTGTTGCTGTTGGTAGCATTACTGT |
| C11 | GT | X-518870/519072 | FW:TTCCATCATAACCGTCTGGGATT RV:TGCCTTTTTCTTAGATGGGCTTTC |
| YKR072c | GAC | YKR072c | FW:AGATACAGAAGATAAGAACGAAAA RV:TTATTGATGCTTATCTATTATACC |
| SCYOR267c | TGT | YOR267c | FW:TACTAACGTCAACACTGCTGCCAA RV:GGATCTACTTGCAGTATACGGG |
| YKL172w | GAA | YKL172w | FW:CAGGACGCTACCGAAGCTCAAAAG RV:ACTTTTGGCCAATTTCTCAAGAT |
| ScAAT1 | TTA | XIII-86902/87140 | FW:AAGCGTAAGCAATGGTGTAGATACTT RV:CAAGCCTCTTCAAGCATGACCTTT |
| C4 | TAA + TAG | XV-110701/110935 | FW:AGGAGAAAAATGCTGTTTATTCTGACC RV:TTTTCCTCCGGGACGTGAAATA |
| C9 | TAA | YOR156c | FW:AAGGGTTCGTAAACATATAACTGGCA RV:TATAAGGGAAAAGAGCACGATGGC |
| ScAAT5 | TAA | XVI-897051/8970210 | FW:AGCATAATTGGAGGCAGTAAAGCA RV:TCTCCGTCTTTTTTGTACTGCGTG |
| C6 | CA | XVI-485898/485996 | FW:GTGGCATCATATCTGTCAATTTTATCAC RV:CAATCAAGCAAAAGATCGGCCT |
| YPL009c | CTT | YPL009c | FW:AACCCATTGACCTCGTTACTATCGT RV:TTCGATGGCTCTGATAACTCCATTC |
| Primer | Size Range by Agarose gel (bp) | Size Range by Capillary Sequencing (bp) | Primer Set |
|---|---|---|---|
| ScAAT2 | 310–390 | 376–385 | 1 |
| ScAAt3 | 260 | 244–274 | |
| C5 | 130–170 | / | |
| SCYOR267c | 305–390 | 305–325 | |
| C8 | 150 | 133–146 | |
| C11 | 200–250 | / | |
| YKL172w | 135 | 120–125 | 2 |
| C4 | 240–320 | 248–260 | |
| C9 | 100 | 90–100 | |
| ScAAT5 | 160–180 | 156–162 | |
| C6 | 100–120 | 100–102 | |
| YPL009c | 250–290 | 276–311 |
| Pattern | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| A | 18 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| B | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| C | 4 | 0 | 1 | 1 | 0 | 7 | 0 | 3 |
| D | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| E | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| F | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| G | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| H | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| I | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| J | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Group | Fermentation Time (days) | Yeasts | Total Number of Yeast |
|---|---|---|---|
| Group 1 | 33 | Ymg1 | 1 |
| Group 2 | 40 | Ymj1, Ymg4, Ymg7, Ymg9, Ymj4 | 5 |
| Group 3 | 48 | Ymf1, Ymg5, Ymi3 | 3 |
| Group 4 | 55 | Ymi1, Ymg2, Ymg8, Ymj2 | 4 |
| Group 5 | 62 | Ymg3, Ymh4 | 2 |
| Group 6 | 73 | Ymg6, Ymf3 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rex, F.; Hirschler, A.; Scharfenberger-Schmeer, M. SSR-Marker Analysis—A Method for S. cerevisiae Strain Characterization and Its Application for Wineries. Fermentation 2020, 6, 101. https://doi.org/10.3390/fermentation6040101
Rex F, Hirschler A, Scharfenberger-Schmeer M. SSR-Marker Analysis—A Method for S. cerevisiae Strain Characterization and Its Application for Wineries. Fermentation. 2020; 6(4):101. https://doi.org/10.3390/fermentation6040101
Chicago/Turabian StyleRex, Friederike, Adeline Hirschler, and Maren Scharfenberger-Schmeer. 2020. "SSR-Marker Analysis—A Method for S. cerevisiae Strain Characterization and Its Application for Wineries" Fermentation 6, no. 4: 101. https://doi.org/10.3390/fermentation6040101
APA StyleRex, F., Hirschler, A., & Scharfenberger-Schmeer, M. (2020). SSR-Marker Analysis—A Method for S. cerevisiae Strain Characterization and Its Application for Wineries. Fermentation, 6(4), 101. https://doi.org/10.3390/fermentation6040101