
Microbial and Chemical Analysis of Non-Saccharomyces Yeasts from Chambourcin Hybrid Grapes for Potential Use in Winemaking



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Grape Sampling and Juice Collection
2.2. Growth Media and Fungal Isolation
2.3. Molecular Identification of Fungal Isolates
2.4. Physiological Characterization of Non-Saccharomyces Yeasts
2.5. Laboratory Scale Fermentation
2.6. Analysis of Flavor Compounds of Fermented Chambourcin Juice
2.6.1. Analysis of Nonvolatile Compounds by UHPLC
2.6.2. Analysis of Volatile Compounds by Gas Chromatography-Mass Spectrometry
2.7. Statistical Analysis
3. Results
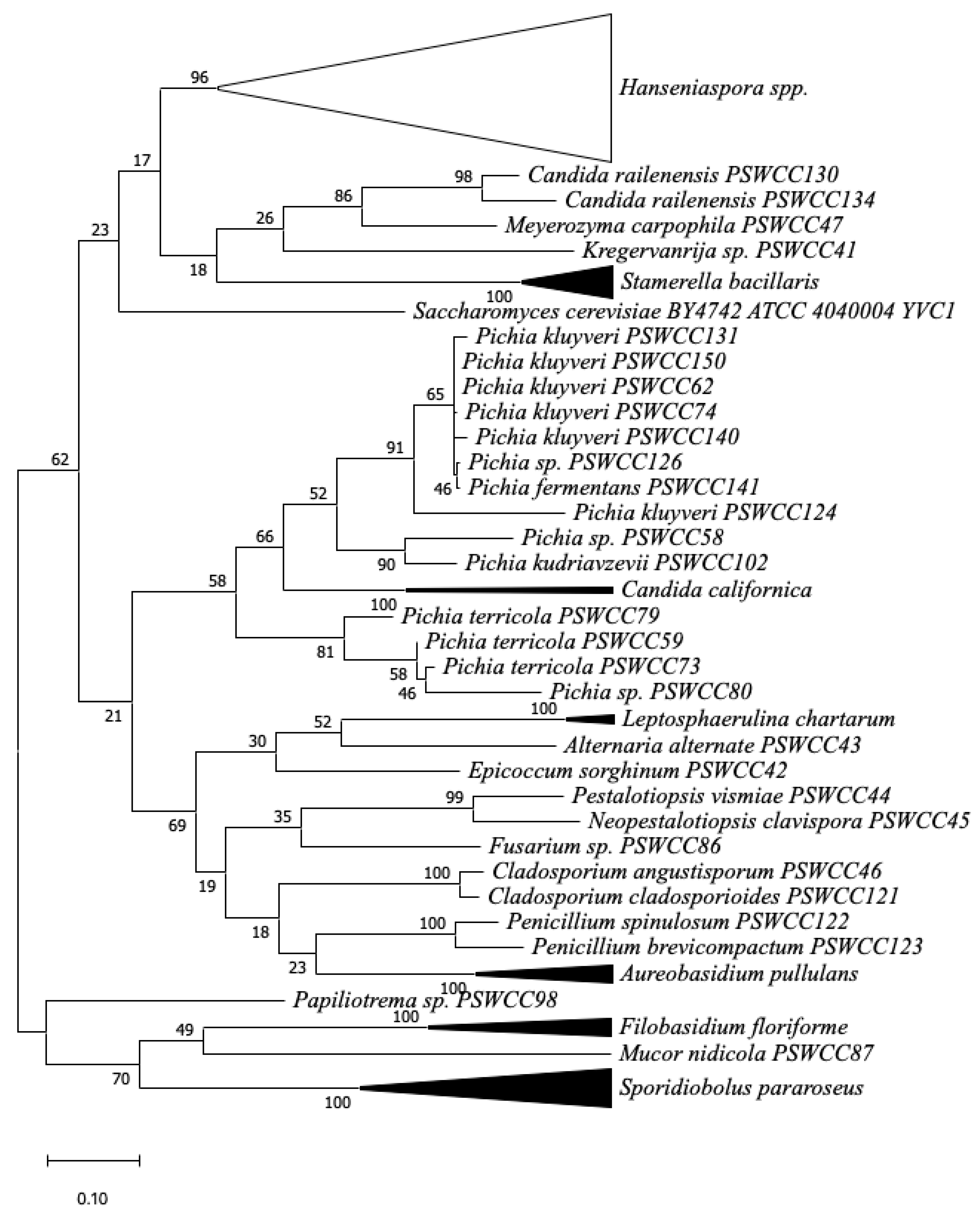
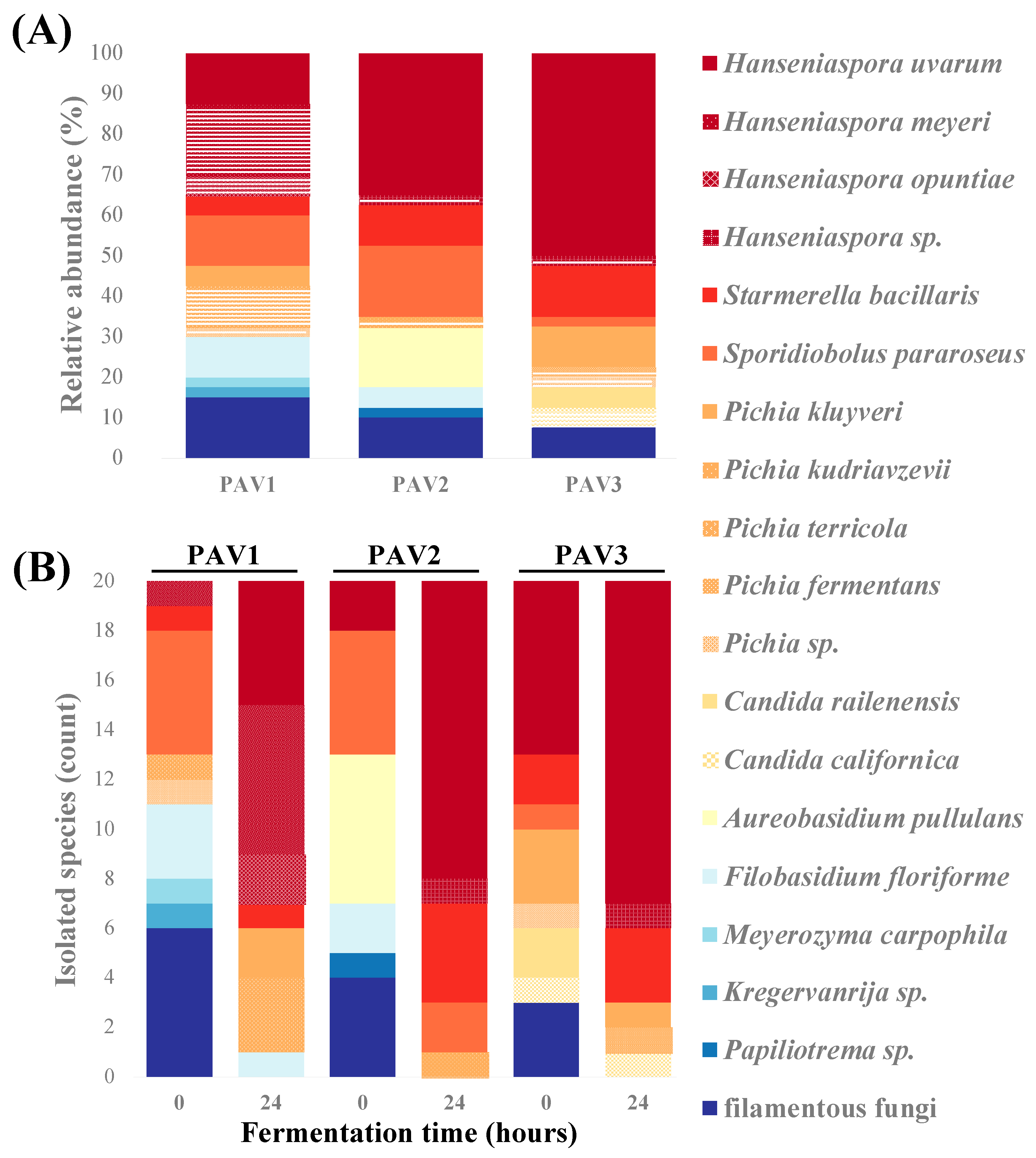
3.1. Spontaneous Fermentation of Chambourcin Grape Must Provides Insights into Fungal Diversity of Red Hybrid Grapes
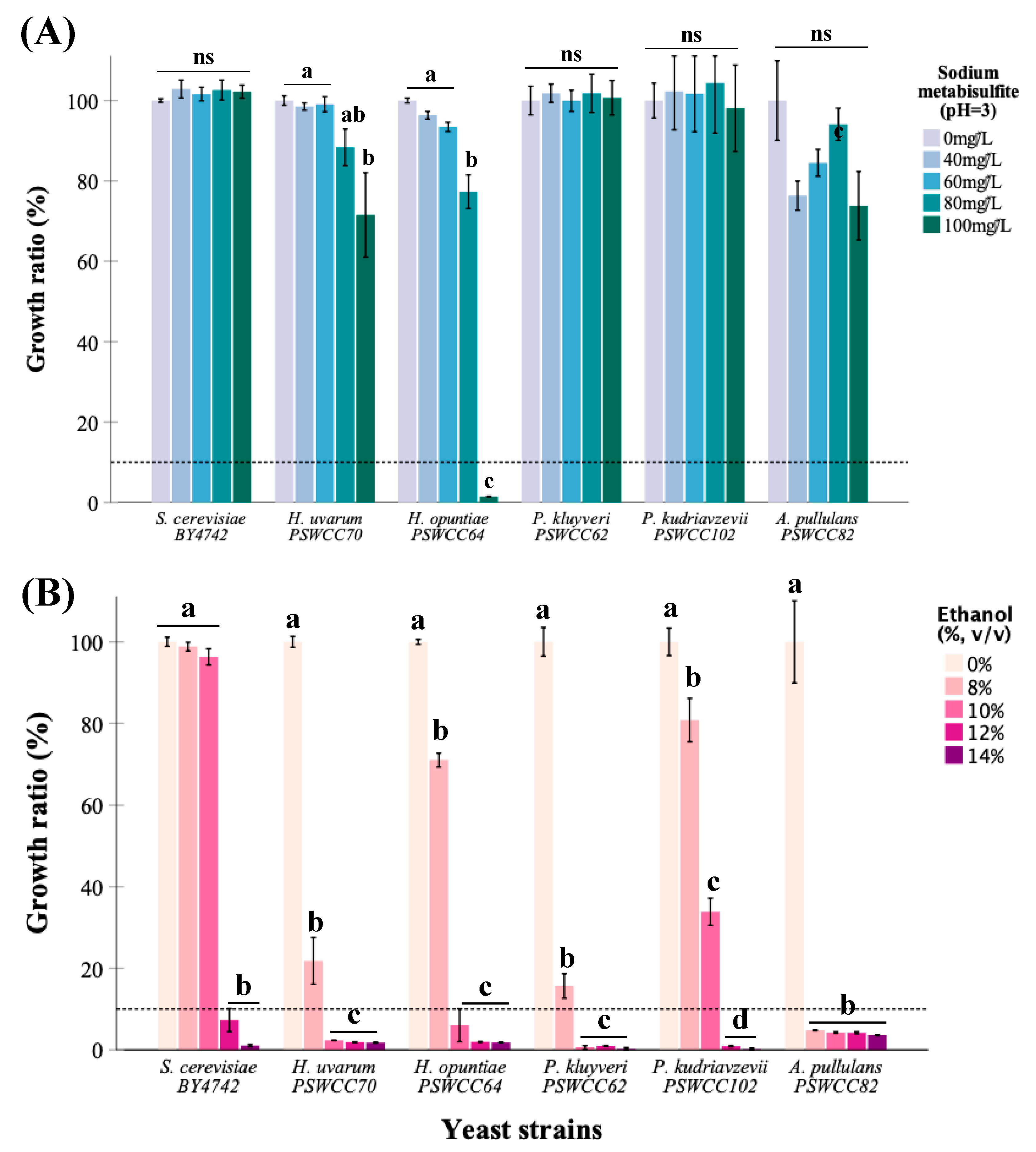
3.2. Physiological Characterization of Non-Saccharomyces Yeasts
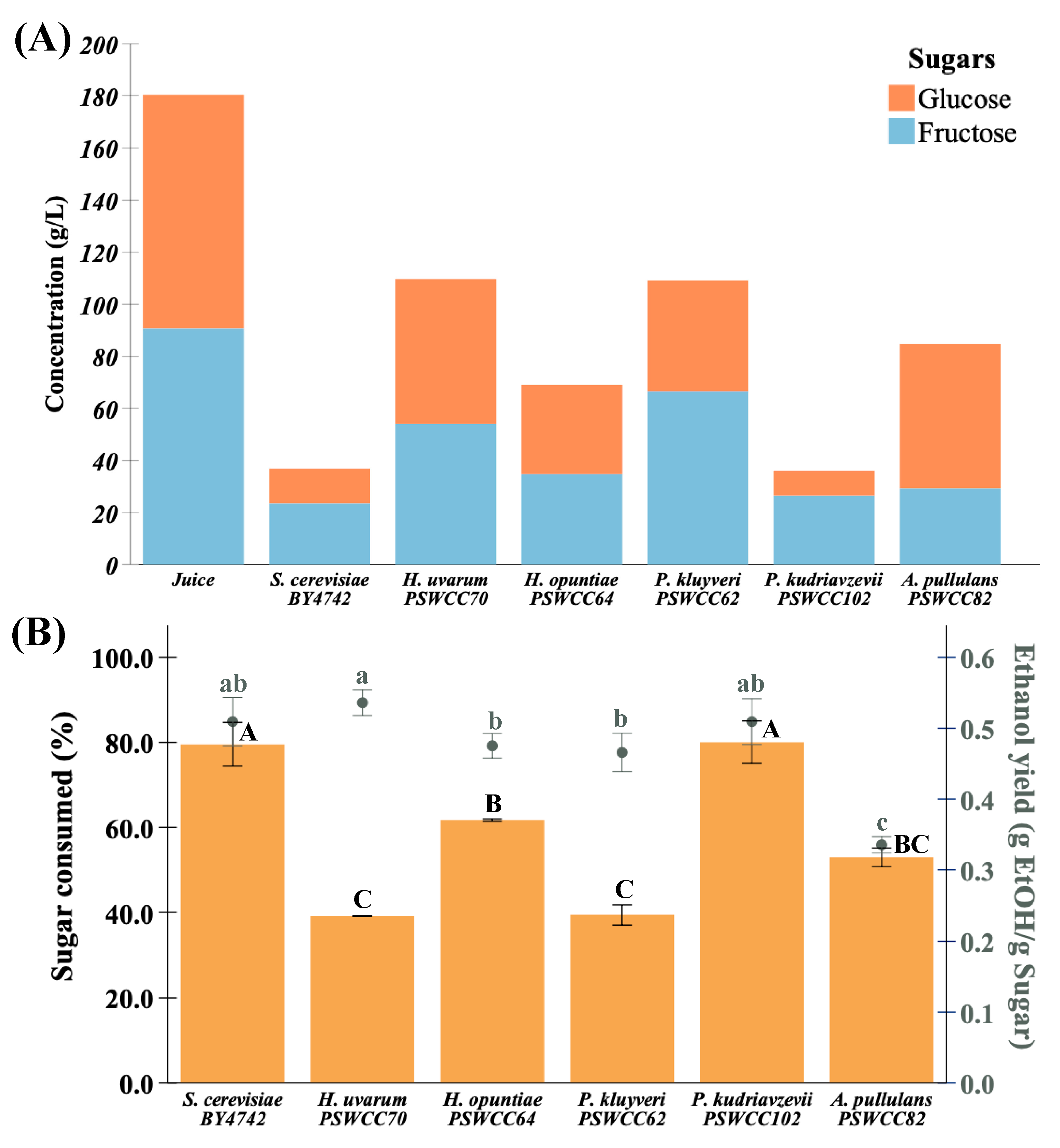
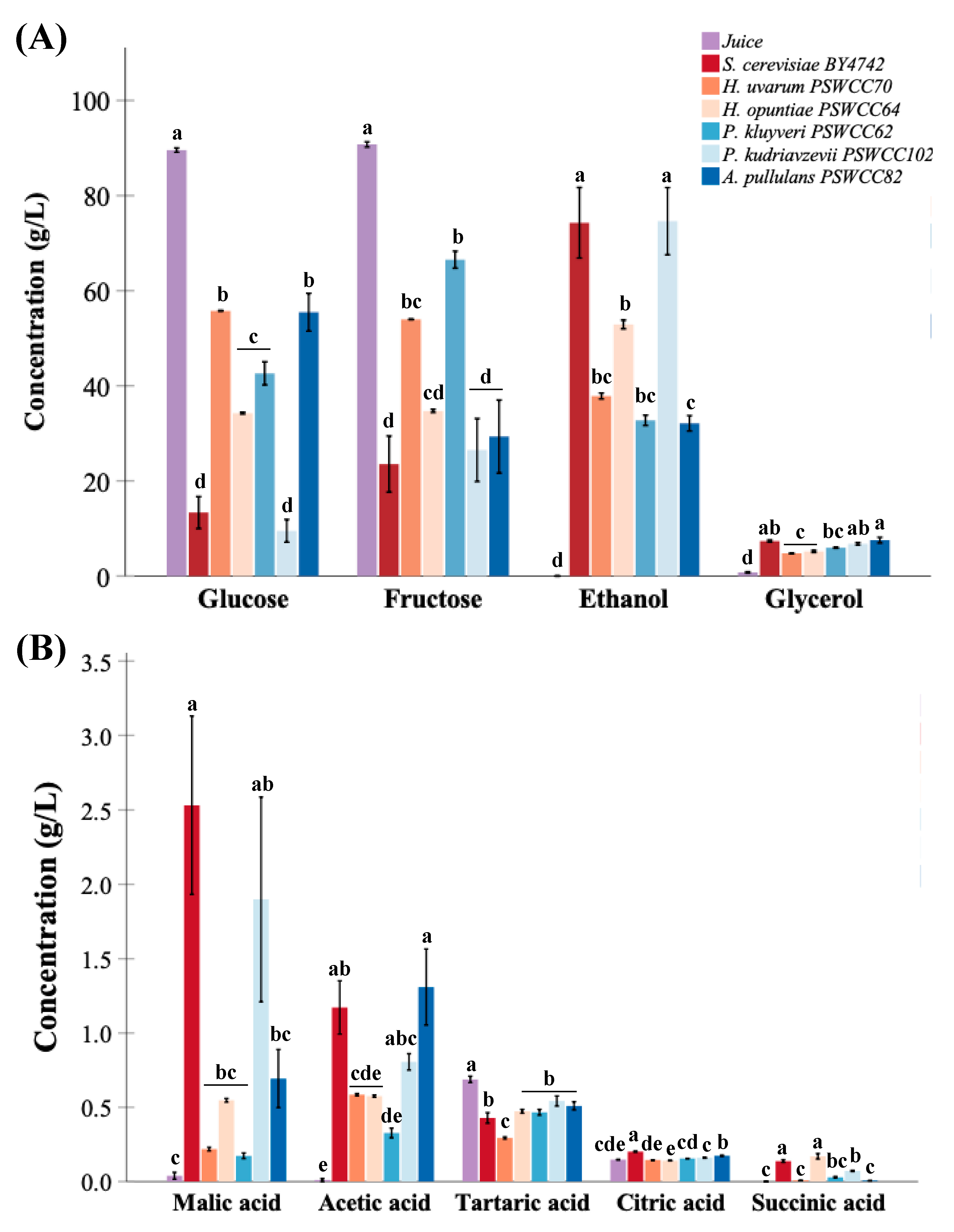
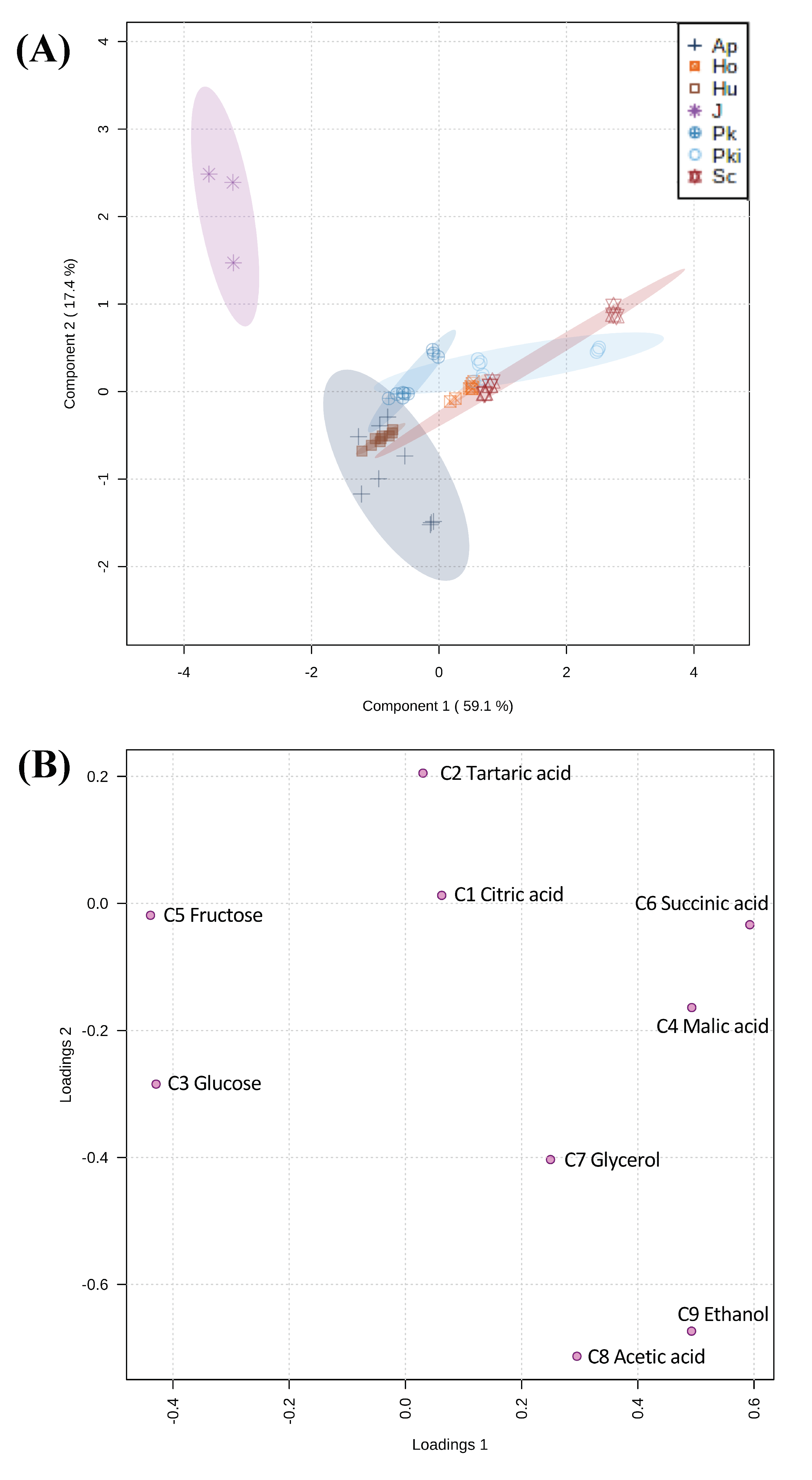
3.3. Characterization of Core Nonvolatile Compounds Highlight Fermentative Potential of Candidate Non-Saccharomyces Yeasts
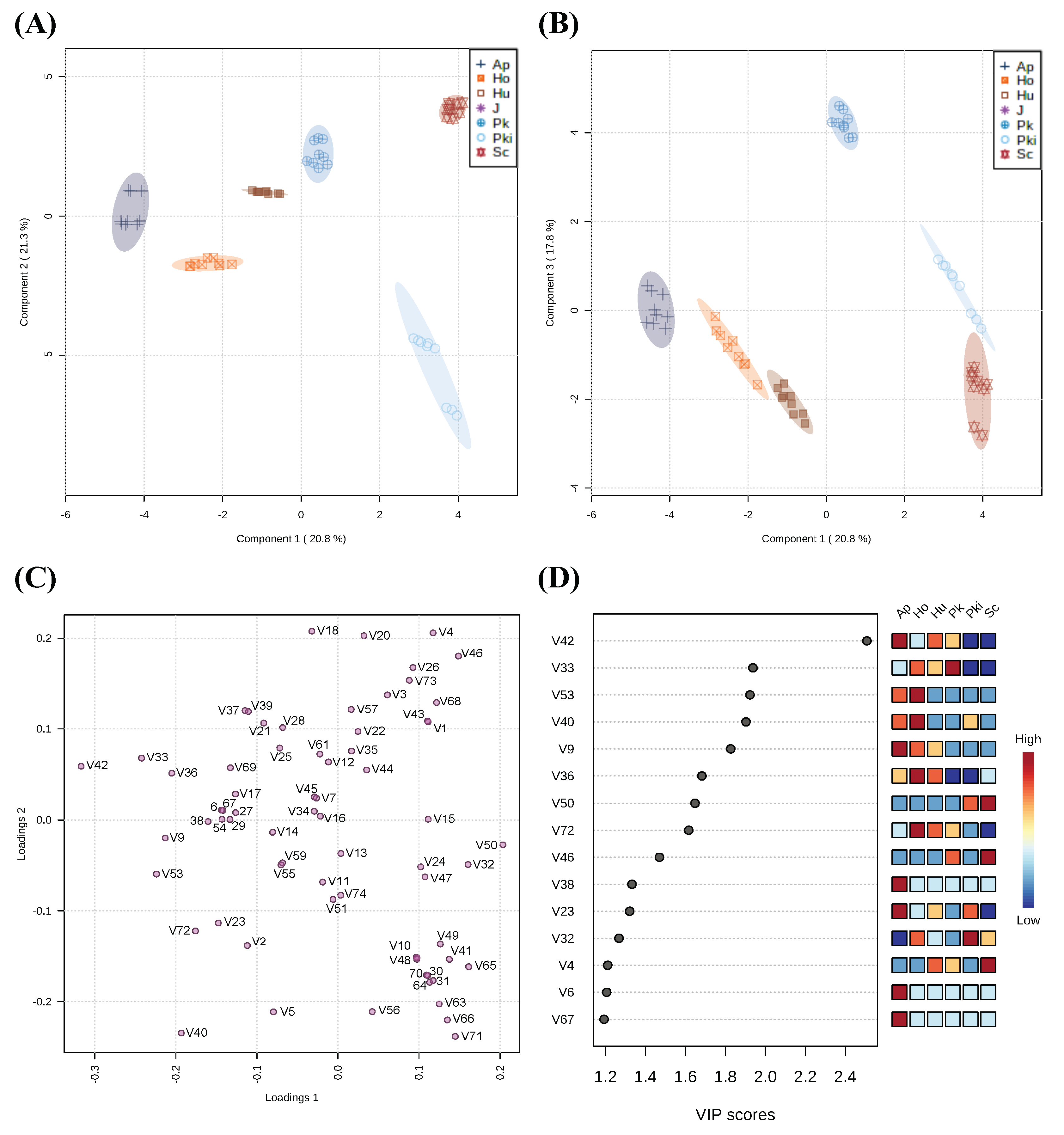
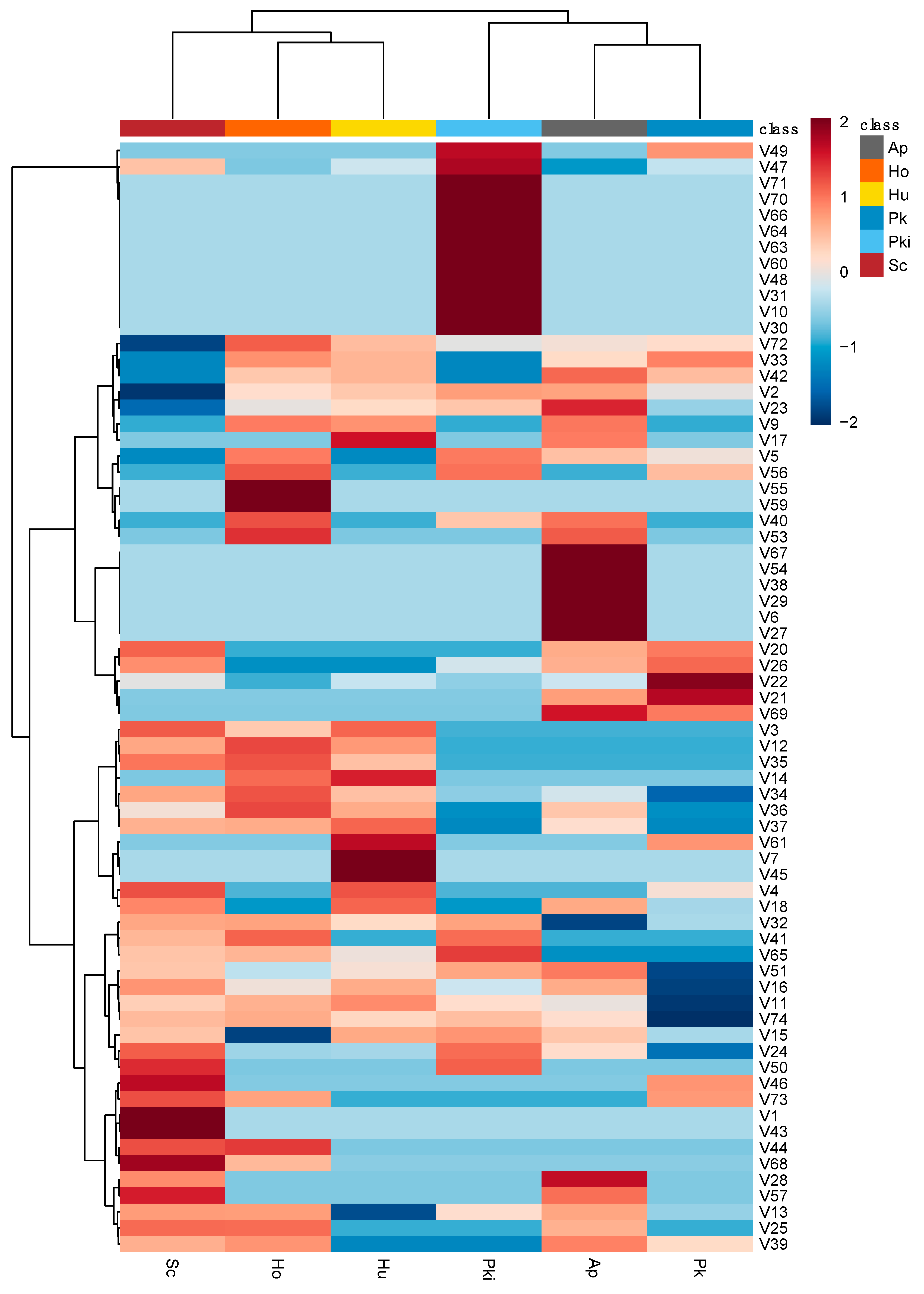
3.4. Distinct Fermentation-Derived Volatile Metabolites Provide Insights into Unique Properties of Candidate Non-Saccharomyces Yeast from Chambourcin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Legras, J.-L.; Merdinoglu, D.; Cornuet, J.-M.; Karst, F. Bread, Beer and Wine: Saccharomyces Cerevisiae Diversity Reflects Human History. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.A.; Echavarri-Erasun, C. Yeast Biotechnology. In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 21–44. ISBN 9780444521491. [Google Scholar]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction Of Alcohol Content in Wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemos Junior, W.J.F.; Nadai, C.; Crepalde, L.T.; De Oliveira, V.S.; De Matos, A.D.; Giacomini, A.; Corich, V. Potential Use of Starmerella Bacillaris as Fermentation Starter for the Production of Low-Alcohol Beverages Obtained from Unripe Grapes. Int. J. Food Microbiol. 2019, 303, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gamero, A.; Quintilla, R.; Groenewald, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-Throughput Screening of a Large Collection of Non-Conventional Yeasts Reveals Their Potential for Aroma Formation in Food Fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine Aroma Evolution Throughout Alcoholic Fermentation Sequentially Inoculated with Non- Saccharomyces/Saccharomyces Yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; Del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of Non-Saccharomyces Yeasts and Oenological Tannin in Red Winemaking: Influence on Colour, Aroma and Sensorial Properties of Young Wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The Application of Non-Saccharomyces Yeast in Fermentations with Limited Aeration as a Strategy for the Production of Wine with Reduced Alcohol Content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Goold, H.D.; Kroukamp, H.; Williams, T.C.; Paulsen, I.T.; Varela, C.; Pretorius, I.S. Yeast’s Balancing Act Between Ethanol And Glycerol Production In Low-Alcohol Wines. Microb. Biotechnol. 2017, 10, 264–278. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The Oenological Potential of Hanseniaspora Uvarum In Simultaneous And Sequential Co-Fermentation With Saccharomyces Cerevisiae For Industrial Wine Production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Torchio, F.; Cravero, F.; Marengo, F.; Giacosa, S.; Gerbi, V.; Rantsiou, K.; Rolle, L.; Cocolin, L. Aroma Profile And Composition Of Barbera Wines Obtained By Mixed Fermentations Of Starmerella Bacillaris (Synonym Candida Zemplinina) And Saccharomyces Cerevisiae. LWT Food Sci. Technol. 2016, 73, 567–575. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution Of Non-Saccharomyces Yeasts To Wine Volatile And Sensory Diversity: A Study On Lachancea Thermotolerans, Metschnikowia Spp. And Starmerella Bacillaris Strains Isolated In Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S. Chemical Constituents of Grapes and Wine. In Wine Science; Elsevier: Amsterdam, The Netherlands, 2014; pp. 347–426. [Google Scholar]
- Vaudano, E.; Quinterno, G.; Costantini, A.; Pulcini, L.; Pessione, E.; Garcia-Moruno, E. Yeast Distribution In Grignolino Grapes Growing In A New Vineyard In Piedmont And The Technological Characterization Of Indigenous Saccharomyces spp. Strains. Int. J. Food Microbiol. 2019, 289, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleet, G.H. Wine Yeasts For The Future. In Proceedings of the Fems Yeast Research; Oxford Academic: Oxford, UK, 2008; Volume 8, pp. 979–995. [Google Scholar]
- Jones, N.K. The Influence of Recent Climate Change on Wine Regions in Quebec, Canada. J. Wine Res. 2012, 23, 103–113. [Google Scholar] [CrossRef]
- Pedneault, K.; Provost, C. Fungus Resistant Grape Varieties as a Suitable Alternative for Organic Wine Production: Benefits, Limits, and Challenges. Sci. Hortic. (Amst.) 2016, 208, 57–77. [Google Scholar] [CrossRef]
- Ghareib, M.; Youssef, K.A.; Khalil, A.A. Ethanol Tolerance of Saccharomyces Cerevisiae and Its Relationship to Lipid Content and Composition. Folia Microbiol. 1988, 33, 447–452. [Google Scholar] [CrossRef]
- Cray, J.A.; Bell, A.N.W.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The Biology of Habitat Dominance; Can Microbes Behave As Weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.W.; Oliver, S.G.; Harrison, D.E.F.; Righelato, R.C. Ethanol Inhibition of Yeast Growth and Fermentation: Differences in the Magnitude and Complexity of the Effect. Eur. J. Appl. Microbiol. Biotechnol. 1981, 11, 151–155. [Google Scholar] [CrossRef]
- Aguilera, F.; Peinado, R.A.; Millán, C.; Ortega, J.M.; Mauricio, J.C. Relationship Between Ethanol Tolerance, H+-Atpase Activity And The Lipid Composition Of The Plasma Membrane In Different Wine Yeast Strains. Int. J. Food Microbiol. 2006, 110, 34–42. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, D.; Lin, Y.; Wang, X.; Kong, H.; Tanaka, S. Substrate and Product Inhibition on Yeast Performance in Ethanol Fermentation. Energy Fuels 2015, 29, 1019–1027. [Google Scholar] [CrossRef]
- Lopez, E.F.; Gomez, E.F. Simultaneous Determination of the Major Organic Acids, Sugars, Glycerol, and Ethanol by Hplc in Grape Musts and White Wines. J. Chromatogr. Sci. 2013, 34, 254–257. [Google Scholar] [CrossRef] [Green Version]
- Rapp, A.; Mandery, H. Wine Aroma. Experientia 1986, 42, 873–884. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine Flavor and Aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past And Future Of Non-Saccharomyces Yeasts: From Spoilage Microorganisms To Biotechnological Tools For Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebeler, S.E.; Thorngate, J.H. Wine Chemistry and Flavor: Looking Into the Crystal Glass. J. Agric. Food Chem. 2009, 57, 8098–8108. [Google Scholar] [CrossRef] [PubMed]
- General Industry Stats. United States Wine and Grape Industry FAQS|Wineamerica. International Association of America Wineries. 2019. Available online: https://Wineamerica.Org/Policy/By-The-Numbers/ (accessed on 18 September 2020).
- Pennsylvania Winery Association. About Pennsylvania Wines—Pennsylvania Wines. 2020. Available online: Https://Pennsylvaniawine.Com/About/ (accessed on 18 September 2020).
- Raymond Eder, M.L.; Reynoso, C.; Lauret, S.C.; Rosa, A.L. Isolation and Identification of the Indigenous Yeast Population during Spontaneous Fermentation of Isabella (Vitis Labrusca L.) Grape Must. Front. Microbiol. 2017, 8, 532. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.R.; Dawson, V.T. The Bacteriostatic Action of Rose Bengal in Media Used For Plate Counts of Soil Fungi. Soil Sci. 1944, 58, 467–472. [Google Scholar] [CrossRef]
- Salfinger, Y.; Tortorello, M.L. Compendium of Methods for the Microbiological Examination of Foods; American Public Health Association: Washington, DC, USA, 2013. [Google Scholar]
- Henson, O.E. Dichloran as an Inhibitor of Mold Spreading In Fungal Plating Media: Effects on Colony Diameter and Enumeration. Appl. Environ. Microbiol. 1981, 42, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolski, P. Short protocols in molecular biology. A compendium of methods from “current protocols in molecular biology”. edited by Frederick, M. Ausubel et al., John Wiley & Sons, 1992., ISBN 0-471-57735-9. Acta Biotechnologica. 1993, 13, 88. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear Ribosomal Internal Transcribed Spacer (Its) Region As A Universal Dna Barcode Marker For Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Zabukovec, P.; Čadež, N.; Čuš, F. Isolation and Identification of Indigenous Wine Yeasts and Their Use in Alcoholic Fermentation. Food Technol. Biotechnol. 2020, 58, 337–347. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Mycological Society of America: Albuquerque, NM, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Brysch-Herzberg, M.; Seidel, M. Yeast Diversity on Grapes in Two German Wine Growing Regions. Int. J. Food Microbiol. 2015, 214, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. Mycobank: An Online Initiative to Launch Mycology into the 21st Century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (Mega) For Macos. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Buehler, A.J.; Evanowski, R.L.; Martin, N.H.; Boor, K.J.; Wiedmann, M. Internal Transcribed Spacer (Its) Sequencing Reveals Considerable Fungal Diversity in Dairy Products. J. Dairy Sci. 2017, 100, 8814–8825. [Google Scholar] [CrossRef]
- Tofalo, R.; Schirone, M.; Torriani, S.; Rantsiou, K.; Cocolin, L.; Perpetuini, G.; Suzzi, G. Diversity of Candida Zemplinina Strains From Grapes And Italian Wines. Food Microbiol. 2012, 29, 18–26. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Torchio, F.; Rolle, L.; Gerbi, V.; Cocolin, L. Exploitation of the Non-Saccharomyces Yeast Starmerella Bacillaris (Synonym Candida Zemplinina) In Wine Fermentation: Physiological and Molecular Characterizations. Int. J. Food Microbiol. 2015, 199, 33–40. [Google Scholar] [CrossRef]
- Capece, A.; Siesto, G.; Romaniello, R.; Lagreca, V.M.; Pietrafesa, R.; Calabretti, A.; Romano, P. Assessment Of Competition In Wine Fermentation Among Wild Saccharomyces Cerevisiae Strains Isolated From Sangiovese Grapes In Tuscany Region. LWT Food Sci. Technol. 2013, 54, 485–492. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Mestre, M.V.; Kuchen, B.; Toro, M.E.; Mercado, L.A.; Vazquez, F.; Combina, M. Optimization Of Fermentation-Relevant Factors: A Strategy To Reduce Ethanol In Red Wine By Sequential Culture Of Native Yeasts. Int. J. Food Microbiol. 2019, 289, 40–48. [Google Scholar] [CrossRef]
- Threlfall, R.; Main, G.; Morris, J. Effect of Freezing Grape Berries and Heating Must Samples on Extraction of Components and Composition Parameters of Red Wine Grape Varieties. Aust. J. Grape Wine Res. 2006, 12, 161–169. [Google Scholar] [CrossRef]
- Chidi, B.S.; Rossouw, D.; Buica, A.S.; Bauer, F.F. Determining The Impact Of Industrial Wine Yeast Strains On Organic Acid Production Under White And Red Wine-Like Fermentation Conditions. S. Afr. J. Enol. Vitic. 2015, 36, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Pedneault, K.; Dorais, M.; Angers, P. Flavor of Cold-Hardy Grapes: Impact of Berry Maturity and Environmental Conditions. J. Agric. Food Chem. 2013, 61, 10418–10438. [Google Scholar] [CrossRef] [PubMed]
- Fenoll, J.; Manso, A.; Hellín, P.; Ruiz, L.; Flores, P. Changes In The Aromatic Composition Of The Vitis Vinifera Grape Muscat Hamburg During Ripening. Food Chem. 2009, 114, 420–428. [Google Scholar] [CrossRef]
- Bromba, M.U.A.; Ziegler, H. Application Hints for Savitzky-Holay Digital Smoothing Filters. Anal. Chem. 1981, 53, 1583–1586. [Google Scholar] [CrossRef]
- Wenig, P.; Odermatt, J. Openchrom: A Cross-Platform Open Source Software for the Mass Spectrometric Analysis of Chromatographic Data; BMC Bioinformatics: Hamburg, Germany, 2010. [Google Scholar]
- Schafer, R.W. What Is A Savitzky-Golay Filter? IEEE Signal Process. Mag. 2011, 28, 111–117. [Google Scholar] [CrossRef]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas Chromatography—Mass Spectrometry Data Processing Made Easy. J. Chromatogr. A 2017, 1503, 57–64. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using Ibm Spss Statistics; SAGE Publications Ltd.: Thousand Oaks, CA, USA, 2013; pp. 1–816. [Google Scholar]
- Chong, J.; Wishart, D.S.; Xia, J. Using Metaboanalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Van Den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; Van Der Werf, M.J. Centering, Scaling, And Transformations: Improving The Biological Information Content Of Metabolomics Data. Bmc Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Manachini, P.L.; Parini, C.; Fortina, M.G. Pectic Enzymes from Aureobasidium Pullulans Lv 10. Enzyme Microb. Technol. 1988, 10, 682–685. [Google Scholar] [CrossRef]
- Wang, H.L. Characterization of Microbial Dynamics and Volatile Metabolome Changes during Fermentation of Chambourcin Grapes in Two Pennsylvania Regions. Master Thesis, The Pennsylvania State University, The Graduate School, State College, PA, USA, 2020. [Google Scholar]
- Liu, D.; Chen, Q.; Zhang, P.; Chen, D.; Howell, K.S. The Fungal Microbiome Is An Important Component Of Vineyard Ecosystems And Correlates With Regional Distinctiveness Of Wine. Msphere 2020, 5, e00534-20. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.M.H.; Boss, P.K.; Walker, M.E.; Sumby, K.M.; Grbin, P.R.; Jiranek, V. Evaluation Of Indigenous Non-Saccharomyces Yeasts Isolated From A South Australian Vineyard For Their Potential As Wine Starter Cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef] [PubMed]
- Baffi, M.A.; Dos Santos Bezerra, C.; Arévalo-Villena, M.; Isabel Briones-Pérez, A.; Gomes, E.; Da Silva, R. Isolation And Molecular Identification Of Wine Yeasts From A Brazilian Vineyard. Ann. Microbiol. 2011, 61, 75–78. [Google Scholar] [CrossRef]
- Bezerra-Bussoli, C.; Baffi, M.A.; Gomes, E.; Da-Silva, R. Yeast Diversity Isolated From Grape Musts during Spontaneous Fermentation from A Brazilian Winery. Curr. Microbiol. 2013, 67, 356–361. [Google Scholar] [CrossRef]
- Eder, M.L.R.; Conti, F.; Rosa, A.L. Differences between Indigenous Yeast Populations in Spontaneously Fermenting Musts from V. Vinifera L. And V. Labrusca L. Grapes Harvested In The Same Geographic Location. Front. Microbiol. 2018, 9, 1320. [Google Scholar] [CrossRef]
- Lopandic, K.; Tiefenbrunner, W.; Gangl, H.; Mandl, K.; Berger, S.; Leitner, G.; Abd-Ellah, G.A.; Querol, A.; Gardner, R.C.; Sterflinger, K.; et al. Molecular Profiling Of Yeasts Isolated During Spontaneous Fermentations Of Austrian Wines. FEMS Yeast Res. 2008, 8, 1063–1075. [Google Scholar] [CrossRef] [Green Version]
- Pina, C.; Santos, C.; Couto, J.A.; Hogg, T. Ethanol Tolerance of Five Non-Saccharomyces Wine Yeasts In Comparison With A Strain of Saccharomyces Cerevisiae—Influence Of Different Culture Conditions. Food Microbiol. 2004, 21, 439–447. [Google Scholar] [CrossRef]
- Kim, I.S.; Kim, Y.S.; Kim, H.; Jin, I.; Yoon, H.S. Saccharomyces Cerevisiae Knu5377 Stress Response During High-Temperature Ethanol Fermentation. Mol. Cells 2013, 35, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xia, Y.; Lin, X.; Wang, G.; Zhang, H.; Xiong, Z.; Yu, H.; Yu, J.; Ai, L. Improvement Of Flavor Profiles In Chinese Rice Wine By Creating Fermenting Yeast With Superior Ethanol Tolerance And Fermentation Activity. Food Res. Int. 2018, 108, 83–92. [Google Scholar] [CrossRef]
- Gao, C.; Fleet, G.H. Degradation Of Malic And Tartaric Acids By High Density Cell Suspensions Of Wine Yeasts. Food Microbiol. 1995, 12, 65–71. [Google Scholar] [CrossRef]
- Hong, S.K.; Lee, H.J.; Park, H.J.; Hong, Y.A.; Rhee, I.K.; Lee, W.H.; Choi, S.W.; Lee, O.S.; Park, H.D. Degradation Of Malic Acid In Wine By Immobilized Issatchenkia Orientalis Cells With Oriental Oak Charcoal And Alginate. Lett. Appl. Microbiol. 2010, 50, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic Acid: Technology Development and Commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Qian, M.C.; Wang, Y. Seasonal Variation of Volatile Composition and Odor Activity Value of “Marion” (Rubus spp. hyb) and “Thornless Evergreen” (R. laciniatus L.) Blackberries. J. Food Sci. 2005, 70, C13–C20. [Google Scholar] [CrossRef]
- Plotto, A.; Barnes, K.W.; Goodner, K.L. Specific Anosmia Observed For β-Ionone, But Not For α-Ionone: Significance for Flavor Research. J. Food Sci. 2006, 71, S401–S406. [Google Scholar] [CrossRef]
- Tripathi, J.; Chatterjee, S.; Gamre, S.; Chattopadhyay, S.; Variyar, P.S.; Sharma, A. Analysis Of Free And Bound Aroma Compounds Of Pomegranate (Punica Granatum L.). LWT Food Sci. Technol. 2014, 59, 461–466. [Google Scholar] [CrossRef]
- Deed, R.C.; Hou, R.; Kinzurik, M.I.; Gardner, R.C.; Fedrizzi, B. The Role Of Yeast Aro8, Aro9 And Aro10 Genes In The Biosynthesis Of 3-(Methylthio)-1-Propanol From L-Methionine During Fermentation In Synthetic Grape Medium. FEMS Yeast Res. 2019, 19, 109. [Google Scholar] [CrossRef]
- Schreier, P.; Paroschy, J.H. Volatile Constituents from Concord, Niagara (Vitis Labrusca, L.) And Elvira (V. Labrusca, L. × V. Riparia, M.) Grapes. Can. Inst. Food Sci. Technol. J. 1981, 14, 112–118. [Google Scholar] [CrossRef]
- Zakarya, D.; Yahiaoui, M.; Fkih-Tetouani, S. Structure–Odour Relations for Bitter Almond Odorants. J. Phys. Org. Chem. 1993, 6, 627–633. [Google Scholar] [CrossRef]
- Masino, F.; Montevecchi, G.; Riponi, C.; Antonelli, A. Composition Of Some Commercial Grappas (Grape Marc Spirit): The Anomalous Presence Of 1,1-Diethoxy-3-Methylbutane: A Case Study. Eur. Food Res. Technol. 2009, 228, 565–569. [Google Scholar] [CrossRef]
- Niu, Y.; Yao, Z.; Xiao, Z.; Zhu, G.; Zhu, J.; Chen, J. Sensory Evaluation Of The Synergism Among Ester Odorants In Light Aroma-Type Liquor By Odor Threshold, Aroma Intensity And Flash Gc Electronic Nose. Food Res. Int. 2018, 113, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Henschke, P. Stuck Fermentation: Causes, Prevention And Cure. In Advances in Juice Clarification and Yeast Inoculation; Australian Society of Viticulture and Oenology: Adelaide, Australia, 1997; pp. 30–41. [Google Scholar]
- Sadineni, V.; Kondapalli, N.; Reddy Obulam, V.S. Effect Of Co-Fermentation With Saccharomyces Cerevisiae And Torulaspora Delbrueckii Or Metschnikowia Pulcherrima On The Aroma And Sensory Properties Of Mango Wine. Ann. Microbiol. 2012, 62, 1353–1360. [Google Scholar] [CrossRef]
- Maisonnave, P.; Sanchez, I.; Moine, V.; Dequin, S.; Galeote, V. Stuck Fermentation: Development of a Synthetic Stuck Wine and Study of a Restart Procedure. Int. J. Food Microbiol. 2013, 163, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Querol, A.; Pérez-Torrado, R.; Alonso-Del-Real, J.; Minebois, R.; Stribny, J.; Oliveira, B.M.; Barrio, E. New Trends in the Uses of Yeasts in Oenology. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2018; Volume 85, pp. 177–210. ISBN 9780128150894. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, C.T.; Du, X.; Wee, J. Microbial and Chemical Analysis of Non-Saccharomyces Yeasts from Chambourcin Hybrid Grapes for Potential Use in Winemaking. Fermentation 2021, 7, 15. https://doi.org/10.3390/fermentation7010015
Feng CT, Du X, Wee J. Microbial and Chemical Analysis of Non-Saccharomyces Yeasts from Chambourcin Hybrid Grapes for Potential Use in Winemaking. Fermentation. 2021; 7(1):15. https://doi.org/10.3390/fermentation7010015
Chicago/Turabian StyleFeng, Chun Tang, Xue Du, and Josephine Wee. 2021. "Microbial and Chemical Analysis of Non-Saccharomyces Yeasts from Chambourcin Hybrid Grapes for Potential Use in Winemaking" Fermentation 7, no. 1: 15. https://doi.org/10.3390/fermentation7010015
APA StyleFeng, C. T., Du, X., & Wee, J. (2021). Microbial and Chemical Analysis of Non-Saccharomyces Yeasts from Chambourcin Hybrid Grapes for Potential Use in Winemaking. Fermentation, 7(1), 15. https://doi.org/10.3390/fermentation7010015