
Development of Korean Indigenous Low-Temperature-Tolerant Yeast for Enhancing the Flavor of Yakju


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Screening of Yakju Yeast
2.2.1. Low-Temperature Fermentation Activity
2.2.2. Screening by Internal Transcribed Spacer (ITS) Region Sequencing
2.2.3. β-Glucosidase Enzyme Activity
2.2.4. Biosynthesis Inhibitor Resistance Activity
2.3. Analysis of Low-Temperature-Fermented Yakju
2.3.1. Brewing Yakju
2.3.2. Fermentation Characteristics of Yakju
2.3.3. Volatile Aromatic Compound Analysis by Gas Chromatography
2.4. Statistical Analysis
3. Results and Discussion
3.1. Screening of Yakju Strains
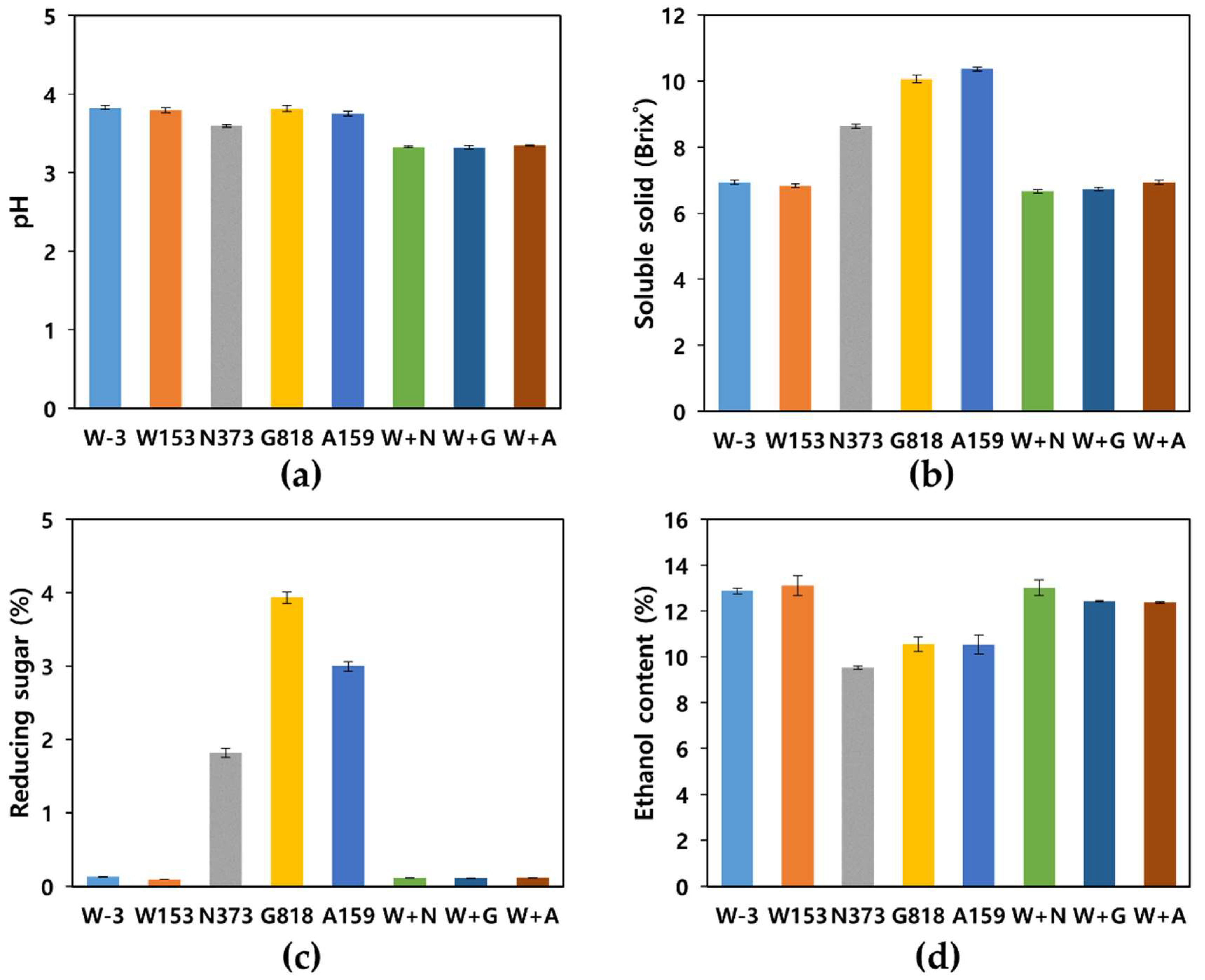
3.2. Characteristics of Yakju
3.3. Volatile Aromatic Compounds
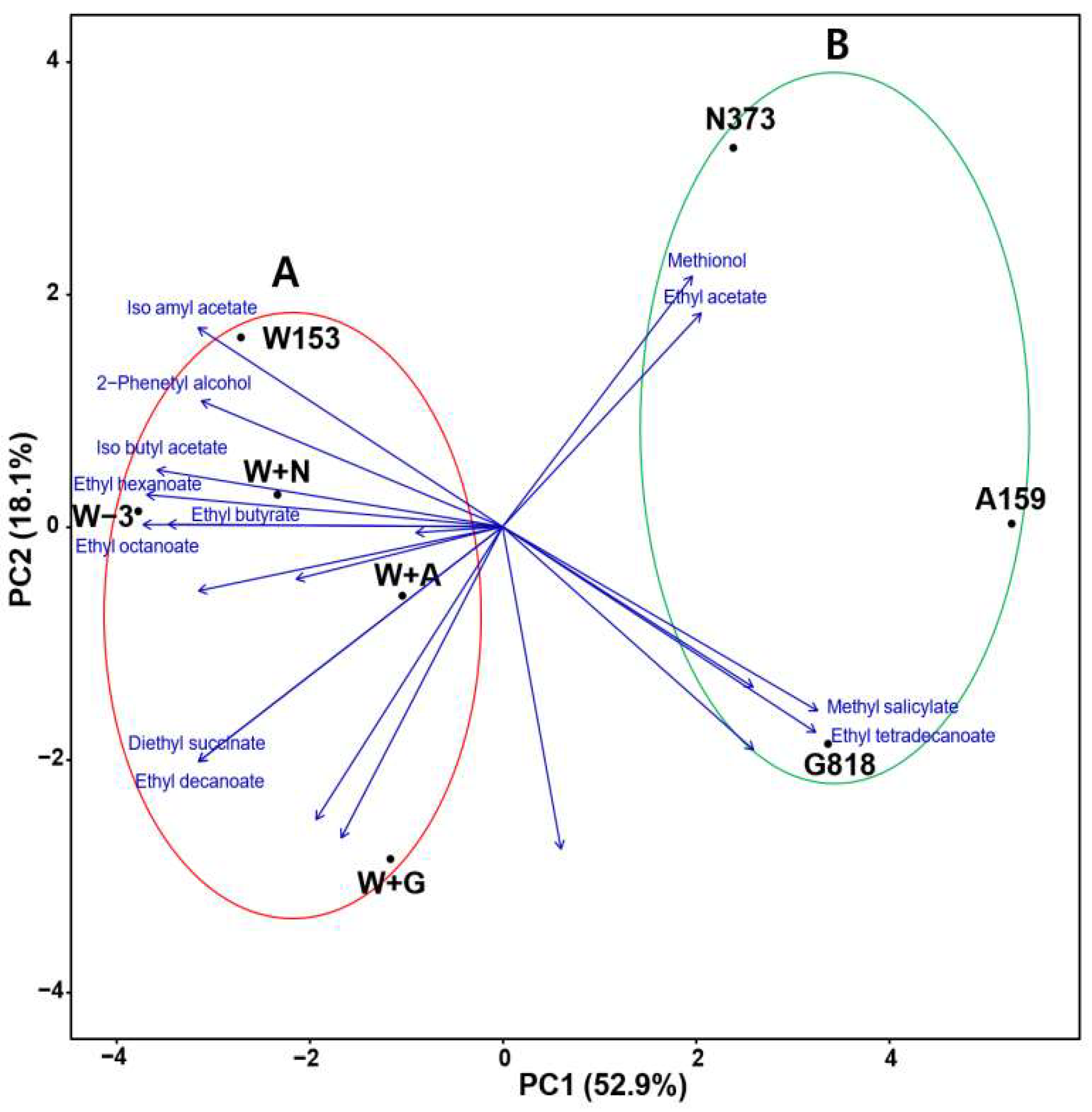
3.4. Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, K.; Tuszyński, T. The effect of temperature on fermentation and beer volatiles at an industrial scale. J. Inst. Brew. 2018, 124, 230–235. [Google Scholar] [CrossRef]
- Torija, M.J.; Rozes, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Effects of fermentation temperature on the strain population of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 80, 47–53. [Google Scholar] [CrossRef]
- Gamero, A.; Tronchoni, J.; Querol, A.; Belloch, C. Production of aroma compounds by cryotolerant Saccharomyces species and hybrids at low and moderate fermentation temperatures. J. Appl. Microbiol. 2013, 114, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.; Randez-Gil, F.; Prieto, J.A. Cold response in Saccharomyces cerevisiae: New functions for old mechanisms. FEMS Microbiol. Rev. 2007, 31, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Schade, B.; Jansen, G.; Whiteway, M.; Entian, K.D.; Thomas, D.Y. Cold adaptation in budding yeast. Mol. Biol. Cell 2004, 15, 5492–5502. [Google Scholar] [CrossRef] [Green Version]
- Seo, D.-J.; Yeo, S.-H.; Mun, J.-Y.; Baek, S.Y. Effects of low temperature-adapted Saccharomyces cerevisiae Y297 strain and fermentation temperature on the quality characteristics of Yakju. Korean J. Food Preserv. 2016, 23, 666–672. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-R.; Kim, J.-H.; Bae, D.-H.; Ahn, B.-H. Characterization of Yakju brewed from glutinous rice and wild-type yeast strains isolated from nuruks. J. Microbiol. Biotechnol. 2010, 20, 1702–1710. [Google Scholar] [PubMed]
- Kim, H.-R.; Kwon, Y.-H.; Jo, S.-J.; Kim, J.-H.; Ahn, B.-H. Characterization and volatile flavor components in glutinous rice wines prepared with different yeasts of nuruks. Korean J. Food Sci. Technol. 2009, 41, 296–301. [Google Scholar]
- Rhee, S.J.; Lee, C.-Y.J.; Kim, K.K.; Lee, C.-H. Comparison of the traditional (Samhaeju) and industrial (Chongju) rice wine brewing in Korea. Food Sci. Biotechnol. 2003, 12, 242–247. [Google Scholar]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of Hanseniaspora uvarum in simultaneous and sequential co-fermentation with Saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Escott, C.; Loira, I.; Del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Larroque, M.; Carrau, F.; Fariña, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Fusco, V.; Böhnlein, C.; Kabisch, J.; Logrieco, A.F.; Habermann, D.; Cho, G.-S.; Benomar, N.; Abriouel, H.; Schmidt-Heydt, M. The life and times of yeasts in traditional food fermentations. Crit. Rev. Food Sci. Nutr. 2020, 60, 3103–3132. [Google Scholar] [CrossRef]
- Loira, I.; Vejarano, R.; Bañuelos, M.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suárez-Lepe, J. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT-Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Bellut, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Application of non-Saccharomyces yeasts isolated from kombucha in the production of alcohol-free beer. Fermentation 2018, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Chasseriaud, L.; Coulon, J.; Marullo, P.; Albertin, W.; Bely, M. New oenological practice to promote non-Saccharomyces species of interest: Saturating grape juice with carbon dioxide. Appl. Microbiol. Biotechnol. 2018, 102, 3779–3791. [Google Scholar] [CrossRef]
- Lleixà, J.; Martín, V.; Portillo, M.d.C.; Carrau, F.; Beltran, G.; Mas, A. Comparison of fermentation and wines produced by inoculation of Hanseniaspora vineae and Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 338. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Jin, G.-J.; Xu, Y.-H.; Tao, Y.-S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of montepulciano d’abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Ciani, M.; Fatichenti, F. Killer toxin of Kluyveromyces phaffii DBVPG 6076 as a biopreservative agent to control apiculate wine yeasts. Appl. Environ. Microbiol. 2001, 67, 3058–3063. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Ingeniis De, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Lõoke, M.; Kristjuhan, K.; Kristjuhan, A. Extraction of genomic DNA from yeasts for PCR-based applications. Biotechniques 2011, 50, 325–328. [Google Scholar] [CrossRef]
- Rodríguez, M.; Lopes, C.; Van Broock, M.; Valles, S.; Ramon, D.; Caballero, A. Screening and typing of Patagonian wine yeasts for glycosidase activities. J. Appl. Microbiol. 2004, 96, 84–95. [Google Scholar] [CrossRef]
- Akita, O.; Ida, T.; Obata, T.; Hara, S. Mutants of Saccharomyces cerevisiae producing a large quantity of β-phenethyl alcohol and β-phenethyl acetate. J. Ferment. Bioeng. 1990, 69, 125–128. [Google Scholar] [CrossRef]
- Lee, S.B.; Banda, C.; Park, H.D. Effect of inoculation strategy of non-Saccharomyces yeasts on fermentation characteristics and volatile higher alcohols and esters in Campbell Early wines. Aust. J. Grape Wine Res. 2019, 25, 384–395. [Google Scholar] [CrossRef]
- Lee, S.-B.; Park, H.-D. Isolation and investigation of potential non-Saccharomyces yeasts to improve the volatile terpene compounds in Korean Muscat Bailey A wine. Microorganisms 2020, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Sarry, J.-E.; Günata, Z. Plant and microbial glycoside hydrolases: Volatile release from glycosidic aroma precursors. Food Chem. 2004, 87, 509–521. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, X.L.; Ullah, N.; Tao, Y.S. Aroma glycosides in grapes and wine. J. food Sci. 2017, 82, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.J.; Yeo, S.H.; Park, J.-H.; Choi, H.S.; Gang, J.-E.; Kim, S.I.; Jeong, S.T.; Kim, S.R. Isolation of aromatic yeasts (non-Saccharomyces cerevisiae) from Korean traditional nuruks and identification of fermentation characteristics. Agric. Sci. 2013, 4, 136. [Google Scholar]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional yeast species for lowering ethanol content of wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, A.; Hidalgo, C.; Henschke, P.; Chambers, P.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.R.; Kim, J.-H.; Bai, D.-H.; Ahn, B. Feasibility of brewing Makgeolli using Pichia anomala Y197-13, a non-Saccharomyces cerevisiae. J. Microbiol. Biotechnol. 2012, 22, 1749–1757. [Google Scholar] [CrossRef] [Green Version]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.-J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast–yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Zhang, B.-Q.; Shen, J.-Y.; Duan, C.-Q.; Yan, G.-L. Use of indigenous Hanseniaspora vineae and Metschnikowia pulcherrima co-fermentation with Saccharomyces cerevisiae to improve the aroma diversity of Vidal Blanc icewine. Front. Microbiol. 2018, 9, 2303. [Google Scholar] [CrossRef]
- Heo, J.; Kwak, H.S.; Kim, M.; Kim, J.-H.; Baek, H.H.; Shin, H.; Lee, Y.-s.; Lee, S.; Kim, S.S. Major sensory attributes and volatile compounds of Korean rice liquor (yakju) affecting overall acceptance by young consumers. Foods 2020, 9, 722. [Google Scholar] [CrossRef]
- Rapp, A.; Mandery, H. Wine aroma. Experientia 1986, 42, 873–884. [Google Scholar] [CrossRef]
- Carpena, M.; Fraga-Corral, M.; Otero, P.; Nogueira, R.A.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Secondary aroma: Influence of wine microorganisms in their aroma profile. Foods 2021, 10, 51. [Google Scholar] [CrossRef]
- Ehsani, M.; Fernández, M.R.; Biosca, J.A.; Julien, A.; Dequin, S. Engineering of 2, 3-butanediol dehydrogenase to reduce acetoin formation by glycerol-overproducing, low-alcohol Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 3196–3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, P.; Palla, G.; Caligiani, A.; Brandolini, V.; Maietti, A.; Salzano, G. Evaluation of stereoisomers of 2, 3-butanediol and acetoin to differentiate Saccharomyces cerevisiae and Kloeckera apiculata wine strains. Biotechnol. Lett. 2000, 22, 1947–1951. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Boscolo, M.; Bezerra, C.W.; Cardoso, D.R.; Lima Neto, B.S.; Franco, D.W. Identification and dosage by HRGC of minor alcohols and esters in Brazilian sugar-cane spirit. J. Braz. Chem. Soc. 2000, 11, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Sun, B.; Li, X.; Fan, G.; Teng, C.; Alaa, A.; Jia, Y. Isolation and characterization of a high ethyl acetate-producing yeast from Laobaigan Daqu and its fermentation conditions for producing high-quality Baijiu. Biotechnol. Biotechnol. Equip. 2018, 32, 1218–1227. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Jin, G.-J.; Mei, W.-C.; Li, T.; Tao, Y.-S. Increase of medium-chain fatty acid ethyl ester content in mixed H. uvarum/S. cerevisiae fermentation leads to wine fruity aroma enhancement. Food Chem. 2018, 239, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tsuchiya, F.; Isogai, A. Relationship between medium-chain fatty acid contents and organoleptic properties of Japanese sake. J. Agric. Food Chem. 2014, 62, 8478–8485. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, A.V.; Muñoz, R.; González, R. Molecular Wine Microbiology; Elsevier: Cham, Switzerland, 2011. [Google Scholar]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Heinz, H.A.; Elkins, J.T. Comparison of unaged and barrel aged whiskies from the same Mash Bill using gas chromatography/mass spectrometry. J. Brew. Distill. 2019, 8, 1–6. [Google Scholar]



{kind=link}
{kind=link}
| Strains | Species | Cell Growth (OD600nm, 15 °C) | β-Glucosidase Activity (U/mL) | Cerulenin Resistance | FPA Resistance |
|---|---|---|---|---|---|
| W153 | Saccharomyces cerevisiae | 1.615 | 0.679 | + | − |
| N373 | Pichia kudriavzevii | 1.285 | 1.408 | + | + |
| G818 | Hanseniaspora vineae | 0.977 | 0.567 | − | + |
| A159 | Wickerhamomyces anomalus | 1.161 | 0.922 | + | − |
| Volatile Compounds | W-3 | W153 | N373 | G818 | A159 | W153 + N373 | W153 + G818 | W153 + A159 |
|---|---|---|---|---|---|---|---|---|
| Higher alcohols (mg/L) | ||||||||
| 1-Propanol | 30.40 ± 0.33 b(1)(2) | 6.82 ± 0.19 g | 10.01 ± 0.62 f | 21.41 ± 0.31 d | 6.78 ± 0.45 g | 28.38 ± 1.13 c | 48.46 ± 0.86 a | 17.12 ± 0.36 e |
| Iso-butanol | 241.22 ± 10.90 a | 144.52 ± 4.81 c | 83.94 ± 0.64 e | 39.25 ± 2.84 f | 25.09 ± 0.98 g | 181.01 ± 7.40 b | 146.87 ± 6.87 c | 114.08 ± 3.78 d |
| Isoamyl alcohol | 158.89 ± 8.89 d | 245.08 ± 5.25 b | 200.81 ± 8.18 c | 196.14 ± 9.31 c | 110.76 ± 6.46 e | 204.56 ± 5.63 c | 262.37 ± 6.86 a | 48.93 ± 5.12 f |
| Methionol | 2.89 ± 0.77 d | 4.19 ± 0.29 c | 10.92 ± 0.39 a | 3.03 ± 0.52 d | 8.26 ± 0.86 b | 4.69 ± 0.93 c | 4.43 ± 0.27 c | 7.67 ± 0.69 b |
| 2-Phenetyl alcohol | 85.44 ± 8.63 cd | 95.92 ± 6.22 ab | 76.80 ± 4.08 d | 27.21 ± 2.18 f | 42.16 ± 1.60 e | 87.86 ± 6.16 bc | 80.54 ± 3.58 cd | 104.26 ± 6.24 a |
| 2,3-Butanediol | 36.75 ± 1.79 a | 11.35 ± 0.95 ef | 14.34 ± 1.63 d | 10.13 ± 0.62 f | 12.58 ± 0.28 de | 13.55 ± 0.77 d | 18.95 ± 0.96 c | 25.72 ± 1.59 b |
| Esters (mg/L) | ||||||||
| Ethyl acetate | 26.08 ± 2.69 e | 28.29 ± 3.19 ef | 237.70 ± 28.8 a | 115.95 ± 7.31 b | 71.74 ± 8.14 cd | 85.72 ± 7.17 c | 56.62 ± 5.13 d | 61.76 ± 3.74 d |
| Isobutyl acetate | 8.41 ± 0.67 a | 9.17 ± 0.77 a | 2.88 ± 0.35 c | 1.06 ± 0.16 d | 2.06 ± 0.30 cd | 9.07 ± 1.32 a | 5.45 ± 0.54 b | 7.97 ± 1.22 a |
| Ethyl butyrate | 0.42 ± 0.03 a | 0.40 ± 0.02 ab | 0.17 ± 0.02 d | 0.09 ± 0.01 e | 0.21 ± 0.02 d | 0.42 ± 0.02 a | 0.36 ± 0.01 bc | 0.33 ± 0.06 c |
| Isoamyl acetate | 10.62 ± 1.56 a | 9.74 ± 0.43 ab | 7.20 ± 2.03 c | 4.04 ± 0.36 d | 0.85 ± 0.17 e | 8.12 ± 1.27 bc | 4.60 ± 0.57 d | 5.02 ± 0.16 d |
| Ethyl hexanoate | 0.50 ± 0.02 b | 0.54 ± 0.01 a | 0.10 ± 0.03 e | 0.07 ± 0.01 ef | 0.05 ± 0.01 f | 0.49 ± 0.02 b | 0.30 ± 0.01 d | 0.35 ± 0.03 c |
| Ethyl octanoate | 1.36 ± 0.22 a | 1.39 ± 0.08 a | 0.16 ± 0.13 d | 0.17 ± 0.04 d | 0.06 ± 0.02 d | 1.27 ± 0.11 a | 0.81 ± 0.12 c | 1.05 ± 0.09 b |
| Ethyl nonanoate | 5.84 ± 0.32 e | 3.81 ± 0.33 f | 7.17 ± 0.41 e | 11.54 ± 1.12 c | 15.88 ± 0.35 a | 8.75 ± 1.12 d | 10.60 ± 1.36 c | 12.98 ± 0.81 b |
| Ethyl decanoate | 26.05 ± 1.02 ab | 22.73 ± 1.31 c | 4.04 ± 0.47 e | 19.37 ± 1.39 d | 1.80 ± 0.08 f | 23.53 ± 1.35 c | 28.08 ± 1.98 a | 23.99 ± 1.19 bc |
| Diethyl succinate | 2.59 ± 0.36 ab | 2.26 ± 0.15 bc | 0.40 ± 0.06 d | 1.93 ± 0.27 c | 0.18 ± 0.05 d | 2.34 ± 0.24 b | 2.80 ± 0.09 a | 2.39 ± 0.08 b |
| Ethyl dodecanoate | 2.63 ± 0.15 d | 1.74 ± 0.09 ef | 1.60 ± 0.11 f | 2.08 ± 0.09 e | 1.49 ± 0.08 f | 3.17 ± 0.18 c | 4.41 ± 0.27 a | 3.74 ± 0.48 b |
| Methyl salicylate | 1.43 ± 0.18 c | 1.48 ± 0.07 c | 1.65 ± 0.16 bc | 2.08 ± 0.17 a | 2.05 ± 0.24 a | 1.40 ± 0.16 c | 1.71 ± 0.19 bc | 1.83 ± 0.15 ab |
| 2-Phenethyl acetate | 3.18 ± 0.28 de | 5.77 ± 0.35 c | 1.59 ± 0.26 ef | 33.76 ± 1.80 a | 0.26 ± 0.02 f | 4.08 ± 0.94 cd | 28.38 ± 1.97 b | 0.75 ± 0.09 f |
| Ethyl tetradecanoate | 2.66 ± 0.13 fg | 2.38 ± 0.33 g | 3.39 ± 0.27 e | 8.44 ± 0.45 b | 10.51 ± 0.51 a | 3.07 ± 0.28 ef | 5.61 ± 0.07 c | 4.66 ± 0.38 d |
| Ethyl hexadecanoate | 10.39 ± 1.26 g | 14.78 ± 0.96 f | 17.78 ± 2.47 f | 38.00 ± 2.33 d | 95.88 ± 1.07 a | 29.40 ± 2.39 e | 43.76 ± 1.48 c | 50.65 ± 1.93 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-B.; Choi, J.-S.; Park, H.-W.; Lee, S.-B.; Park, H.-D. Development of Korean Indigenous Low-Temperature-Tolerant Yeast for Enhancing the Flavor of Yakju. Fermentation 2021, 7, 260. https://doi.org/10.3390/fermentation7040260
Park J-B, Choi J-S, Park H-W, Lee S-B, Park H-D. Development of Korean Indigenous Low-Temperature-Tolerant Yeast for Enhancing the Flavor of Yakju. Fermentation. 2021; 7(4):260. https://doi.org/10.3390/fermentation7040260
Chicago/Turabian StylePark, Jong-Beom, Jun-Su Choi, Hye-Won Park, Sae-Byuk Lee, and Heui-Dong Park. 2021. "Development of Korean Indigenous Low-Temperature-Tolerant Yeast for Enhancing the Flavor of Yakju" Fermentation 7, no. 4: 260. https://doi.org/10.3390/fermentation7040260
APA StylePark, J.-B., Choi, J.-S., Park, H.-W., Lee, S.-B., & Park, H.-D. (2021). Development of Korean Indigenous Low-Temperature-Tolerant Yeast for Enhancing the Flavor of Yakju. Fermentation, 7(4), 260. https://doi.org/10.3390/fermentation7040260