
Correlations between the Chemical, Microbiological Characteristics and Sensory Profile of Fungal Fermented Food

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Inoculums
2.2. Fungal Fermentation
2.3. Analytical Methods
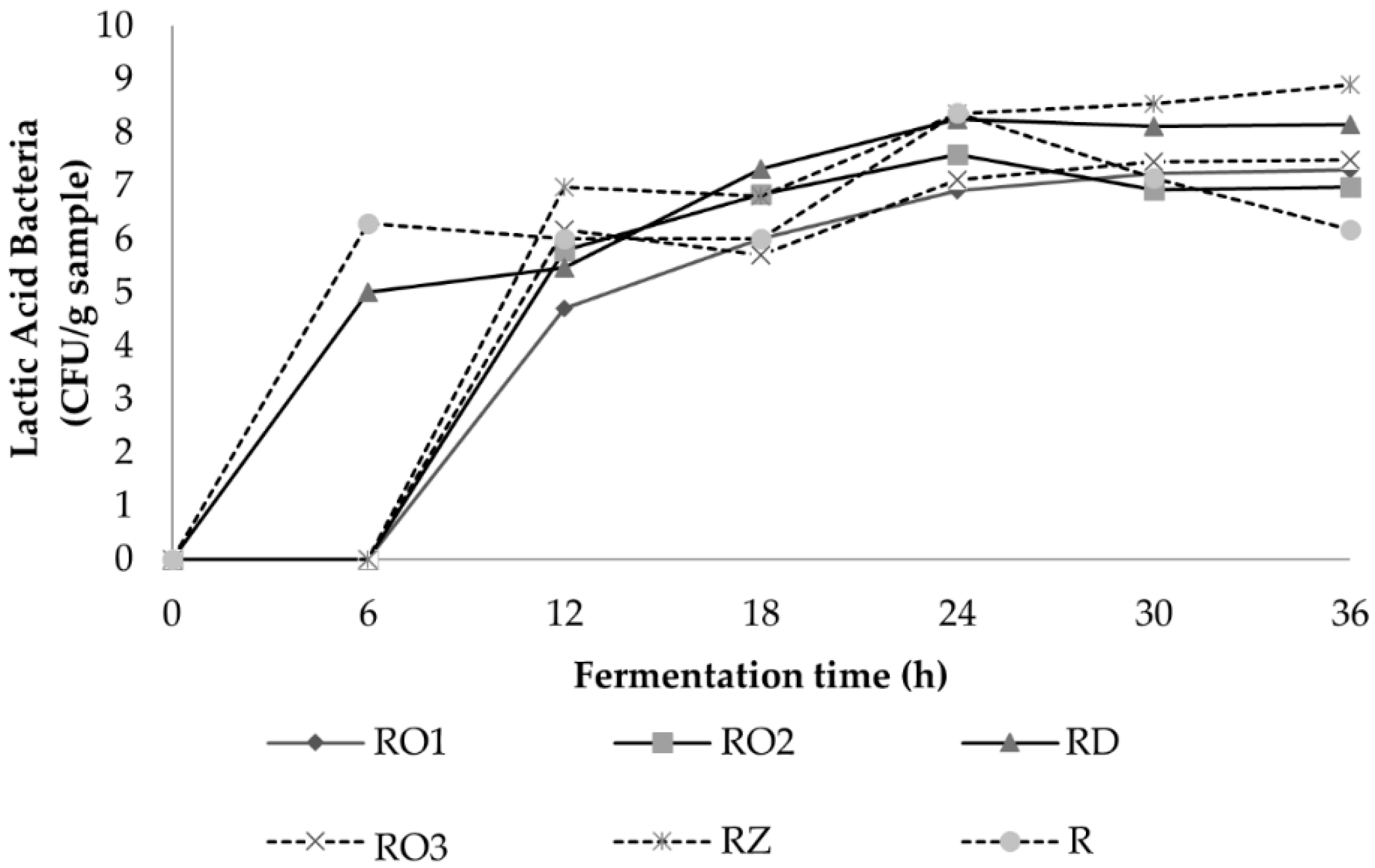
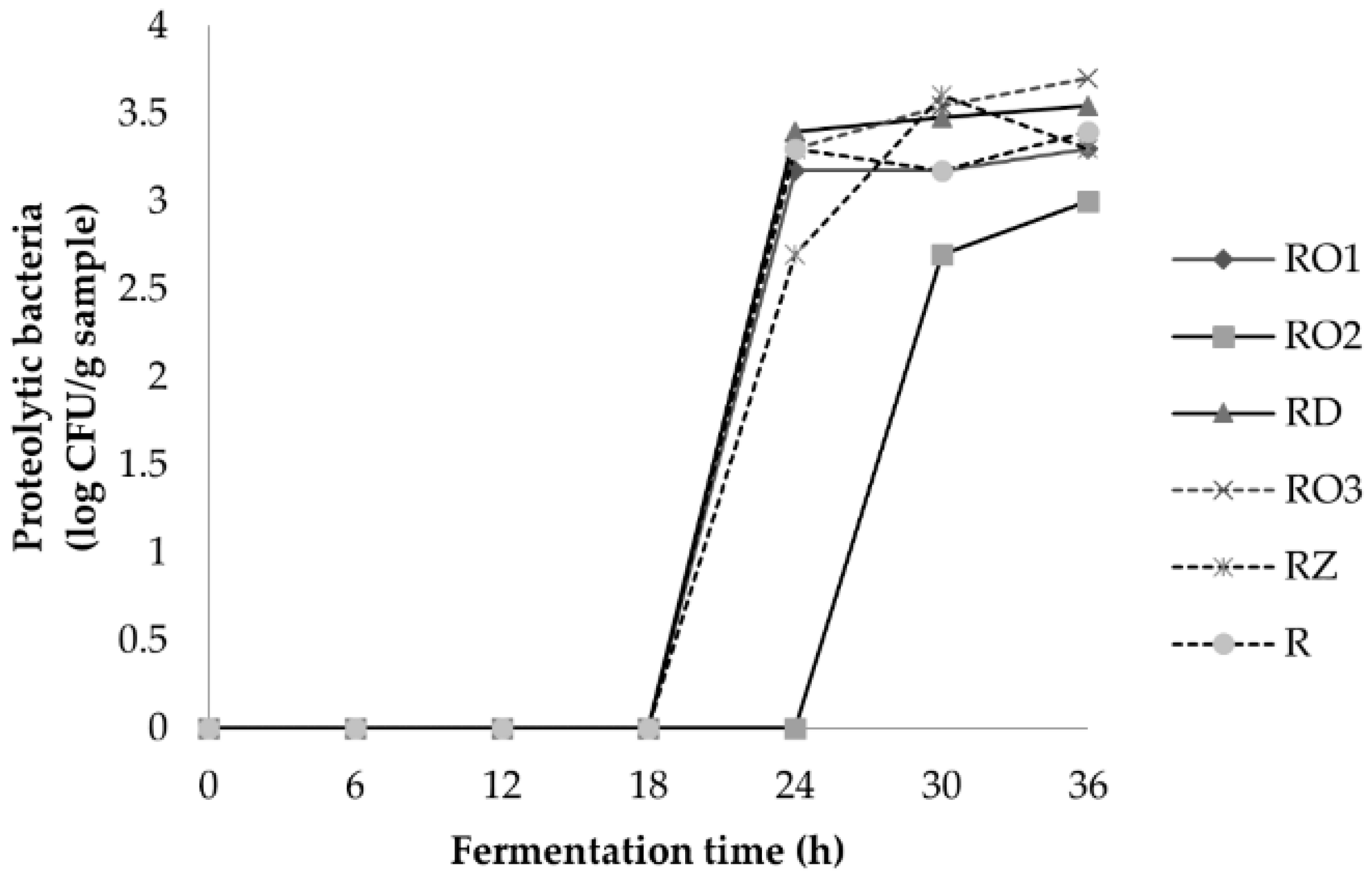
2.3.1. Microflora Analysis
2.3.2. Amino Acid Composition
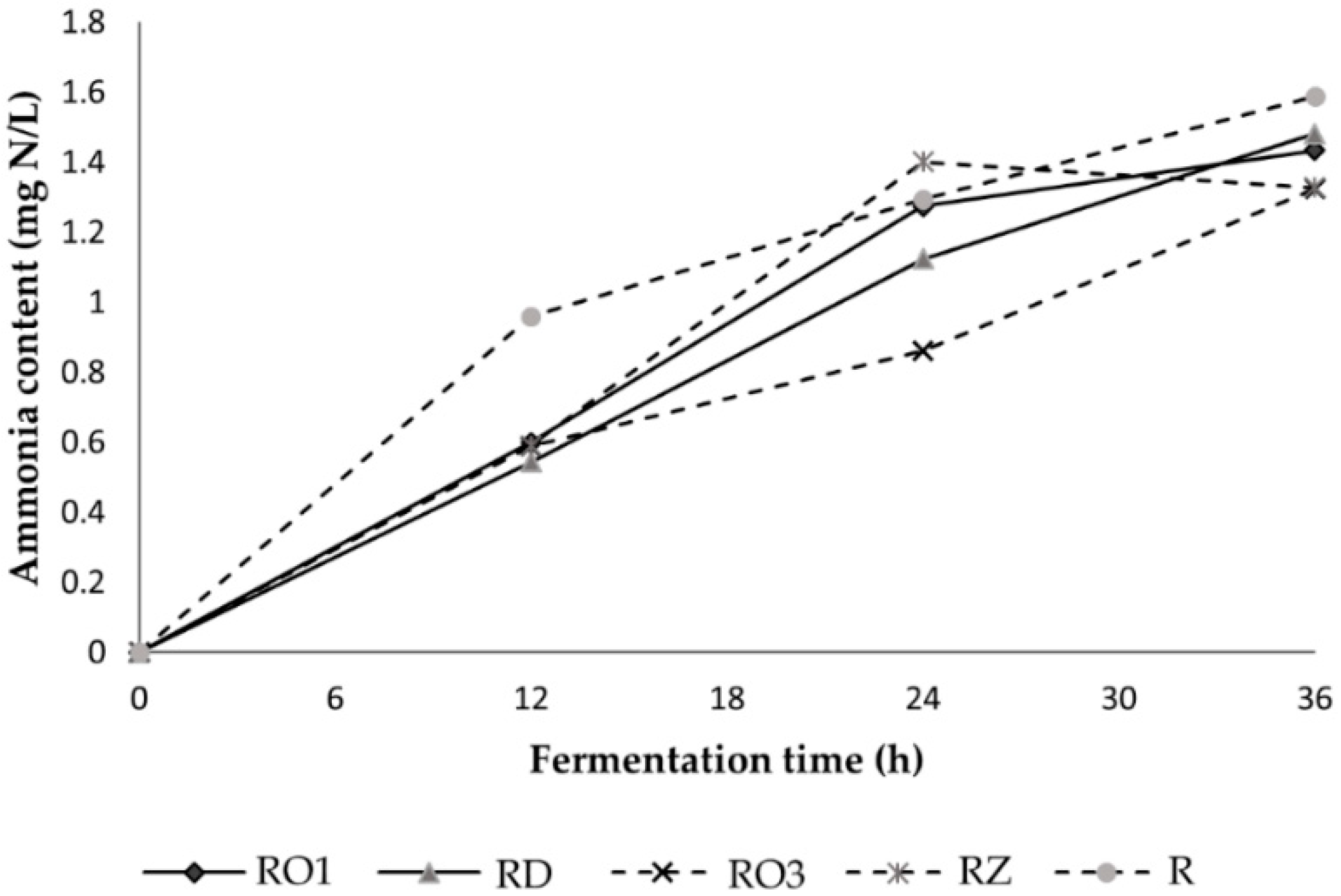
2.3.3. Analysis of Ammonia
2.4. Sensory Evaluation
2.5. Statistical Analyses
3. Results and Discussions
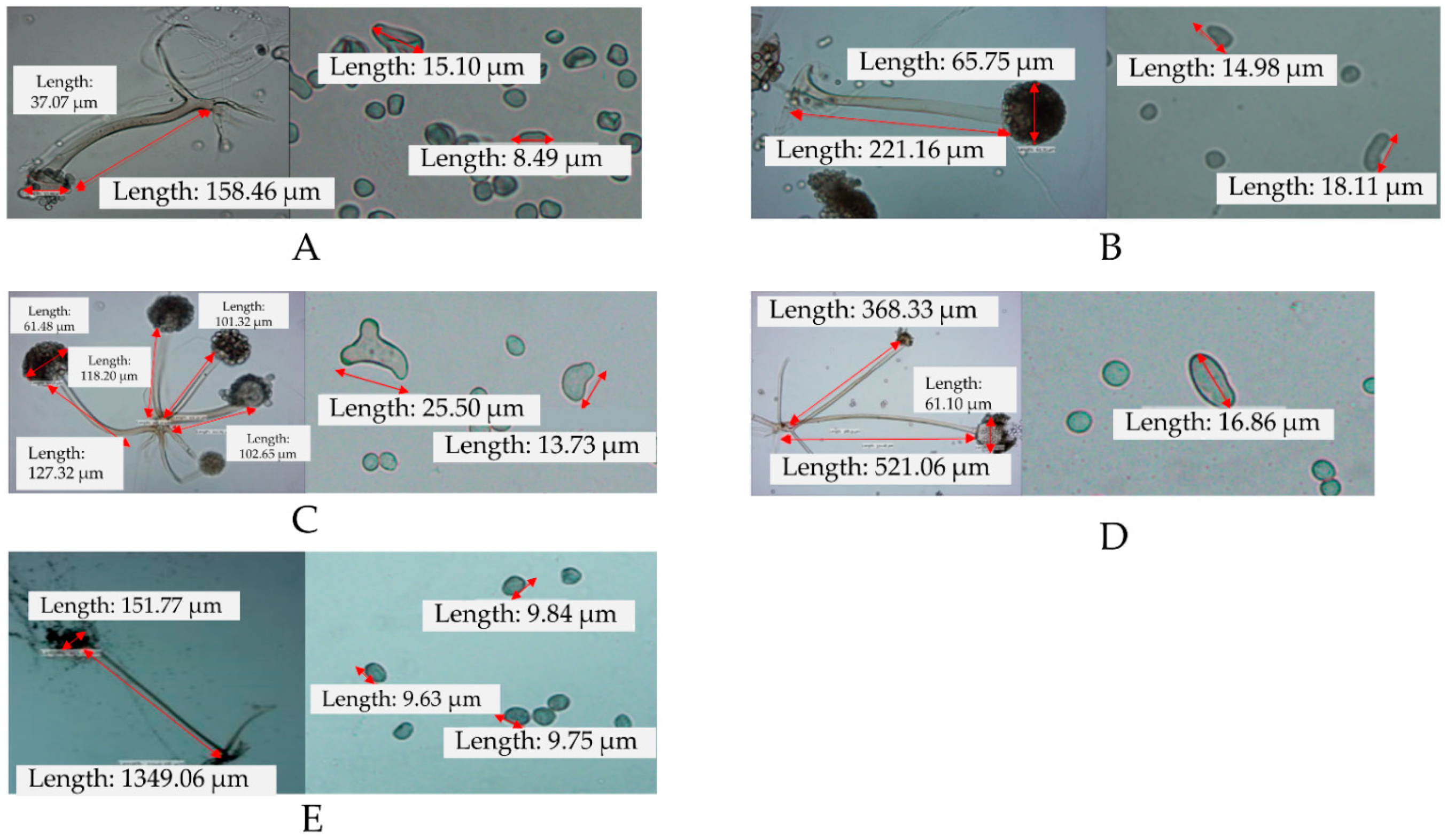
3.1. Isolation and Identification of the Rhizopus sp.
3.2. Fungal Fermentation
3.2.1. Chemical Profiles of Fungal-Fermented Soybean Products
3.2.2. Microflora Profile during Fermentation
3.3. Sensory Evaluation of Tempe Made from Different Fungal Strains
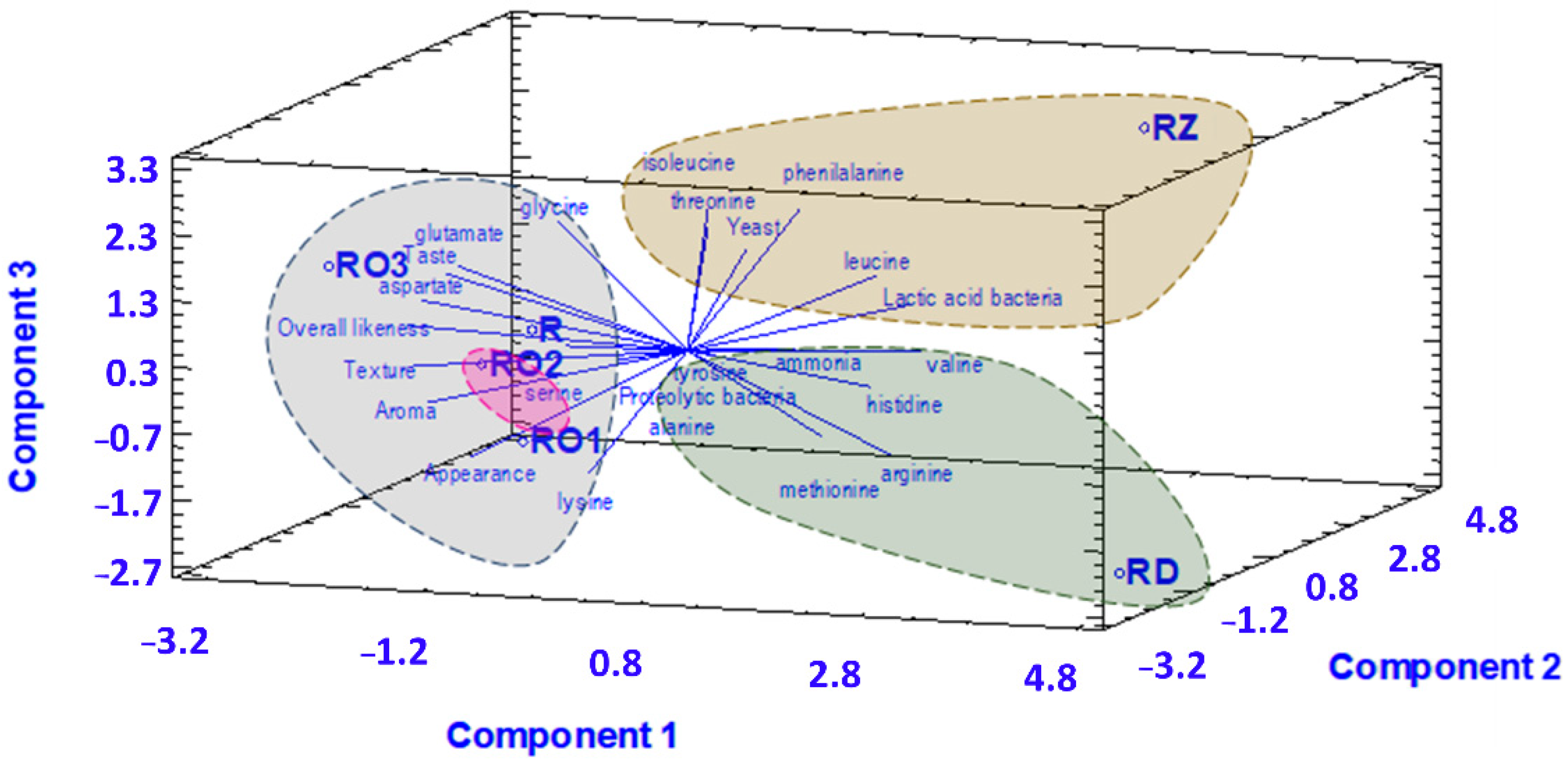
3.4. Correlation between Chemical and Microbioligal Characteristic with the Sensory Characteristics of Tempe
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castanheira, É.G.; Freire, F. Greenhouse Gas Assessment of Soybean Production: Implications of Land Use Change and Different Cultivation Systems. J. Clean. Prod. 2013, 54, 49–60. [Google Scholar] [CrossRef]
- Wiedemann, S.; McGahan, E.; Murphy, C.; Yan, M.-J.; Henry, B.; Thoma, G.; Ledgard, S. Environmental Impacts and Resource Use of Australian Beef and Lamb Exported to the USA Determined Using Life Cycle Assessment. J. Clean. Prod. 2015, 94, 67–75. [Google Scholar] [CrossRef]
- Liu, K. Chemistry and Nutritional Value of Soybean Components. In Soybeans: Chemistry, Technology, and Utilization; Springer: Boston, MA, USA, 1997. [Google Scholar]
- Hassan, S. Soybean, Nutrition and Health. In Soybean—Bio-Active Compounds; El-Shemy, H.A., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2013. [Google Scholar]
- Adeyemo, S.M.; Onilude, A.A. Enzymatic Reduction of Anti-Nutritional Factors in Fermenting Soybeans by Lactobacillus Plantarum Isolates from Fermenting Cereals. Niger. Food J. 2013, 31, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant Food Anti-Nutritional Factors and Their Reduction Strategies: An Overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef]
- Hubert, J.; Berger, M.; Dayde, J. Effects of Fermentation on the Phytochemical Composition and Antioxidant Properties of Soy Germ. Food Chem. 2008, 109, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Ahnan-Winarno, A.D.; Cordeiro, L.; Winarno, F.G.; Gibbons, J.; Xiao, H. Tempeh: A Semicentennial Review on Its Health Benefits, Fermentation, Safety, Processing, Sustainability, and Affordability. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1717–1767. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-H.; Lee, Y.-L.; Li, R.-C.; Mau, J.-L. Non-Volatile Flavour Components of Ganoderma Tsugae. Food Chem. 2005, 90, 409–415. [Google Scholar] [CrossRef]
- Bavaro, S.L.; Susca, A.; Frisvad, J.C.; Tufariello, M.; Chytiri, A.; Perrone, G.; Mita, G.; Logrieco, A.F.; Bleve, G. Isolation, Characterization, and Selection of Molds Associated to Fermented Black Table Olives. Front. Microbiol. 2017, 8, 1356. [Google Scholar] [CrossRef] [Green Version]
- Hartanti, A.T.; Rahayu, G.; Hidayat, I. Rhizopus Species from Fresh Tempeh Collected from Several Regions in Indonesia. Hayati J. Biosci. 2015, 22, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. Introduction to Foodborne Fungi, 4th ed.; Centraalbureau voor Schimmelcultures: Baarn, The Netherlands, 1995. [Google Scholar]
- Azeke, M.A.; Fretzdorff, B.; Buening-Pfaue, H.; Betsche, T. Comparative Effect of Boiling and Solid Substrate Fermentation Using the Tempeh Fungus (Rhizopus Oligosporus) on the Flatulence Potential of African Yambean (Sphenostylis Stenocarpa L.) Seeds. Food Chem. 2007, 103, 1420–1425. [Google Scholar] [CrossRef]
- Nurrahman, N.; Astusti, M.; Suparmo; Soesatyo, M.H.N.E. Pertumbuhan Jamur, Sifat Organoleptik Dan Aktivitas Antioksidan Tempe Kedelai Hitam Yang Diproduksi Dengan Berbagai Jenis Inokulum. Agritech 2012, 32, 60–65. [Google Scholar]
- Parris, N.; Foglia, T.A. Simplified Alcoholic Extraction Procedure for Ammonia in Meat Tissue. J. Agric. Food Chem. 1983, 31, 887–889. [Google Scholar] [CrossRef]
- Meilgaard, M.; Civille, G.V.; Carr, B.T. Sensory Evaluation Techniques, 4th ed.; CRC Press: London, UK, 2006; ISBN 9788578110796. [Google Scholar]
- Tamang, J.P.; Kort, R.; Hutkins, R.; Cotter, P.D.; Endo, A.; Liu, S.Q.; Mayo, B. Fermented Foods in a Global Age: East Meets West. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, B.; Ray, R.R. Current Commercial Perspective of Rhizopus Oryzae: A Review. J. Appl. Sci. 2011, 11, 2470–2486. [Google Scholar] [CrossRef] [Green Version]
- Pedizzi, C.; Lema, J.M.; Carballa, M. Enhancing Thermophilic Co-Digestion of Nitrogen-Rich Substrates by Air Side-Stream Stripping. Bioresour. Technol. 2017, 241, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Varzakas, T.H.; Pyle, D.L.; Niranjan, K. Mycelial penetration and enzymic diffusion on soybean tempe. In Advances in Solid State Fermentation; Roussos, S., Lonsane, B.K., Raimbault, M., Gonzalez, G.V., Eds.; Springer: Berlin, Germany, 1997; pp. 59–70. [Google Scholar]
- Higasa, S.; Negishi, Y.; Aoyagi, Y.; Sugahara, T. Changes in Free Amino Acids of Tempe during Preparation with Velvet Beans (Mucuna Pruriens). J. Jpn. Soc. Food Sci. Technol.—Nippon Shokuhin Kagaku Kogaku Kaishi 1996, 43, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Schindler, A.; Dunkel, A.; Staehler, F.; Backes, M.; Ley, J.; Meyerhof, W.; Hofmann, T. Discovery of Salt Taste Enhancing Arginyl Dipeptides in Protein Digests and Fermented Fish Sauces by Means of a Sensomics Approach. J. Agric. Food Chem. 2011, 59, 12578–12588. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Shiga, K.; Kodama, Y.; Imamura, M.; Uchida, R.; Obata, A.; Fukusaki, E. Analysis of the Correlation between Dipeptides and Taste Differences among Soy Sauces by Using Metabolomics-Based Component Profiling. J. Biosci. Bioeng. 2014, 118, 56–63. [Google Scholar] [CrossRef]
- Arai, S.; Noguchi, M.; Kurosawa, S.; Kato, H.; Fujimaki, M. Applying Proteolytic Enzymes on Soybean. 6. Deodorization Effect of Aspergillopeptidase-A and Debittering Effect of Aspergillus Acid Carboxypeptidase. J. Food Sci. 1970, 35, 392–395. [Google Scholar] [CrossRef]
- Sparringa, R.A.; Owens, J.D. Protein Utilization during Soybean Tempe Fermentation. J. Agric. Food Chem. 1999, 47, 4375–4378. [Google Scholar] [CrossRef] [PubMed]
- Visessanguan, W.; Benjakul, S.; Potachareon, W.; Panya, A.; Riebroy, S. Accelerated Proteolysis of Soy Proteins during Fermentation of Thua-nao Inoculated with Bacillus Subtilis. J. Food Biochem. 2005, 29, 349–366. [Google Scholar] [CrossRef]
- Shurtleff, W.; Aoyagi, A. The Book of Tempeh; Soyinfo Center: New York, NY, USA, 2001. [Google Scholar]
- Santhirasegaram, V.; George, D.S.; Anthony, K.K.; Kaur, H.; Singh, B.; Saruan, N.M.; Razali, Z.; Somasundram, C. Effects of Soybean Processing and Packaging on the Quality of Commonly Consumed Local Delicacy Tempe. J. Food Qual. 2016, 39, 675–684. [Google Scholar] [CrossRef]
- Barus, T.; Suwanto, A.; Wahyudi, A.T.; Wijaya, H. Role of Bacteria in Tempe Bitter Taste Formation; Microbiological and Molecular Biological Analysis Based on 16S RRNA Gene. Microbiol. Indones. 2008, 2, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Efriwati, S.A.; Rahayu, G.; Nuraida, L. Population Dynamics of Yeasts and Lactic Acid Bacteria (LAB) during Tempeh Production. Hayati J. Biosci. 2013, 20, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Ashenafi, M.; Busse, M. Growth of Bacillus Cereus in Fermenting Tempeh Made from Various Beans and Its Inhibition by Lactobacillus Plantarum. J. Appl. Microbiol. 1991, 70, 329–333. [Google Scholar]
- Arroyo-López, F.N.; Querol, A.; Bautista-Gallego, J.; Garrido-Fernández, A. Role of Yeasts in Table Olive Production. Int. J. Food Microbiol. 2008, 128, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Cheirsilp, B.; Shimizu, H.; Shioya, S. Enhanced Kefiran Production by Mixed Culture of Lactobacillus Kefiranofaciens and Saccharomyces Cerevisiae. J. Biotechnol. 2003, 100, 43–53. [Google Scholar] [CrossRef]
- Moreno, M.R.F.; Leisner, J.J.; Tee, L.K.; Ley, C.; Radu, S.; Rusul, G.; Vancanneyt, M.; De Vuyst, L. Microbial Analysis of Malaysian Tempeh, and Characterization of Two Bacteriocins Produced by Isolates of Enterococcus Faecium. J. Appl. Microbiol. 2002, 92, 147–157. [Google Scholar] [CrossRef]
- Nout, M.J.R.; Kiers, J.L. Tempe Fermentation, Innovation and Functionality: Update into the Third Millenium. J. Appl. Microbiol. 2005, 98, 789–805. [Google Scholar] [CrossRef]
- Hachmeister, K.A.; Fung, D. Tempeh: A Mold-Modified Indigenous Fermented Food Made from Soybeans and/or Cereal Grains. Crit. Rev. Microbiol. 1993, 19, 137–188. [Google Scholar] [CrossRef]
- Kadar, A.D.; Astawan, M.; Putri, S.P.; Fukusaki, E. Metabolomics Based Study of the Effect of Raw Materials to the End Product of Tempe—an Indonesian Fermented Soybean. Metabolites 2020, 10, 367. [Google Scholar] [CrossRef]
- Handoyo, T.; Morita, N. Structural and Functional Properties of Fermented Soybean (Tempeh) by Using Rhizopus Oligosporus. Int. J. Food Prop. 2006, 9, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Wikandari, R.; Utami, T.A.N.; Hasniah, N.; Sardjono. Chemical, Nutritional, Physical and Sensory Characterization of Tempe Made from Various Underutilized Legumes. Pakistan J. Nutr. 2020, 19, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, E.; Benoit, I.; van den Brink, J.; Wiebenga, A.; Coutinho, P.M.; Henrissat, B.; de Vries, R.P. Carbohydrate-Active Enzymes from the Zygomycete Fungus Rhizopus Oryzae: A Highly Specialized Approach to Carbohydrate Degradation Depicted at Genome Level. BMC Genom. 2011, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Radiati, A.; Sumarto, S. Analisis Sifat Fisik, Sifat Organoleptik, dan Kandungan Gizi Pada Produk Tempe dari Kacang Non-Kedelai. J. Apl. Teknol. Pangan 2015, 5, 16–22. [Google Scholar]
- Affandi, D.R.; Amanto, B.S.; Ishartani, D.; Nurani, S. Karakteristik Sensoris Tempe Koro Glinding (Phaseolus Lunatus) Pada Berbagai Waktu Fermentasi. In Proceedings of the Peranan Sumber Daya Pertanian, Perkebunan, dan Peternakan dalam Mendukung Ketahanan Pangan Nasional, Fakultas Pertanian UNS, Surakarta, Indonesia, 30 March 2017; pp. 794–799. [Google Scholar]
- Dewi, M.; Tirtaningtyas, P.; Raissha, T.; Kartika, E.; Hanny, C.; Mutukumira, A.N. Sensory Characteristics of Seasoning Powders from Overripe Tempeh, a Solid State Fermented Soybean. Procedia Chem. 2015, 14, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Utami, R.; Wijaya, C.H.; Lioe, H.N. Taste of Water-Soluble Extracts Obtained from Over-Fermented Tempe. Int. J. Food Prop. 2016, 19, 2063–2073. [Google Scholar] [CrossRef]
- Athaillah, Z.A.; Muzdalifah, D.; Lestari, A.; Fitria, A.; Udin, L.Z.; Artanti, N.; Lioe, H.N. Phenolic Compound Profile and Functionality of Aqueous Overripe Tempe Extracts. Curr. Res. Nutr. Food Sci. 2019, 7, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged Tempe-Type Fermentation in Order to Improve Bioactive Potential and Nutritional Parameters of Quinoa Seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Spanier, A.M.; Shahidi, F.; Parliment, T.H.; Mussinan, C.; Ho, C.-T.; Contis, E.T. Food Flavors and Chemistry: Advances of the New Millennium; The Royal Society of Chemistry: Cambridge, UK, 2007. [Google Scholar]
- Soccol, C.R.; Stonoga, V.I.; Raimbault, M. Production of L-Lactic Acid by Rhizopus Species. World J. Microbiol. Biotechnol. 1994, 10, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Suliantari; Suryaatmadja, S.L.; Kusumaningrum, H. Microbial Content and Diversity in Several Tempe from Bogor Areas. In Proceedings of the Prosiding Seminar Hasil-Hasil PPM IPB, Bogor, Indonesia, 1–2 December 2015; pp. 229–237. [Google Scholar]
- de Oliveira, P.M.; Santos, L.P.; Coelho, L.F.; Avila Neto, P.M.; Sass, D.C.; Contiero, J. Production of L (+) Lactic Acid by Lactobacillus Casei Ke11: Fed Batch Fermentation Strategies. Fermentation 2021, 7, 151. [Google Scholar] [CrossRef]
- Hartanti, A.T.; Raharjo, A.; Gunawan, A.W. Rhizopus Rotting on Agricultural Products in Jakarta. Hayati J. Biosci. 2020, 27, 37–44. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Micromorphological Characteristics | Refs. [11,12] | Samples | ||||||
|---|---|---|---|---|---|---|---|---|
| R. oligosporus | R. oryzae | R. delemar | RO1 | RO2 | RO3 | RD | RZ | |
| Sporangiophore | Solitary or 2–4 in groups of sporangiophore. A length of 150–1000 µm, with a diameter of 10–18 µm. | Solitary or 2–5 in groups of Sporangiophore. A length of 150–2000 µm, with a diameter of 6–14 µm and colorless to yellowish-brown. | Solitary or 2–3 in groups, 403.2–812.0 µm long | Solitary Length = 143.5–158.5 µm | Solitary Length = 189–221 µm | In groups Length = 101.3–179.4 µm | In groups, Length = 261–521 µm | Solitary Length = 408 µm |
| Sporangium | Globose or subglobose, brownish-black, diameter of 100–180 µm | Globose or subglobose with spinulose wall; becoming dark brown to black–brown; 50–200 µm in diameter | Globose to subglobose (55–185 µm diameter), solitary or 2–4 in groups and light brown to dark brown | Brownish, globose, 61.2 µm in diameter | Brownish, globose, 65.8–72.8 µm in diameter | Brownish, globose,65.8–72.8 µm in diameter | Light brown, globose, 76.4 µm in diameter | Black–brown, globose, 74.6 µm in diameter |
| Columellae | Globose to subglobose with a funnel shape of the apophysis | Ovoid or globose, 30–120 µm in diameter | ovoid (41.1–85.0 × 37.6–85.7 µm) | Globose 26.2–37.1 µm in diameter | Globose, 29.3 µm in diameter | Diameter = 27.3–44.06 µm | Ovoid Diameter = 36.7–61.1 µm | Diameter = 4557 µm |
| Sporangiospore | Irregularly shaped, globose, ellipsoidal, 7–10 µm in length | Globose, ovoid or irregularly shaped, often polygonal, striate, 4–10 micrometer-long sporangiospores | Ovoid, sometimes subglobose, smooth, with distinct striation (4.8–9.5 µm diameter), irregular, reached 53 μm | Irregularly shaped, 8.3–15.1 µm in diameter | Irregularly shaped, 4.8–18.1 µm in diameter | Irregularly shaped, 6.5–25.5 µm in diameter | Globose, ovoid, 6.4–16. µm in diameter | Polygonal, striate, 4.23–6 µm in diameter |
| Species | Rhizopus oligosporus | Rhizopus oligosporus | Rhizopus oligosporus | Rhizopus delemar | Rhizopus oryzae | |||
| No. | Amino Acid | Concentration (mg/g) | |||||
|---|---|---|---|---|---|---|---|
| RO1 | RO2 | RO3 | RD | RZ | R | ||
| 1 | Glutamic acid | 5.43 | 4.62 | 8.99 | 2.06 | 4.44 | 7.77 |
| 2 | Aspartic acid | 20.97 | 14.64 | 26.07 | 3.38 | 9.34 | 21.16 |
| 3 | Alanine | 2.84 | 1.04 | 7.71 | 5.62 | 2.62 | 5.87 |
| 4 | Arginine | 4.74 | 4.82 | 4.51 | 9.27 | 5.50 | 5.06 |
| 5 | Glycine | 2.77 | 2.38 | 7.77 | 1.07 | 4.77 | 4.37 |
| 6 | Histidine | 8.27 | 6.66 | 9.59 | 14.83 | 11.36 | 12.13 |
| 7 | Isoleucine | 12.12 | 10.97 | 12.05 | 7.37 | 19.36 | 18.91 |
| 8 | Leucine | 4.38 | 6.87 | 3.86 | 5.47 | 7.96 | 5.39 |
| 9 | Lysine | 14.78 | 12.30 | 13.97 | 14.78 | 11.37 | 14.78 |
| 10 | Methionine | 0.74 | 2.57 | 3.22 | 13.47 | 3.39 | 8.62 |
| 11 | Phenylalanine | 0.27 | 8.01 | 13.31 | 8.30 | 20.47 | 12.14 |
| 12 | Serine | 2.21 | 30.23 | 10.79 | 0.89 | 1.84 | 1.73 |
| 13 | Threonine | 6.30 | 5.99 | 33.90 | 11.54 | 28.13 | 8.26 |
| 14 | Tyrosine | 24.40 | 5.29 | 32.80 | 30.10 | 23.30 | 26.39 |
| 15 | Valine | 1.43 | 2.85 | 6.25 | 16.25 | 12.90 | 9.02 |
| Tempe | Appearance | Aroma | Texture | Taste | Overall |
|---|---|---|---|---|---|
| RO1 | 5.1 b* | 5.21 a | 4.86 a | 4.59 ab | 4.93 bc |
| RO2 | 5.28 b | 5.34 a | 5.07 a | 5.1 b | 5.14 c |
| RO3 | 5.34 b | 5.31 a | 5.14 a | 5.1 b | 5.28 c |
| RD | 5.07 b | 5.1 a | 4.62 a | 3.93 a | 4.14 a |
| RZ | 3.86 a | 4.93 a | 4.45 a | 4.45 ab | 4.28 ab |
| R | 5.24 b | 5.24 a | 5 a | 4.9 b | 5.14 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wikandari, R.; Kinanti, D.A.; Permatasari, R.D.; Rahmaningtyas, N.L.; Chairunisa, N.R.; Sardjono; Hellwig, C.; Taherzadeh, M.J. Correlations between the Chemical, Microbiological Characteristics and Sensory Profile of Fungal Fermented Food. Fermentation 2021, 7, 261. https://doi.org/10.3390/fermentation7040261
Wikandari R, Kinanti DA, Permatasari RD, Rahmaningtyas NL, Chairunisa NR, Sardjono, Hellwig C, Taherzadeh MJ. Correlations between the Chemical, Microbiological Characteristics and Sensory Profile of Fungal Fermented Food. Fermentation. 2021; 7(4):261. https://doi.org/10.3390/fermentation7040261
Chicago/Turabian StyleWikandari, Rachma, Dyah Ayu Kinanti, Regina Devi Permatasari, Nur Lisa Rahmaningtyas, Nidya Rizkadianari Chairunisa, Sardjono, Coralie Hellwig, and Mohammad J. Taherzadeh. 2021. "Correlations between the Chemical, Microbiological Characteristics and Sensory Profile of Fungal Fermented Food" Fermentation 7, no. 4: 261. https://doi.org/10.3390/fermentation7040261
APA StyleWikandari, R., Kinanti, D. A., Permatasari, R. D., Rahmaningtyas, N. L., Chairunisa, N. R., Sardjono, Hellwig, C., & Taherzadeh, M. J. (2021). Correlations between the Chemical, Microbiological Characteristics and Sensory Profile of Fungal Fermented Food. Fermentation, 7(4), 261. https://doi.org/10.3390/fermentation7040261