

Lactobacilli and Moesziomyces Biosurfactants: Toward a Closed-Loop Approach for the Dairy Industry

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Cheese Whey Fermentations for Biosurfactants Production
2.2.1. Lactobacilli Biosurfactant Production
2.2.2. MEL Production
2.3. Extraction of Biosurfactants
2.4. β-Galactosidase Assay
2.5. Surface Tension Measurement
2.6. Fermented Milk Production
2.7. Microbiological Analysis
2.8. Physicochemical Analysis
2.9. Analytical Methods
2.10. Statistical Analysis
3. Results and Discussion
3.1. Production of Lactobacilli Biosurfactants (LAB-BS) Using Different Strains
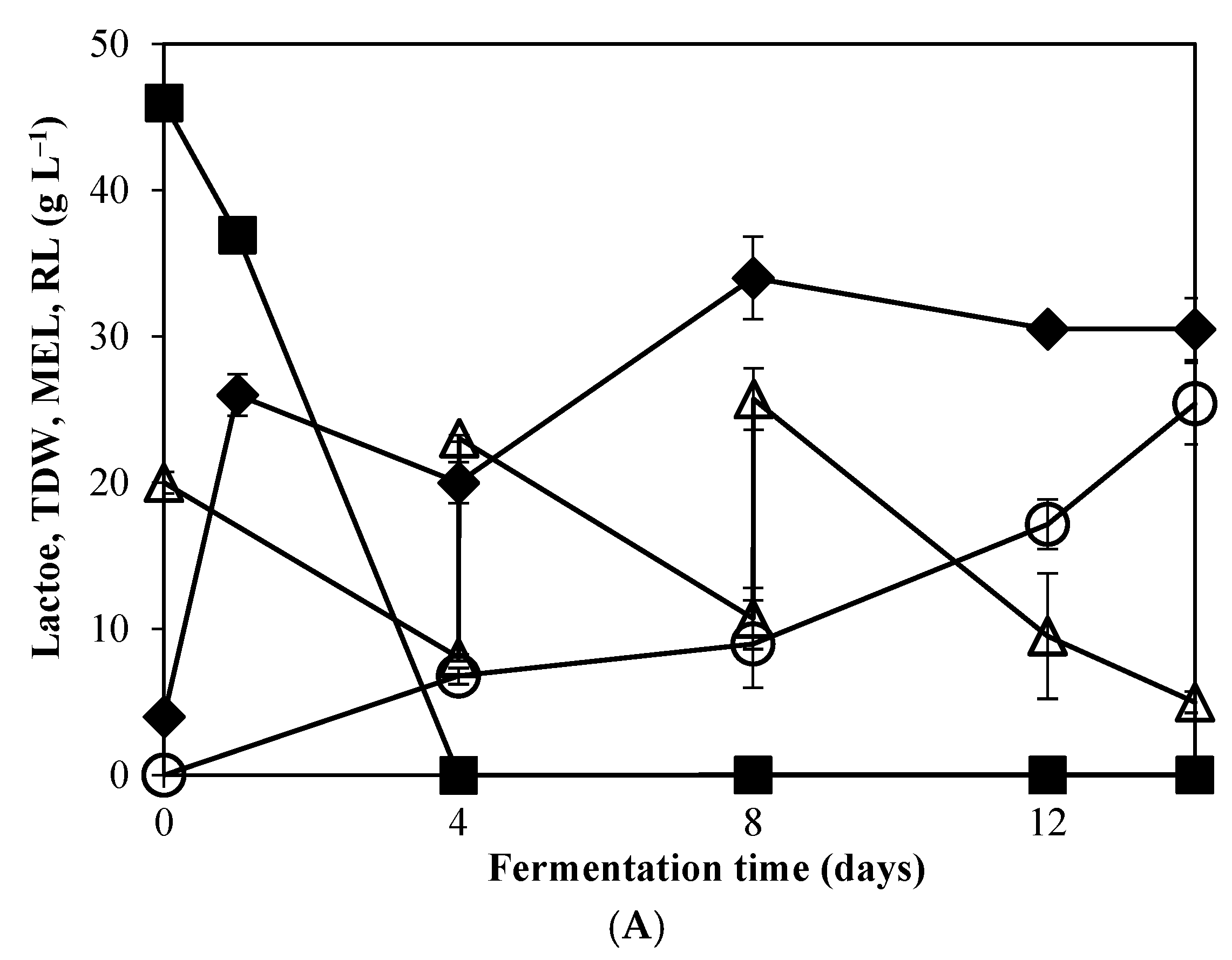
3.2. Production of MEL Biosurfactants
3.3. Fermented Milk Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismail, N.L. Chapter 5. In Surfactants and Detergents; Shahruddin, S., Ed.; IntechOpen: London, UK, 2022; ISBN 978-1-83962-897-9. [Google Scholar]
- Hayes, D.G.; Smith, G.A. Chapter 1—Biobased surfactants: Overview and industrial state of the art. In Biobased Surfactants, 2nd ed.; Hayes, D.G., Solaiman, D.K.Y., Ashby, R.D.B.T.-B.S., Eds.; AOCS Press: Urbana, IL, USA, 2019; pp. 3–38. ISBN 978-0-12-812705-6. [Google Scholar]
- Montoya Vallejo, C.; Flórez Restrepo, M.A.; Guzmán Duque, F.L.; Quintero Díaz, J.C. Production, Characterization and Kinetic Model of Biosurfactant Produced by Lactic Acid Bacteria. Electron. J. Biotechnol. 2021, 53, 14–22. [Google Scholar] [CrossRef]
- Banat, I.M.; Carboué, Q.; Saucedo-Castañeda, G.; de Jesús Cázares-Marinero, J. Biosurfactants: The Green Generation of Speciality Chemicals and Potential Production Using Solid-State Fermentation (SSF) Technology. Bioresour. Technol. 2021, 320, 124222. [Google Scholar] [CrossRef] [PubMed]
- Markande, A.R.; Patel, D.; Varjani, S. A Review on Biosurfactants: Properties, Applications and Current Developments. Bioresour. Technol. 2021, 330, 124963. [Google Scholar] [CrossRef] [PubMed]
- Jimoh, A.A.; Lin, J. Biotechnological Applications of Paenibacillus Sp. D9 Lipopeptide Biosurfactant Produced in Low-Cost Substrates. Appl. Biochem. Biotechnol. 2020, 191, 921–941. [Google Scholar] [CrossRef]
- Ambaye, T.G.; Vaccari, M.; Prasad, S.; Rtimi, S. Preparation, Characterization and Application of Biosurfactant in Various Industries: A Critical Review on Progress, Challenges and Perspectives. Environ. Technol. Innov. 2021, 24, 102090. [Google Scholar] [CrossRef]
- Farias, C.B.B.; Almeida, F.C.G.; Silva, I.A.; Souza, T.C.; Meira, H.M.; Soares da Silva, R.d.C.F.; Luna, J.M.; Santos, V.A.; Converti, A.; Banat, I.M.; et al. Production of Green Surfactants: Market Prospects. Electron. J. Biotechnol. 2021, 51, 28–39. [Google Scholar] [CrossRef]
- Gaur, V.K.; Sharma, P.; Sirohi, R.; Varjani, S.; Taherzadeh, M.J.; Chang, J.-S.; Yong Ng, H.; Wong, J.W.C.; Kim, S.-H. Production of Biosurfactants from Agro-Industrial Waste and Waste Cooking Oil in a Circular Bioeconomy: An Overview. Bioresour. Technol. 2022, 343, 126059. [Google Scholar] [CrossRef]
- Domínguez Rivera, Á.; Martínez Urbina, M.Á.; López y López, V.E. Advances on Research in the Use of Agro-Industrial Waste in Biosurfactant Production. World J. Microbiol. Biotechnol. 2019, 35, 155. [Google Scholar] [CrossRef]
- Ferreira, A.; Vecino, X.; Ferreira, D.; Cruz, J.M.; Moldes, A.B.; Rodrigues, L.R. Novel Cosmetic Formulations Containing a Biosurfactant from Lactobacillus Paracasei. Colloids Surf. B Biointerfaces 2017, 155, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Adu, S.A.; Naughton, P.J.; Marchant, R.; Banat, I.M. Microbial Biosurfactants in Cosmetic and Personal Skincare Pharmaceutical Formulations. Pharmaceutics 2020, 12, 1099. [Google Scholar] [CrossRef]
- Kachrimanidou, V.; Papadaki, A.; Lappa, I.; Papastergiou, S.; Kleisiari, D.; Kopsahelis, N. Biosurfactant Production from Lactobacilli: An Insight on the Interpretation of Prevailing Assessment Methods. Appl. Biochem. Biotechnol. 2022, 194, 882–900. [Google Scholar] [CrossRef] [PubMed]
- Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactant-Producing Lactobacilli: Screening, Production Profiles, and Effect of Medium Composition. Appl. Environ. Soil Sci. 2011, 201254. [Google Scholar] [CrossRef] [Green Version]
- Cornea, C.P.; Roming, F.I.; Sicuia, O.A.; Voaideş, C.; Zamfir, M.; Grosu-Tudor, S.S. Biosurfactant Production by Lactobacillus Spp. Strains Isolated from Romanian Traditional Fermented Food Products. Rom. Biotechnol. Lett. 2016, 21, 11312–11320. [Google Scholar]
- Schoina, V.; Terpou, A.; Papadaki, A.; Bosnea, L.; Kopsahelis, N.; Kanellaki, M. Enhanced Aromatic Profile and Functionality of Cheese Whey Beverages by Incorporation of Probiotic Cells Immobilized on Pistacia Terebinthus Resin. Foods 2020, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Dimitrellou, D.; Kandylis, P.; Kourkoutas, Y. Assessment of Freeze-Dried Immobilized Lactobacillus Casei as Probiotic Adjunct Culture in Yogurts. Foods 2019, 8, 374. [Google Scholar] [CrossRef] [Green Version]
- Dimitrellou, D.; Salamoura, C.; Kontogianni, A.; Katsipi, D.; Kandylis, P.; Zakynthinos, G.; Varzakas, T. Effect of Milk Type on the Microbiological, Physicochemical and Sensory Characteristics of Probiotic Fermented Milk. Microorganisms 2019, 7, 274. [Google Scholar] [CrossRef] [Green Version]
- Sakandar, H.A.; Zhang, H. Trends in Probiotic(s)-Fermented Milks and Their in Vivo Functionality: A Review. Trends Food Sci. Technol. 2021, 110, 55–65. [Google Scholar] [CrossRef]
- Gallardo-Escamilla, F.J.; Kelly, A.L.; Delahunty, C.M. Mouthfeel and Flavour of Fermented Whey with Added Hydrocolloids. Int. Dairy J. 2007, 17, 308–315. [Google Scholar] [CrossRef]
- Teimouri, S.; Abbasi, S.; Scanlon, M.G. Stabilisation Mechanism of Various Inulins and Hydrocolloids: Milk–Sour Cherry Juice Mixture. Int. J. Dairy Technol. 2018, 71, 208–215. [Google Scholar] [CrossRef]
- Mouafo, H.T.; Sokamte, A.T.; Mbawala, A.; Ndjouenkeu, R.; Devappa, S. Biosurfactants from Lactic Acid Bacteria: A Critical Review on Production, Extraction, Structural Characterization and Food Application. Food Biosci. 2022, 46, 101598. [Google Scholar] [CrossRef]
- Zouari, R.; Besbes, S.; Ellouze-Chaabouni, S.; Ghribi-Aydi, D. Cookies from Composite Wheat–Sesame Peels Flours: Dough Quality and Effect of Bacillus Subtilis SPB1 Biosurfactant Addition. Food Chem. 2016, 194, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, A.; Kiran, G.S.; Selvin, J. Revealing the Effect of Lipopeptide on Improving the Probiotics Characteristics: Flavor and Texture Enhancer in the Formulated Yogurt. Food Chem. 2022, 375, 131718. [Google Scholar] [CrossRef]
- Ribeiro, B.G.; Guerra, J.M.C.; Sarubbo, L.A. Potential Food Application of a Biosurfactant Produced by Saccharomyces Cerevisiae URM 6670. Front. Bioeng. Biotechnol. 2020, 8, 434. [Google Scholar] [CrossRef]
- López-Prieto, A.; Rodríguez-López, L.; Rincón-Fontán, M.; Moldes, A.B.; Cruz, J.M. Effect of Biosurfactant Extract Obtained from the Corn-Milling Industry on Probiotic Bacteria in Drinkable Yogurt. J. Sci. Food Agric. 2019, 99, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Kachrimanidou, V.; Alimpoumpa, D.; Papadaki, A.; Lappa, I.; Alexopoulos, K.; Kopsahelis, N. Cheese Whey Utilization for Biosurfactant Production: Evaluation of Bioprocessing Strategies Using Novel Lactobacillus Strains. Biomass Convers. Biorefinery 2022, 12, 4621–4635. [Google Scholar] [CrossRef]
- Nascimento, M.F.; Barreiros, R.; Oliveira, A.C.; Ferreira, F.C.; Faria, N.T. Moesziomyces Spp. Cultivation Using Cheese Whey: New Yeast Extract-Free Media, β-Galactosidase Biosynthesis and Mannosylerythritol Lipids Production. Biomass Convers. Biorefinery 2022, 1–14. [Google Scholar] [CrossRef]
- Sharma, D.; Saharan, B.S.; Chauhan, N.; Bansal, A.; Procha, S. Production and Structural Characterization of Lactobacillus Helveticus Derived Biosurfactant. Sci. World J. 2014, 493548. [Google Scholar] [CrossRef] [Green Version]
- Faria, N.T.; Santos, M.; Ferreira, C.; Marques, S.; Ferreira, F.C.; Fonseca, C. Conversion of Cellulosic Materials into Glycolipid Biosurfactants, Mannosylerythritol Lipids, by Pseudozyma Spp. under SHF and SSF Processes. Microb. Cell Fact. 2014, 13, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Splechtna, B.; Steinböck, M.; Kneifel, W.; Lettner, H.P.; Kulbe, K.D.; Haltrich, D. Purification and Characterization of Two Novel β-Galactosidases from Lactobacillus Reuteri. J. Agric. Food Chem. 2006, 54, 4989–4998. [Google Scholar] [CrossRef]
- Osman, A.; Tzortzis, G.; Rastall, R.A.; Charalampopoulos, D. BbgIV Is an Important Bifidobacterium β-Galactosidase for the Synthesis of Prebiotic Galactooligosaccharides at High Temperatures. J. Agric. Food Chem. 2012, 60, 740–748. [Google Scholar] [CrossRef]
- Terpou, A.; Gialleli, A.I.; Bekatorou, A.; Dimitrellou, D.; Ganatsios, V.; Barouni, E.; Koutinas, A.A.; Kanellaki, M. Sour Milk Production by Wheat Bran Supported Probiotic Biocatalyst as Starter Culture. Food Bioprod. Process. 2017, 101, 184–192. [Google Scholar] [CrossRef]
- Kachrimanidou, V.; Kopsahelis, N.; Vlysidis, A.; Papanikolaou, S.; Kookos, I.K.; Monje Martínez, B.; Escrig Rondán, M.C.; Koutinas, A.A. Downstream Separation of Poly(Hydroxyalkanoates) Using Crude Enzyme Consortia Produced via Solid State Fermentation Integrated in a Biorefinery Concept. Food Bioprod. Process. 2016, 100, 323–334. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of Potential Probiotic Lactic Acid Bacteria from Fermented Olives by in Vitro Tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Production of Mannosylerythritol Lipids and Their Application in Cosmetics. Appl. Microbiol. Biotechnol. 2013, 97, 4691–4700. [Google Scholar] [CrossRef]
- El Bouchikhi, S.; Pagès, P.; El Alaoui, Y.; Ibrahimi, A.; Bensouda, Y. Syneresis Investigations of Lacto-Fermented Sodium Caseinate in a Mixed Model System. BMC Biotechnol. 2019, 19, 57. [Google Scholar] [CrossRef]
- Tian, Q.; Lai, L.; Zhou, Z.; Mei, P.; Lu, Q.; Wang, Y.; Xiang, D.; Liu, Y. Interaction Mechanism of Different Surfactants with Casein: A Perspective on Bulk and Interfacial Phase Behavior. J. Agric. Food Chem. 2019, 67, 6336–6349. [Google Scholar] [CrossRef] [PubMed]
- Albadran, H.A.; Chatzifragkou, A.; Khutoryanskiy, V.V.; Charalampopoulos, D. Development of Surfactant-Coated Alginate Capsules Containing Lactobacillus Plantarum. Food Hydrocoll. 2018, 82, 490–499. [Google Scholar] [CrossRef]
- Domagała, J. Instrumental Texture, Syneresis and Microstructure of Yoghurts Prepared from Goat, Cow and Sheep Milk. Int. J. Food Prop. 2009, 12, 605–615. [Google Scholar] [CrossRef]
- Joon, R.; Mishra, S.K.; Singh Brar, G.; Kumar Singh, P.; Kumar Mishra, S.; Panwar, H. Instrumental Texture and Syneresis Analysis of Yoghurt Prepared from Goat and Cow Milk. Pharma Innov. J. 2017, 6, 971–974. [Google Scholar]
- Gomes, J.J.L.; Duarte, A.M.; Batista, A.S.M.; de Figueiredo, R.M.F.; de Sousa, E.P.; de Souza, E.L.; Queiroga, R.d.C.R.d.E. Physicochemical and Sensory Properties of Fermented Dairy Beverages Made with Goat’s Milk, Cow’s Milk and a Mixture of the Two Milks. LWT Food Sci. Technol. 2013, 54, 18–24. [Google Scholar] [CrossRef]
- Sharma, D.; Saharan, B.S.; Kapil, S. Biosurfactants of Probiotic Lactic Acid Bacteria; Springer: Cham, Switzerland, 2016; ISBN 9783319262130. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CWP + VO (Supplemented) | CWP + VO |
|---|---|---|
| Fermentation time (days) | 14 | 8 |
| TDWmax (g L−1) | 30.50 ± 2.12 | 42.00 ± 4.24 |
| MELmax (g L−1) | 25.40 ± 2.79 | 13.75 ± 1.99 |
| YMEL/S (g g−1) | 0.25 ± 0.03 | 0.17 ± 0.02 |
| qpmax (g L−1 d−1) | 1.81 ± 0.20 | 1.72 ± 0.25 |
| MEL purity (g g−1) | 0.84 ± 0.01 | 0.47 ± 0.14 |
| Type of Fermented Milk | log10CFU/mL | Syneresis (%) | Titratable Acidity (Lactic Acid/100 g Fermented Product) | |||
|---|---|---|---|---|---|---|
| Whole cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.08 ± 0.13 | 8.42 ± 0.13 | 34.81 ± 0.70 | 50.04 ± 1.27 | 0.36 ± 0.02 | 0.43 ± 0.02 |
| LAB-BS | 8.11 ± 0.12 | 8.60 ± 0.13 | 34.91 ± 1.01 | 51.71 ± 1.42 | 0.35 ± 0.01 | 0.48 ± 0.02 |
| MEL | 8.12 ± 0.14 | 8.37 ± 0.14 | 34.39 ± 1.49 | 47.09 ± 1.44 | 0.33 ± 0.01 | 0.41 ± 0.02 |
| Semi skimmed cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.05 ± 0.13 | 8.22 ± 0.13 | 36.30 ± 1.62 | 57.06 ± 1.94 | 0.40 ± 0.02 | 0.52 ± 0.03 |
| LAB-BS | 8.06 ± 0.13 | 8.43 ± 0.14 | 35.64 ± 1.34 | 59.10 ± 1.61 | 0.42 ± 0.03 | 0.54 ± 0.03 |
| MEL | 8.07 ± 0.13 | 8.19 ± 0.13 | 36.78 ± 1.38 | 55.20 ± 1.78 | 0.43 ± 0.02 | 0.50 ± 0.03 |
| Type of Fermented Milk | log10CFU/mL | Syneresis (%) | Titratable Acidity (Lactic Acid/100 g Fermented Product) | |||
|---|---|---|---|---|---|---|
| Whole cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.29 ± 0.18 | 8.70 ± 0.15 | 37.68 ± 0.91 | 55.00 ± 1.65 | 0.39 ± 0.03 | 0.66 ± 0.03 |
| LAB-BS | 8.36 ± 0.14 | 8.95 ± 0.18 | 38.90 ± 1.01 | 58.36 ± 1.77 | 0.42 ± 0.03 | 0.68 ± 0.03 |
| MEL | 8.18 ± 0.13 | 8.66 ± 0.13 | 36.27 ± 1.52 | 53.48 ± 1.25 | 0.37 ± 0.02 | 0.62 ± 0.03 |
| Semi skimmed cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.14 ± 0.14 | 8.39 ± 0.13 | 39.23 ± 1.62 | 59.94 ± 2.39 | 0.37 ± 0.04 | 0.56 ± 0.02 |
| LAB-BS | 8.16 ± 0.13 | 8.51 ± 0.14 | 40.54 ± 1.34 | 61.53 ± 1.91 | 0.41 ± 0.03 | 0.60 ± 0.05 |
| MEL | 8.14 ± 0.14 | 8.33 ± 0.13 | 39.51 ± 1.38 | 57.56 ± 2.33 | 0.36 ± 0.04 | 0.54 ± 0.04 |
| Type of Fermented Milk | log10CFU/mL | Syneresis (%) | Titratable Acidity (Lactic Acid/100 g Fermented Product) | |||
|---|---|---|---|---|---|---|
| Whole cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.68 ± 0.21 | 9.28 ± 0.14 | 51.57 ± 0.48 | 57.18 ± 1.36 | 0.59 ± 0.04 | 0.72 ± 0.04 |
| LAB-BS | 8.71 ± 0.18 | 9.58 ± 0.13 | 52.31 ± 0.90 | 59.89 ± 1.16 | 0.61 ± 0.03 | 0.79 ± 0.05 |
| MEL | 8.69 ± 0.14 | 8.95 ± 0.14 | 48.47 ± 0.73 | 55.35 ± 1.26 | 0.55 ± 0.02 | 0.64 ± 0.03 |
| Semi skimmed cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.31 ± 0.09 | 8.60 ± 0.14 | 53.53 ± 1.95 | 61.79± 1.72 | 0.48 ± 0.04 | 0.61 ± 0.06 |
| LAB-BS | 8.34 ± 0.11 | 8.69 ± 0.12 | 55.00 ± 1.27 | 65.94 ± 1.68 | 0.53 ± 0.05 | 0.66 ± 0.04 |
| MEL | 8.28 ± 0.08 | 8.58 ± 0.14 | 51.90 ± 1.29 | 58.13 ± 1.31 | 0.46 ± 0.05 | 0.57 ± 0.03 |
| Cell recycling whole cow milk | Day 0 | Day 20 | Day 0 | Day 20 | Day 0 | Day 20 |
| Control | 8.02 ± 0.09 | 8.14 ± 0.14 | 34.75 ± 1.87 | 42.01 ± 1.12 | 0.33 ± 0.02 | 0.35 ± 0.04 |
| LAB-BS | 8.07 ± 0.13 | 8.38 ± 0.13 | 36.32 ± 1.26 | 43.12 ± 0.71 | 0.34 ± 0.02 | 0.43 ± 0.02 |
| MEL | 8.01 ± 0.13 | 8.09 ± 0.12 | 34.40 ± 1.04 | 41.68 ± 1.34 | 0.30 ± 0.03 | 0.32 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachrimanidou, V.; Alexandri, M.; Nascimento, M.F.; Alimpoumpa, D.; Torres Faria, N.; Papadaki, A.; Castelo Ferreira, F.; Kopsahelis, N. Lactobacilli and Moesziomyces Biosurfactants: Toward a Closed-Loop Approach for the Dairy Industry. Fermentation 2022, 8, 517. https://doi.org/10.3390/fermentation8100517
Kachrimanidou V, Alexandri M, Nascimento MF, Alimpoumpa D, Torres Faria N, Papadaki A, Castelo Ferreira F, Kopsahelis N. Lactobacilli and Moesziomyces Biosurfactants: Toward a Closed-Loop Approach for the Dairy Industry. Fermentation. 2022; 8(10):517. https://doi.org/10.3390/fermentation8100517
Chicago/Turabian StyleKachrimanidou, Vasiliki, Maria Alexandri, Miguel Figueiredo Nascimento, Dimitra Alimpoumpa, Nuno Torres Faria, Aikaterini Papadaki, Frederico Castelo Ferreira, and Nikolaos Kopsahelis. 2022. "Lactobacilli and Moesziomyces Biosurfactants: Toward a Closed-Loop Approach for the Dairy Industry" Fermentation 8, no. 10: 517. https://doi.org/10.3390/fermentation8100517
APA StyleKachrimanidou, V., Alexandri, M., Nascimento, M. F., Alimpoumpa, D., Torres Faria, N., Papadaki, A., Castelo Ferreira, F., & Kopsahelis, N. (2022). Lactobacilli and Moesziomyces Biosurfactants: Toward a Closed-Loop Approach for the Dairy Industry. Fermentation, 8(10), 517. https://doi.org/10.3390/fermentation8100517