
Nonconventional Yeasts Engineered Using the CRISPR-Cas System as Emerging Microbial Cell Factories




Abstract
:1. Introduction
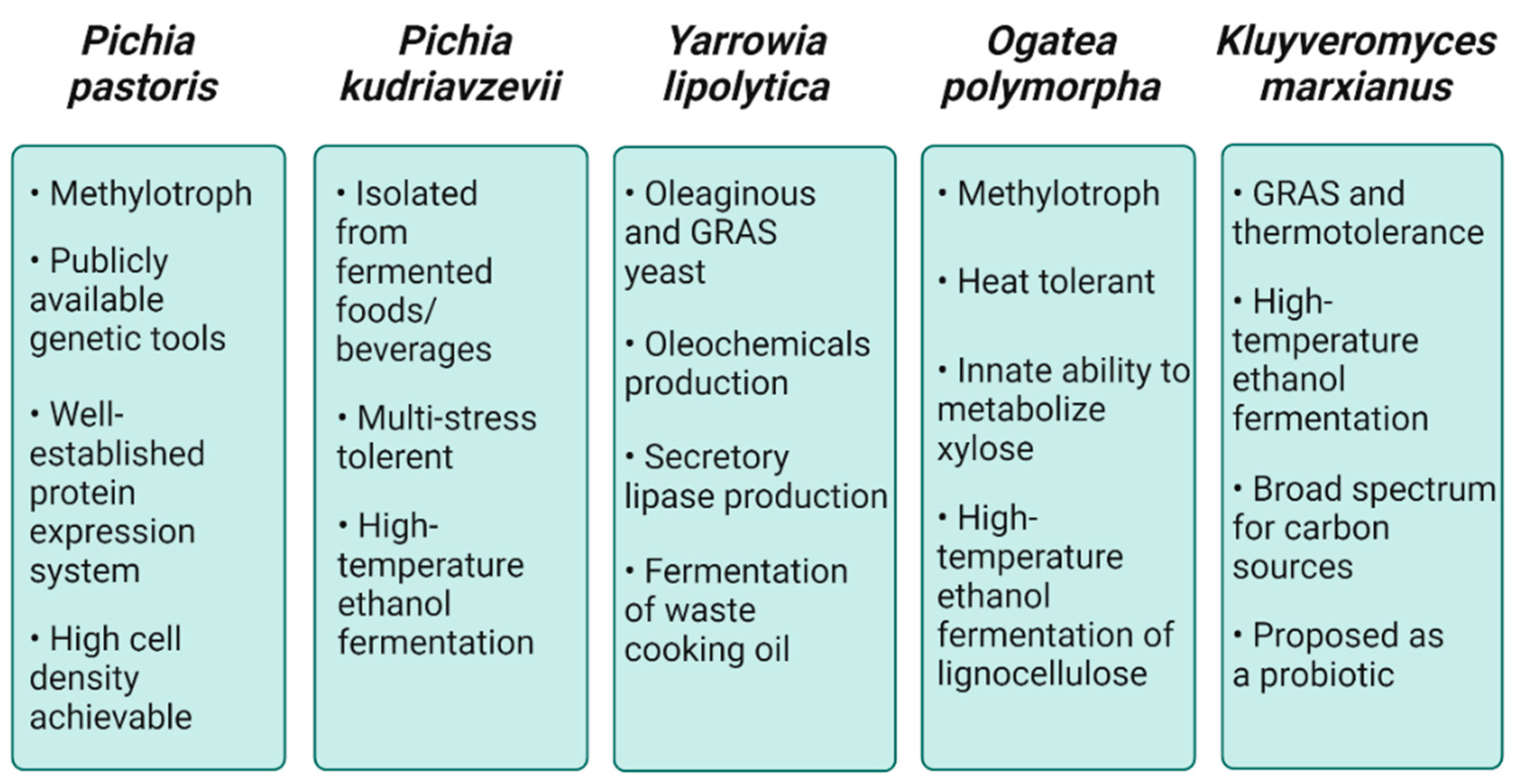
2. Industrial Value of NCYs
2.1. Pichia pastoris
2.2. Pichia kudriavzevii
2.3. Yarrowia lipolytica
2.4. Ogataea polymorpha
2.5. Kluyveromyces marxianus
3. Genetic Engineering Tools for NCYs
4. CRISPR-Cas System-Guided Metabolic Engineering in NCYs
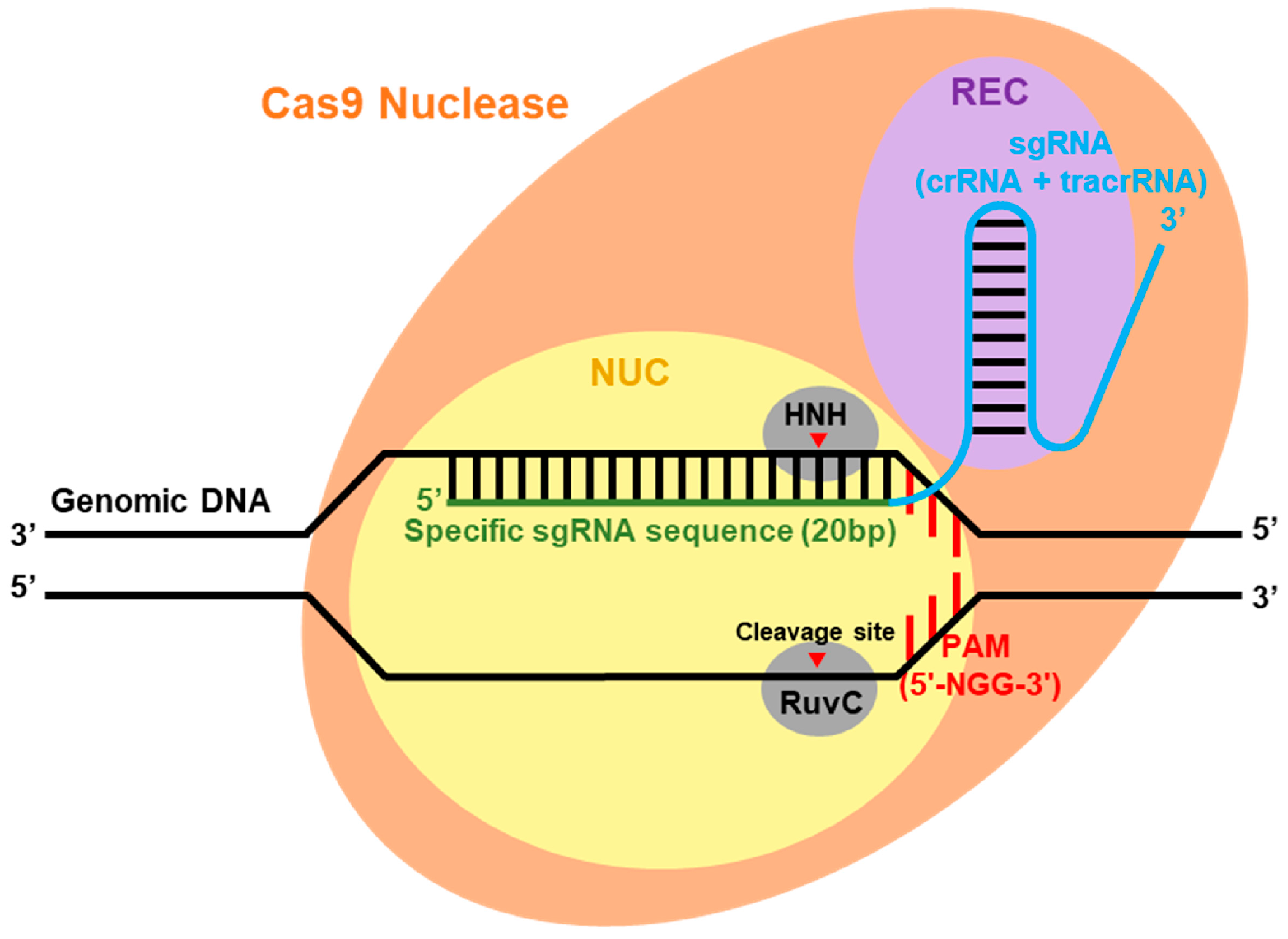
4.1. CRISPR-Cas System: Classification, Components, and Mechanism
4.2. Challenges and Strategies of CRISPR-Cas9-Guided Genome Editing in NCYs

{kind=link}
{kind=link}
{kind=link}
| Strain | sgRNA Promoter | Plasmid (Backbone) | Cas9 Promoter | Editing Efficiency 1 (%) | Reference |
|---|---|---|---|---|---|
| Y. lipolytica | SCR1′-tRNA2 TEFin (Pol II) 3 | pCRISPRyl pCASyl pGGA | TEF1 | 0–68.9 | [71,72,73,74] |
| O. polymorpha | ScSNR52 (Pol III) ScTDH3 (Pol III) | pCRCT pYTK079 | ScTEF1 AaTEF | 1–75 | [75,76] |
| P. pastoris | HTX1 (Pol II) 3 PFK300 (Pol II) 3 LAT1(Pol II) 3 | pPpT4 p414 BB3cH | HTX1 GAP | 75–93.8 | [77,78,79] |
| K. marxianus | ScTDH3 (Pol II) 3 RPR1′-tRNA (Pol III) 2 | pYTK079 pIW601 | AaTEF 1 ScTEF1 | 10–82 | [76,80] |
| P. kudriavzevii | RPR1 (Pol III) RPR1′-tRNA (Pol III) 2 | pRS416 pRS415 pCast | TEF1 | 64 | [30,81,82] |
4.3. Biotechnological Application of CRISPR-Cas9-Introduced NCYs
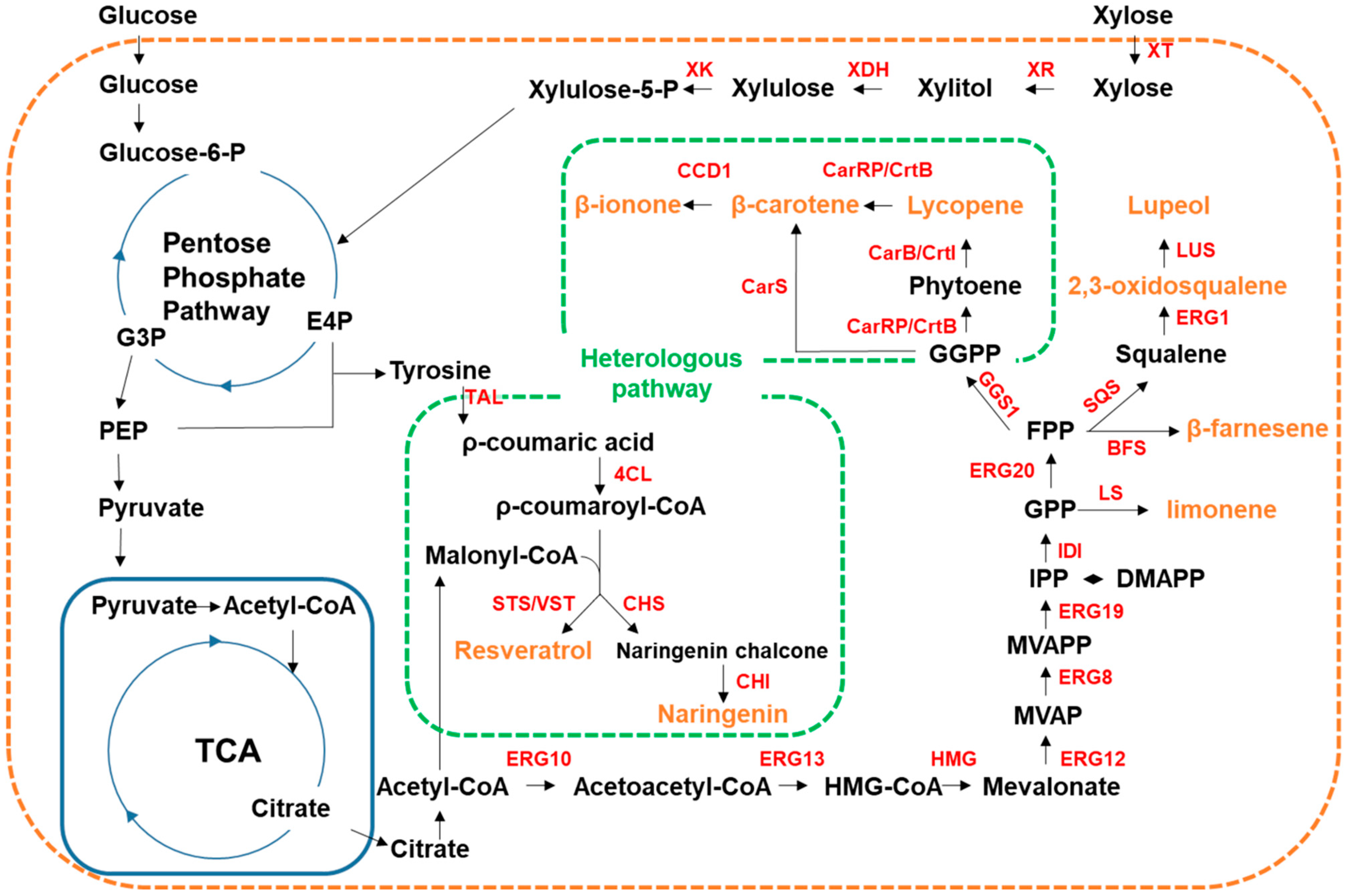
4.3.1. Secondary Plant Metabolites
| Strain | Target Genes | Product | Reference | |
|---|---|---|---|---|
| Endogenous Gene Editing | Heterologous Gene Editing | |||
| Y. lipolytica | HMG1, GGS1 | crtE (Pantoea ananatis), crtI (P. ananatis), crtB (P. ananatis) | Lycopene 3.38 mg/g DCW 2 | [71] |
| GGS1 | carB (Mucor circinelloides), carRP (M. circinelloides), | β-carotene 4.8 mg/g DCW | [72] | |
| GGS1, ERG13, HMG | carB (M. circinelloides), carRP (M. circinelloides) | β-carotene 4.5 g/L | [73] | |
| GGS1, HMG1, ERG8, ERG10, ERG12, ERG13, ERG20, ERG19, IDI | carB (M. circinelloides), carRP (M. circinelloides), CCD1 (Petunia hybrid), PK (Bifidobacterium bifidum), PTA (Bacillus subtilis) | β-ionone 358.4 mg/L 0.98 g/L (fed-batch) | [112] | |
| XK, HMG11, ERG121 | LS (Agastache rugosa) 1, NDPS (Solanum lycopersicum) 1, XR (Scheffersomyces stipitis), XDH (S. stipitis) | Limonene 20.57 mg/L | [113] | |
| HMG, ERG12, IDI, ERG20, SQS | BFS (Artemisia annua), LS (Citrus limon), LS (Perilla frutescens), CnVS (Callitropsis nootkatensis), crtI (Xanthophyllomyces dendrorhous), crtYB (X. dendrohous), acs (Salmonella enterica) | β-farnesene 955 mg/L Limonene 35.9 mg/L Valencene 113.9 mg/L Squalene 402.4 mg/L β-carotene 164 mg/L 2,3-oxidosqualene 22 mg/L | [111] | |
| HMG1, ERG1, ERG9, OLE1, PAH1, DGK1 | LUS (Ricinus communis) | Lupeol 441.72 mg/L | [114] | |
| GGS1 | carS (Schizochytrium sp.) | β-carotene 0.41 mg/g DCW | [74] | |
| XT1, XR1, XDH1, XKS1 | TAL (Rhodotorula glutinis), 4CL (Arabidopsis thaliana), CHS (A. thaliana), CHI (A. thaliana) | Naringenin 715.3 mg/L | [109] | |
| ARO4, ARO7 | TAL (Flavobacterium johnsoniae), VST (Vitis vinifera), 4CL (A. thaliana) | Resveratrol 12.4 g/L | [103] | |
| O. polymorpha | - | TAL (Herpetosiphon aurantiacus), STS (V. vinifera), 4CL (A. thaliana) | Resveratrol 97.23 mg/L | [75] |
4.3.2. Other Industrial Products
| Strain | Target Genes | Product | Reference | |
|---|---|---|---|---|
| Endogenous Gene Editing | Heterologous Gene Editing | |||
| Y. lipolytica | - | CAD (Aspergillus terreus), mttA (A. terreus) 1 | Itaconic acid 22.03 g/L | [116] |
| P. kudriavzevii | ICD, mttA1 | CAD (A. terreus) 1 | Itaconic acid 1.23 g/L | [30] |
| Y. lipolytica | SCT1, OLE1 | FAR (M. aquaeolei) | Fatty alcohol 5.75 g/L | [120] |
| Y. lipolytica | PLA2 | - | Lipid 25 g/L | [119] |
| Y. lipolytica | AXP | celB (Pyrococcus furiosus) | β-glycosidase 187.5 µkatoNPGal/L 2 | [124] |
| K. marxianus | ARO1, ARO2, ARO3, ARO4, ARO7, ARO8, ARO9, PHA2, TAL1, TKL1, RPE1, RKI1, LAC4 | xfpk (Bifidobacterium breve), ppsA (Escherichia coli), pta (Salmonella enterica) | 2-penylethanol 850 mg/L | [48] |
| Y. lipolytica | - | FAP (Chlorella variabilis) | Hydrocarbons 58.7 mg/L | [125] |
| K. marxianus | ACO2b, SDH2, RIP1, MSS51 | - | Ethyl acetate 150 mg/L | [123] |
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Li, W.; Shen, X.; Wang, J.; Sun, X.; Yuan, Q. Engineering microorganisms for the biosynthesis of dicarboxylic acids. Biotechnol. Adv. 2021, 48, 107710. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.; Shishir, M.R.I.; Gowd, V.; Chen, W. Hesperidin-an emerging bioactive compound against metabolic diseases and its potential biosynthesis pathway in microorganism. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Srinivasan, P.; Smolke, C.D. Biosynthesis of medicinal tropane alkaloids in yeast. Nature 2020, 585, 614–619. [Google Scholar] [CrossRef]
- Raschmanová, H.; Weninger, A.; Glieder, A.; Kovar, K.; Vogl, T. Implementing CRISPR-Cas technologies in conventional and non-conventional yeasts: Current state and future prospects. Biotechnol. Adv. 2018, 36, 641–665. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.-S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1. [Google Scholar] [CrossRef]
- Rainha, J.; Gomes, D.; Rodrigues, L.R.; Rodrigues, J.L. Synthetic biology approaches to engineer Saccharomyces cerevisiae towards the industrial production of valuable polyphenolic compounds. Life 2020, 10, 56. [Google Scholar] [CrossRef]
- Rainha, J.; Rodrigues, J.L.; Rodrigues, L.R. CRISPR-Cas9: A powerful tool to efficiently engineer Saccharomyces cerevisiae. Life 2020, 11, 13. [Google Scholar] [CrossRef]
- Kręgiel, D.; Pawlikowska, E.; Antolak, H. Nonconventional yeasts in fermentation processes: Potentialities and limitations. In Old Yeasts: New Questions; IntechOpen: London, UK, 2017; p. 87. [Google Scholar]
- Bertels, L.-K.; Fernández Murillo, L.; Heinisch, J.J. The pentose phosphate pathway in yeasts-More than a poor cousin of glycolysis. Biomolecules 2021, 11, 725. [Google Scholar] [CrossRef]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent advances in systems and synthetic biology approaches for developing novel cell-factories in non-conventional yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar] [CrossRef]
- Jensen, M.K.; Keasling, J.D. Recent applications of synthetic biology tools for yeast metabolic engineering. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.M.; Hochedlinger, K. A variant CRISPR-Cas9 system adds versatility to genome engineering. Proc. Natl. Acad. Sci. USA 2013, 110, 15514–15515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, O.; Arava, Y. Expanding the CRISPR/Cas9 toolbox for gene engineering in S. cerevisiae. Curr. Microbiol. 2020, 77, 468–478. [Google Scholar] [CrossRef]
- Mitsui, R.; Yamada, R.; Ogino, H. CRISPR system in the yeast Saccharomyces cerevisiae and its application in the bioproduction of useful chemicals. World J. Microbiol. Biotechnol. 2019, 35, 111. [Google Scholar] [CrossRef] [PubMed]
- Geijer, C.; Ledesma-Amaro, R.; Tomás-Pejó, E. Unraveling the potential of non-conventional yeasts in biotechnology. FEMS Yeast Res. 2022, 22, foab071. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Gao, J.; Zhou, Y. CRISPR-mediated genome editing in non-conventional yeasts for biotechnological applications. Microb. Cell Fact. 2019, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Rebello, S.; Abraham, A.; Madhavan, A.; Sindhu, R.; Binod, P.; Karthika Bahuleyan, A.; Aneesh, E.M.; Pandey, A. Non-conventional yeast cell factories for sustainable bioprocesses. FEMS Microbiol. Lett. 2018, 365, fny222. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, V.; Radecka, D.; Aerts, G.; Verstrepen, K.J.; Lievens, B.; Thevelein, J.M. Phenotypic landscape of non-conventional yeast species for different stress tolerance traits desirable in bioethanol fermentation. Biotechnol. Biofuels 2017, 10, 216. [Google Scholar] [CrossRef]
- Chen, M.-T.; Lin, S.; Shandil, I.; Andrews, D.; Stadheim, T.A.; Choi, B.-K. Generation of diploid Pichia pastoris strains by mating and their application for recombinant protein production. Microb. Cell Fact. 2012, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Yarimizu, T.; Nonklang, S.; Nakamura, J.; Tokuda, S.; Nakagawa, T.; Lorreungsil, S.; Sutthikhumpha, S.; Pukahuta, C.; Kitagawa, T.; Nakamura, M. Identification of auxotrophic mutants of the yeast Kluyveromyces marxianus by non-homologous end joining-mediated integrative transformation with genes from Saccharomyces cerevisiae. Yeast 2013, 30, 485–500. [Google Scholar] [CrossRef]
- Kao, Y.-T.; Gonzalez, K.L.; Bartel, B. Peroxisome function, biogenesis, and dynamics in plants. Plant Physiol. 2018, 176, 162–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P. Biotechnological strains of Komagataella (Pichia) pastoris are Komagataella phaffii as determined from multigene sequence analysis. J. Ind. Microbiol. Biotechnol. 2009, 36, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New developments in Pichia pastoris expression system, review and update. Curr. Pharm. Biotechnol. 2018, 19, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Nwuche, C.O.; Nweze, J.E.; Ndubuisi, I.A.; Ogbonna, J.C. Potentials of multi-stress tolerant yeasts, Saccharomyces cerevisiae and Pichia kudriavzevii for fuel ethanol production from industrial cassava wastes. Process Biochem. 2021, 111, 305–314. [Google Scholar] [CrossRef]
- Greppi, A.; Saubade, F.; Botta, C.; Humblot, C.; Guyot, J.P.; Cocolin, L. Potential probiotic Pichia kudriavzevii strains and their ability to enhance folate content of traditional cereal-based African fermented food. Food Microbiol. 2017, 62, 169–177. [Google Scholar] [CrossRef]
- Choi, D.H.; Park, E.H.; Kim, M.D. Isolation of thermotolerant yeast Pichia kudriavzevii from nuruk. Food Sci. Biotechnol. 2017, 26, 1357–1362. [Google Scholar] [CrossRef]
- Johansen, P.G.; Owusu-Kwarteng, J.; Parkouda, C.; Padonou, S.W.; Jespersen, L. Occurrence and importance of yeasts in indigenous fermented food and beverages produced in sub-Saharan Africa. Front. Microbiol. 2019, 10, 1789. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Vila-Santa, A.; Liu, N.; Prozorov, T.; Xie, D.; Faria, N.T.; Ferreira, F.C.; Mira, N.P.; Shao, Z. Metabolic engineering of an acid-tolerant yeast strain Pichia kudriavzevii for itaconic acid production. Metab. Eng. Commun. 2020, 10, e00124. [Google Scholar] [CrossRef]
- Li, C.; Liu, Q.; Wang, Y.; Yang, X.; Chen, S.; Zhao, Y.; Wu, Y.; Li, L. Salt stress improves thermotolerance and high-temperature bioethanol production of multi-stress-tolerant Pichia kudriavzevii by stimulating intracellular metabolism and inhibiting oxidative damage. Biotechnol. Biofuels 2021, 14, 222. [Google Scholar] [CrossRef]
- Isono, N.; Hayakawa, H.; Usami, A.; Mishima, T.; Hisamatsu, M. A comparative study of ethanol production by Issatchenkia orientalis strains under stress conditions. J. Biosci. Bioeng. 2012, 113, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.F.; Gan, H.M.; Ling, H.L.; Rashid, N.A.A. Genome sequence of Pichia kudriavzevii M12, a potential producer of bioethanol and phytase. Eukaryot. Cell 2012, 11, 1300–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Zhan, T.; Xu, H.; Chen, J.; Bi, C.; Fan, F.; Zhang, X. Characterization of JEN family carboxylate transporters from the acid-tolerant yeast Pichia kudriavzevii and their applications in succinic acid production. Microb. Biotechnol. 2021, 14, 1130–1147. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef]
- Cavallo, E.; Charreau, H.; Cerrutti, P.; Foresti, M.L. Yarrowia lipolytica: A model yeast for citric acid production. FEMS Yeast Res. 2017, 17, fox084. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Chi, P.; Zou, Y.; Xu, Y.; Xu, S.; Bilal, M.; Fickers, P.; Cheng, H. Metabolic engineering of Yarrowia lipolytica for thermoresistance and enhanced erythritol productivity. Biotechnol. Biofuels 2020, 13, 176. [Google Scholar] [CrossRef]
- Muhammad, A.; Feng, X.; Rasool, A.; Sun, W.; Li, C. Production of plant natural products through engineered Yarrowia lipolytica. Biotechnol. Adv. 2020, 43, 107555. [Google Scholar] [CrossRef]
- Kim, O.C.; Suwannarangsee, S.; Oh, D.B.; Kim, S.; Seo, J.W.; Kim, C.H.; Kang, H.A.; Kim, J.Y.; Kwon, O. Transcriptome analysis of xylose metabolism in the thermotolerant methylotrophic yeast Hansenula polymorpha. Bioprocess Biosyst. Eng. 2013, 36, 1509–1518. [Google Scholar] [CrossRef]
- Choudhary, J.; Singh, S.; Nain, L. Thermotolerant fermenting yeasts for simultaneous saccharification fermentation of lignocellulosic biomass. Electron. J. Biotechnol. 2016, 21, 82–92. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Gombert, A.K.; Heinzle, E.; Wittmann, C. Physiology of the yeast Kluyveromyces marxianus during batch and chemostat cultures with glucose as the sole carbon source. FEMS Yeast Res. 2007, 7, 422–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banat, I.M.; Nigam, P.; Marchant, R. Isolation of thermotolerant, fermentative yeasts growing at 52 °C and producing ethanol at 45 °C and 50 °C. World J. Microbiol. Biotechnol. 1992, 8, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Cao, M.E.; Rico-Díaz, A.; Cerdán, M.E.; Becerra, M.; González-Siso, M.I. Valuation of agro-industrial wastes as substrates for heterologous production of α-galactosidase. Microb. Cell Fact. 2018, 17, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurcholis, M.; Lertwattanasakul, N.; Rodrussamee, N.; Kosaka, T.; Murata, M.; Yamada, M. Integration of comprehensive data and biotechnological tools for industrial applications of Kluyveromyces marxianus. Appl. Microbiol. Biotechnol. 2020, 104, 475–488. [Google Scholar] [CrossRef]
- Bilal, M.; Ji, L.; Xu, Y.; Xu, S.; Lin, Y.; Iqbal, H.M.N.; Cheng, H. Bioprospecting Kluyveromyces marxianus as a robust host for industrial biotechnology. Front. Bioeng. Biotechnol. 2022, 10, 851768. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Merino, R.A.; Varela, J.A.; Coughlan, A.Y.; Hoshida, H.; Da Silveira, W.B.; Wilde, C.; Kuijpers, N.G.A.; Geertman, J.M.; Wolfe, K.H.; Morrissey, J.P. Ploidy variation in Kluyveromyces marxianus separates dairy and non-dairy isolates. Front. Genet. 2018, 9, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, A.S.; Morrissey, J.P. Rational engineering of Kluyveromyces marxianus to create a chassis for the production of aromatic products. Microb. Cell Fact. 2020, 19, 207. [Google Scholar] [CrossRef]
- Cernak, P.; Estrela, R.; Poddar, S.; Skerker, J.M.; Cheng, Y.F.; Carlson, A.K.; Chen, B.; Glynn, V.M.; Furlan, M.; Ryan, O.W.; et al. Engineering Kluyveromyces marxianus as a robust synthetic biology platform host. mBio 2018, 9, e01410–e01418. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Han, Y.; Wang, X.; Zhao, X.; Chi, Z.; Chi, Z.; Liu, G. A new engineered endo-inulinase with improved activity and thermostability: Application in the production of prebiotic fructo-oligosaccharides from inulin. Food Chem. 2019, 294, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, C.; Wheeldon, I. Genome and metabolic engineering in non-conventional yeasts: Current advances and applications. Synth. Syst. Biotechnol. 2017, 2, 198–207. [Google Scholar]
- Da Silva, N.A.; Srikrishnan, S. Introduction and expression of genes for metabolic engineering applications in Saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 197–214. [Google Scholar] [CrossRef]
- Lee, M.E.; DeLoache, W.C.; Cervantes, B.; Dueber, J.E. A highly characterized yeast toolkit for modular, multipart assembly. ACS Synth. Biol. 2015, 4, 975–986. [Google Scholar] [CrossRef]
- Liu, L.; Otoupal, P.; Pan, A.; Alper, H.S. Increasing expression level and copy number of a Yarrowia lipolytica plasmid through regulated centromere function. FEMS Yeast Res. 2014, 14, 1124–1127. [Google Scholar]
- Camattari, A.; Goh, A.; Yip, L.Y.; Tan, A.H.M.; Ng, S.W.; Tran, A.; Liu, G.; Liachko, I.; Dunham, M.J.; Rancati, G. Characterization of a panARS-based episomal vector in the methylotrophic yeast Pichia pastoris for recombinant protein production and synthetic biology applications. Microb. Cell Fact. 2016, 15, 139. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Nishi, T.; Noguchi, R.; Ito, Y.; Watanabe, T.; Nishiyama, T.; Aikawa, S.; Hasunuma, T.; Ishii, J.; Okubo, Y. A stable, autonomously replicating plasmid vector containing Pichia pastoris centromeric DNA. Appl. Environ. Microbiol. 2018, 84, e02882. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lin, J.; Zhang, T.; Xu, K.; Ren, C.; Zhang, Z. Simultaneous screening and validation of effective zinc finger nucleases in yeast. PLoS ONE 2013, 8, e64687. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Lin, Y.; Guo, Y.; Qi, X.; Wang, Q. Metabolic and genomic characterisation of stress-tolerant industrial Saccharomyces cerevisiae strains from TALENs-assisted multiplex editing. FEMS Yeast Res. 2018, 18, foy045. [Google Scholar] [CrossRef]
- Song, A.J.; Palmiter, R.D. Detecting and avoiding problems when using the Cre–lox system. Trends Genet. 2018, 34, 333–340. [Google Scholar] [CrossRef]
- Mali, P.; Esvelt, K.M.; Church, G.M. Cas9 as a versatile tool for engineering biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.M.; Montoliu, L. On the origin of CRISPR-Cas technology: From prokaryotes to mammals. Trends Microbiol. 2016, 24, 811–820. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Dong, H.; Cui, Y.; Cong, L.; Zhang, D. Application of different types of CRISPR/Cas-based systems in bacteria. Microb. Cell Fact. 2020, 19, 172. [Google Scholar] [CrossRef]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Asmamaw, M.; Zawdie, B. Mechanism and applications of CRISPR/Cas-9-mediated genome editing. Biologics 2021, 15, 353–361. [Google Scholar]
- Palermo, G.; Chen, J.S.; Ricci, C.G.; Rivalta, I.; Jinek, M.; Batista, V.S.; Doudna, J.A.; McCammon, J.A. Key role of the REC lobe during CRISPR–Cas9 activation by ‘sensing’,‘regulating’, and ‘locking’ the catalytic HNH domain. Q. Rev. Biophys. 2018, 51, e91. [Google Scholar] [CrossRef] [Green Version]
- Ming, S.H.; Tian-Rui, X.U.; Ce-Shi, C.H. The big bang of genome editing technology: Development and application of the CRISPR/Cas9 system in disease animal models. Zool. Res. 2016, 37, 191–204. [Google Scholar]
- Daley, J.M.; Palmbos, P.L.; Wu, D.; Wilson, T.E. Nonhomologous end joining in yeast. Annu. Rev. Genet. 2005, 39, 431–451. [Google Scholar] [CrossRef]
- Brandsma, I.; Gent, D.C. Pathway choice in DNA double strand break repair: Observations of a balancing act. Genome Integr. 2012, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Qiu, Y.; Shi, S. CRISPR/Cas9 systems for the development of Saccharomyces cerevisiae cell factories. Front. Bioeng. Biotechnol. 2020, 8, 594347. [Google Scholar] [CrossRef]
- Schwartz, C.; Shabbir-Hussain, M.; Frogue, K.; Blenner, M.; Wheeldon, I. Standardized markerless gene integration for pathway engineering in Yarrowia lipolytica. ACS Synth. Biol. 2017, 6, 402–409. [Google Scholar] [CrossRef]
- de Souza, C.P.; Ribeiro, B.D.; Zarur Coelho, M.A.; Almeida, R.V.; Nicaud, J.M. Construction of wild-type Yarrowia lipolytica IMUFRJ 50682 auxotrophic mutants using dual CRISPR/Cas9 strategy for novel biotechnological approaches. Enzyme Microb. Technol. 2020, 140, 109621. [Google Scholar] [CrossRef]
- Zhang, X.K.; Wang, D.N.; Chen, J.; Liu, Z.J.; Wei, L.J.; Hua, Q. Metabolic engineering of β-carotene biosynthesis in Yarrowia lipolytica. Biotechnol. Lett. 2020, 42, 945–956. [Google Scholar] [CrossRef]
- Gao, S.; Tong, Y.; Zhu, L.; Ge, M.; Jiang, Y.; Chen, D.; Yang, S. Production of β-carotene by expressing a heterologous multifunctional carotene synthase in Yarrowia lipolytica. Biotechnol. Lett. 2017, 39, 921–927. [Google Scholar] [CrossRef]
- Wang, L.; Deng, A.; Zhang, Y.; Liu, S.; Liang, Y.; Bai, H.; Cui, D.; Qiu, Q.; Shang, X.; Yang, Z.; et al. Efficient CRISPR–Cas9 mediated multiplex genome editing in yeasts. Biotechnol. Biofuels 2018, 11, 277. [Google Scholar] [CrossRef]
- Juergens, H.; Varela, J.A.; Gorter de Vries, A.R.; Perli, T.; Gast, V.J.; Gyurchev, N.Y.; Rajkumar, A.S.; Mans, R.; Pronk, J.T.; Morrissey, J.P. Genome editing in Kluyveromyces and Ogataea yeasts using a broad-host-range Cas9/gRNA co-expression plasmid. FEMS Yeast Res. 2018, 18, foy012. [Google Scholar] [CrossRef]
- Siripong, W.; Angela, C.; Tanapongpipat, S.; Runguphan, W. Metabolic engineering of Pichia pastoris for production of isopentanol (3-methyl-1-butanol). Enzyme Microb. Technol. 2020, 138, 109557. [Google Scholar] [CrossRef]
- Liu, Q.; Shi, X.; Song, L.; Liu, H.; Zhou, X.; Wang, Q.; Zhang, Y.; Cai, M. CRISPR–Cas9-mediated genomic multiloci integration in Pichia pastoris. Microb. Cell Fact. 2019, 18, 144. [Google Scholar] [CrossRef] [Green Version]
- Gassler, T.; Heistinger, L.; Mattanovich, D.; Gasser, B.; Prielhofer, R. CRISPR/Cas9-mediated homology-directed genome editing in Pichia pastoris. In Recombinant Protein Production in Yeast; Springer: Berlin, Germany, 2019; pp. 211–225. [Google Scholar]
- Löbs, A.-K.; Engel, R.; Schwartz, C.; Flores, A.; Wheeldon, I. CRISPR–Cas9-enabled genetic disruptions for understanding ethanol and ethyl acetate biosynthesis in Kluyveromyces marxianus. Biotechnol. Biofuels 2017, 10, 164. [Google Scholar] [CrossRef]
- Tran, V.G.; Cao, M.; Fatma, Z.; Song, X.; Zhao, H. Development of a CRISPR/Cas9-based tool for gene deletion in Issatchenkia orientalis. mSphere 2019, 4, e00345-19. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.G.; Kim, C.; Kuanyshev, N.; Kang, N.K.; Fatma, Z.; Wu, Z.Y.; Cheng, M.H.; Singh, V.; Yoshikuni, Y.; Zhao, H.; et al. Cas9-based metabolic engineering of Issatchenkia orientalis for enhanced utilization of cellulosic hydrolysates. J. Agric. Food Chem. 2022, 70, 12085–12094. [Google Scholar] [CrossRef]
- Emerson, C.H.; Bertuch, A.A. Consider the workhorse: Nonhomologous end-joining in budding yeast. Biochem. Cell Biol. 2016, 94, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Matsumoto, Y.; Trujillo, K.M.; Lees-Miller, S.P.; Osley, M.A.; Tomkinson, A.E. Role of the yeast DNA repair protein Nej1 in end processing during the repair of DNA double strand breaks by non-homologous end joining. DNA Repair 2015, 31, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hefferin, M.L.; Chen, L.; Shim, E.Y.; Tseng, H.M.; Kwon, Y.; Sung, P.; Lee, S.E.; Tomkinson, A.E. Role of Dnl4–Lif1 in nonhomologous end-joining repair complex assembly and suppression of homologous recombination. Nat. Struct. Mol. Biol. 2007, 14, 639–646. [Google Scholar] [CrossRef]
- Utomo, J.C.; Hodgins, C.L.; Ro, D.K. Multiplex Genome Editing in Yeast by CRISPR/Cas9–A Potent and Agile Tool to Reconstruct Complex Metabolic Pathways. Front. Plant Sci. 2021, 12, 719148. [Google Scholar] [CrossRef]
- Yan, Y.; Finnigan, G.C. Development of a multi-locus CRISPR gene drive system in budding yeast. Sci. Rep. 2018, 8, 17277. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, A.S.; Varela, J.A.; Juergens, H.; Daran, J.M.G.; Morrissey, J.P. Biological parts for Kluyveromyces marxianus synthetic biology. Front. Bioeng. Biotechnol. 2019, 7, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Näätsaari, L.; Mistlberger, B.; Ruth, C.; Hajek, T.; Hartner, F.S.; Glieder, A. Deletion of the Pichia pastoris KU70 homologue facilitates platform strain generation for gene expression and synthetic biology. PLoS ONE 2012, 7, e39720. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Duan, X.; Cai, P.; Gao, L.; Wu, X.; Yao, L.; Zhou, Y.J. Fusing an exonuclease with Cas9 enhances homologous recombination in Pichia pastoris. Microb. Cell Fact. 2022, 21, 182. [Google Scholar] [CrossRef]
- Paffett, K.S.; Clikeman, J.A.; Palmer, S.; Nickoloff, J.A. Overexpression of Rad51 inhibits double-strand break-induced homologous recombination but does not affect gene conversion tract lengths. DNA Repair 2005, 4, 687–698. [Google Scholar] [CrossRef]
- Cai, P.; Duan, X.; Wu, X.; Gao, L.; Ye, M.; Zhou, Y.J. Recombination machinery engineering facilitates metabolic engineering of the industrial yeast Pichia pastoris. Nucleic Acids Res. 2021, 49, 7791–7805. [Google Scholar] [CrossRef]
- Ji, Q.; Mai, J.; Ding, Y.; Wei, Y.; Ledesma-Amaro, R.; Ji, X.-J. Improving the homologous recombination efficiency of Yarrowia lipolytica by grafting heterologous component from Saccharomyces cerevisiae. Metab. Eng. Commun. 2020, 11, e00152. [Google Scholar] [CrossRef]
- Weninger, A.; Hatzl, A.M.; Schmid, C.; Vogl, T.; Glieder, A. Combinatorial optimization of CRISPR/Cas9 expression enables precision genome engineering in the methylotrophic yeast Pichia pastoris. J. Biotechnol. 2016, 235, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhao, Y. Self-processing of ribozyme-flanked RNAs into guide RNAs in vitro and in vivo for CRISPR-mediated genome editing. J. Integr. Plant Biol. 2014, 56, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Ryan, O.W.; Skerker, J.M.; Maurer, M.J.; Li, X.; Tsai, J.C.; Poddar, S.; Lee, M.E.; DeLoache, W.; Dueber, J.E.; Arkin, A.P.; et al. Selection of chromosomal DNA libraries using a multiplex CRISPR system. eLife 2014, 3, e03703. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinstiver, B.P.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Topkar, V.V.; Zheng, Z.; Joung, J.K. Broadening the targeting range of Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition. Nat. Biotechnol. 2015, 33, 1293–1298. [Google Scholar] [CrossRef] [Green Version]
- Rees, H.A.; Yeh, W.-H.; Liu, D.R. Development of hRad51–Cas9 nickase fusions that mediate HDR without double-stranded breaks. Nat. Commun. 2019, 10, 2212. [Google Scholar] [CrossRef] [Green Version]
- Schusterbauer, V.; Fischer, J.E.; Gangl, S.; Schenzle, L.; Rinnofner, C.; Geier, M.; Sailer, C.; Glieder, A.; Thallinger, G.G. Whole Genome Sequencing Analysis of Effects of CRISPR/Cas9 in Komagataella phaffii: A Budding Yeast in Distress. J. Fungi 2022, 8, 992. [Google Scholar] [CrossRef]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.G.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Kawser Hossain, M.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Kumar Saha, S.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Molecular mechanisms of the anti-obesity and anti-diabetic properties of flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Sáez, J.; Wang, G.; Marella, E.R.; Sudarsan, S.; Cernuda Pastor, M.; Borodina, I. Engineering the oleaginous yeast Yarrowia lipolytica for high-level resveratrol production. Metab. Eng. 2020, 62, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Pyne, M.E.; Narcross, L.; Martin, V.J.J. Engineering plant secondary metabolism in microbial systems. Plant Physiol. 2019, 179, 844–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, J.; Wu, X.; Gong, G.; Koffas, M.A.G. Pathway enzyme engineering for flavonoid production in recombinant microbes. Metab. Eng. Commun. 2019, 9, e00104. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Sun, X.; Yan, Y.; Yuan, Q.; Wang, J.; Shen, X. Metabolic engineering of microorganisms for the production of flavonoids. Front. Bioeng. Biotechnol. 2020, 8, 589069. [Google Scholar] [CrossRef]
- Soreanu, I.; Hendler, A.; Dahan, D.; Dovrat, D.; Aharoni, A. Marker-free genetic manipulations in yeast using CRISPR/CAS9 system. Curr. Genet. 2018, 64, 1129–1139. [Google Scholar] [CrossRef]
- Lv, Y.; Marsafari, M.; Koffas, M.; Zhou, J.; Xu, P. Optimizing oleaginous yeast cell factories for flavonoids and hydroxylated flavonoids biosynthesis. ACS Synth. Biol. 2019, 8, 2514–2523. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhang, P.; Shang, Y.; Zhou, Y.; Ye, B.C. Metabolically engineering of Yarrowia lipolytica for the biosynthesis of naringenin from a mixture of glucose and xylose. Bioresour. Technol. 2020, 314, 123726. [Google Scholar] [CrossRef]
- Toti, E.; Chen, C.Y.O.; Palmery, M.; Villaño Valencia, D.; Peluso, I. Non-provitamin A and provitamin A carotenoids as immunomodulators: Recommended dietary allowance, therapeutic index, or personalized nutrition? Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnesen, J.A.; Kildegaard, K.R.; Cernuda Pastor, M.; Jayachandran, S.; Kristensen, M.; Borodina, I. Yarrowia lipolytica strains engineered for the production of terpenoids. Front. Bioeng. Biotechnol. 2020, 8, 945. [Google Scholar] [CrossRef]
- Lu, Y.; Yang, Q.; Lin, Z.; Yang, X. A modular pathway engineering strategy for the high-level production of β-ionone in Yarrowia lipolytica. Microb. Cell Fact. 2020, 19, 49. [Google Scholar] [CrossRef]
- Yao, F.; Liu, S.C.; Wang, D.N.; Liu, Z.J.; Hua, Q.; Wei, L.J. Engineering oleaginous yeast Yarrowia lipolytica for enhanced limonene production from xylose and lignocellulosic hydrolysate. FEMS Yeast Res. 2020, 20, foaa046. [Google Scholar] [CrossRef]
- Zhang, J.L.; Bai, Q.Y.; Peng, Y.Z.; Fan, J.; Jin, C.C.; Cao, Y.X.; Yuan, Y.J. High production of triterpenoids in Yarrowia lipolytica through manipulation of lipid components. Biotechnol. Biofuels 2020, 13, 133. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lu, X.; Zong, H.; Li, J.; Zhuge, B. Itaconic acid production in microorganisms. Biotechnol. Lett. 2018, 40, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cui, Z.; Zhao, X.; Zhang, J.; Zhang, L.; Tian, Y.; Qi, Q.; Liu, J. Enhanced itaconic acid production in Yarrowia lipolytica via heterologous expression of a mitochondrial transporter MTT. Appl. Microbiol. Biotechnol. 2019, 103, 2181–2192. [Google Scholar] [CrossRef]
- Blazeck, J.; Miller, J.; Pan, A.; Gengler, J.; Holden, C.; Jamoussi, M.; Alper, H.S. Metabolic engineering of Saccharomyces cerevisiae for itaconic acid production. Appl. Microbiol. Biotechnol. 2014, 98, 8155–8164. [Google Scholar] [CrossRef] [PubMed]
- Yook, S.D.; Kim, J.; Gong, G.; Ko, J.K.; Um, Y.; Han, S.O.; Lee, S.M. High-yield lipid production from lignocellulosic biomass using engineered xylose-utilizing Yarrowia lipolytica. Glob. Change Biol. Bioenergy 2020, 12, 670–679. [Google Scholar] [CrossRef]
- Li, J.X.; Xu, J.; Ruan, J.C.; Meng, H.M.; Su, H.; Han, X.F.; Lu, M.; Li, F.L.; Wang, S.A. Disrupting a phospholipase A2 gene increasing lipid accumulation in the oleaginous yeast Yarrowia lipolytica. J. Appl. Microbiol. 2021, 130, 100–108. [Google Scholar] [CrossRef]
- Zhang, J.L.; Cao, Y.X.; Peng, Y.Z.; Jin, C.C.; Bai, Q.Y.; Zhang, R.S.; Liu, D.; Yuan, Y.J. High production of fatty alcohols in Yarrowia lipolytica by coordination with glycolysis. Sci. China Chem. 2019, 62, 1007–1016. [Google Scholar] [CrossRef]
- Reyes-Sánchez, F.J.; Páez-Lerma, J.B.; Rojas-Contreras, J.A.; López-Miranda, J.; Soto-Cruz, N.Ó.; Reinhart-Kirchmayr, M. Study of the enzymatic capacity of Kluyveromyces marxianus for the synthesis of esters. J. Mol. Microbiol. Biotechnol. 2019, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lang, X.; Moran Cabrera, M.; De Keyser, S.; Sun, X.; Da Silva, N.; Wheeldon, I. CRISPR-mediated multigene integration enables Shikimate pathway refactoring for enhanced 2-phenylethanol biosynthesis in Kluyveromyces marxianus. Biotechnol. Biofuels 2021, 14, 3. [Google Scholar] [CrossRef]
- Löbs, A.K.; Schwartz, C.; Thorwall, S.; Wheeldon, I. Highly multiplexed CRISPRi repression of respiratory functions enhances mitochondrial localized ethyl acetate biosynthesis in Kluyveromyces marxianus. ACS Synth. Biol. 2018, 7, 2647–2655. [Google Scholar] [CrossRef] [PubMed]
- Swietalski, P.; Hetzel, F.; Seitl, I.; Fischer, L. Secretion of a low and high molecular weight β-glycosidase by Yarrowia lipolytica. Microb. Cell Fact. 2020, 19, 100. [Google Scholar] [CrossRef] [PubMed]
- Bruder, S.; Moldenhauer, E.J.; Lemke, R.D.; Ledesma-Amaro, R.; Kabisch, J. Drop-in biofuel production using fatty acid photodecarboxylase from Chlorella variabilis in the oleaginous yeast Yarrowia lipolytica. Biotechnol. Biofuels 2019, 12, 202. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Kim, I.J.; Kim, S.R. Nonconventional Yeasts Engineered Using the CRISPR-Cas System as Emerging Microbial Cell Factories. Fermentation 2022, 8, 656. https://doi.org/10.3390/fermentation8110656
Park J, Kim IJ, Kim SR. Nonconventional Yeasts Engineered Using the CRISPR-Cas System as Emerging Microbial Cell Factories. Fermentation. 2022; 8(11):656. https://doi.org/10.3390/fermentation8110656
Chicago/Turabian StylePark, Jongbeom, In Jung Kim, and Soo Rin Kim. 2022. "Nonconventional Yeasts Engineered Using the CRISPR-Cas System as Emerging Microbial Cell Factories" Fermentation 8, no. 11: 656. https://doi.org/10.3390/fermentation8110656
APA StylePark, J., Kim, I. J., & Kim, S. R. (2022). Nonconventional Yeasts Engineered Using the CRISPR-Cas System as Emerging Microbial Cell Factories. Fermentation, 8(11), 656. https://doi.org/10.3390/fermentation8110656