
Identification of Mutations Responsible for Improved Xylose Utilization in an Adapted Xylose Isomerase Expressing Saccharomyces cerevisiae Strain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and General Methods
2.2. Growth Analysis
2.3. Genome Sequencing and Analysis
2.4. Mating-Type Testing and MAT Switching
2.5. Tetrad Dissection
2.6. SNP PCR
2.7. Batch Fermentation
3. Results and Discussion
3.1. Genome Sequence Analysis of the Adapted Strain
3.2. Determination of Phenotype Dominance
3.3. Tetrad Dissection to Identify Causative Mutations
3.4. Validation of PBS2 and PHO13 Genes
3.5. Analysis of Strains with PBS2 and PHO13 Deletions
3.6. Analysis of PBS2 and PHO13 Requirement for Strains Expressing the XR/XDH Pathway
3.7. Xylose Fermentation in Strains with Targeted PBS2 and PHO13 Deletions
3.8. Mutation to HOG1 Pathway Kinases Improves Xylose Utilization
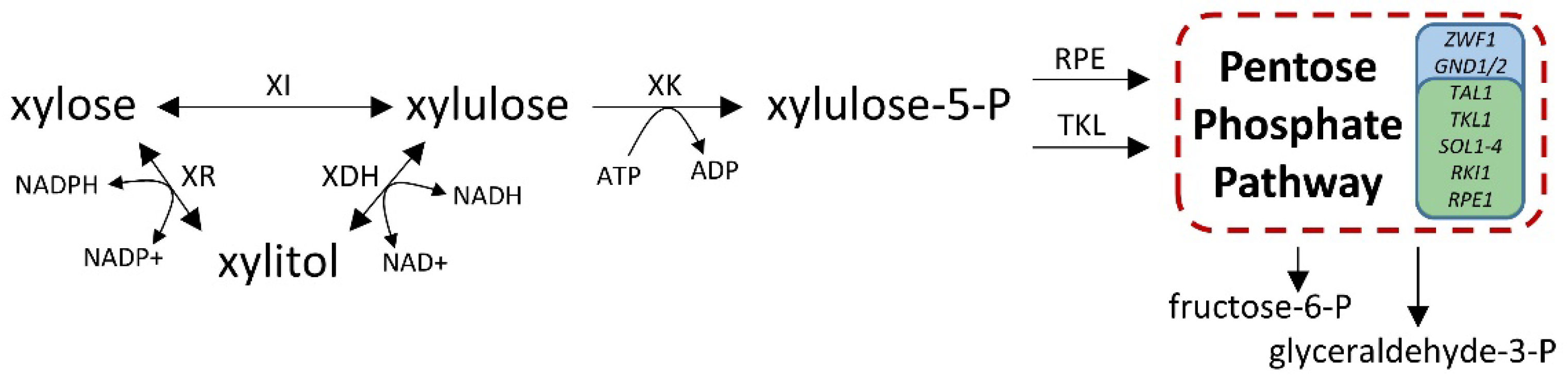
3.9. Model Showing Involvement of PBS2 and PHO13 in Separate Steps of Xylose Utilization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevoigt, E. Progress in metabolic engineering of Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2008, 72, 379–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, M.L.A.; Bracher, J.M.; Papapetridis, I.; Verhoeven, M.D.; de Bruijn, H.; de Waal, P.P.; van Maris, A.J.A.; Klaassen, P.; Pronk, J.T. Saccharomyces cerevisiae strains for second-generation ethanol production: From academic exploration to industrial implementation. FEMS Yeast Res. 2017, 17, fox044. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Yook, S.; Koh, H.; Rao, C.V.; Jin, Y.S. Engineering xylose metabolism in yeasts to produce biofuels and chemicals. Curr. Opin. Biotechnol. 2021, 67, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D.B.; Bowman, M.J.; Braker, J.D.; Dien, B.S.; Hector, R.E.; Lee, C.C.; Mertens, J.A.; Wagschal, K. Plant cell walls to ethanol. Biochem. J. 2012, 442, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Borgstrom, C.; Wasserstrom, L.; Almqvist, H.; Broberg, K.; Klein, B.; Noack, S.; Liden, G.; Gorwa-Grauslund, M.F. Identification of modifications procuring growth on xylose in recombinant Saccharomyces cerevisiae strains carrying the Weimberg pathway. Metab. Eng. 2019, 55, 1–11. [Google Scholar] [CrossRef]
- Kotter, P.; Ciriacy, M. Xylose Fermentation by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1993, 38, 776–783. [Google Scholar] [CrossRef]
- Bruinenberg, P.M.; de Bot, P.H.M.; van Dijken, J.P.; Scheffers, W.A. The role of redox balances in the anaerobic fermentation of xylose by yeasts. Eur. J. Appl. Microbiol. Biotechnol. 1983, 18, 287–292. [Google Scholar] [CrossRef]
- van Maris, A.J.; Winkler, A.A.; Kuyper, M.; de Laat, W.T.; van Dijken, J.P.; Pronk, J.T. Development of efficient xylose fermentation in Saccharomyces cerevisiae: Xylose isomerase as a key component. Adv. Biochem. Eng. Biotechnol. 2007, 108, 179–204. [Google Scholar] [CrossRef] [Green Version]
- Harhangi, H.R.; Akhmanova, A.S.; Emmens, R.; van der Drift, C.; de Laat, W.T.; van Dijken, J.P.; Jetten, M.S.; Pronk, J.T.; Op den Camp, H.J. Xylose metabolism in the anaerobic fungus Piromyces sp. strain E2 follows the bacterial pathway. Arch. Microbiol. 2003, 180, 134–141. [Google Scholar] [CrossRef]
- Gardonyi, M.; Hahn-Hagerdal, B. The Streptomyces rubiginosus xylose isomerase is misfolded when expressed in Saccharomyces cerevisiae. Enzym. Microb. Technol. 2003, 32, 252–259. [Google Scholar] [CrossRef]
- Kuyper, M.; Hartog, M.M.; Toirkens, M.J.; Almering, M.J.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Metabolic engineering of a xylose-isomerase-expressing Saccharomyces cerevisiae strain for rapid anaerobic xylose fermentation. FEMS Yeast Res. 2005, 5, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brat, D.; Boles, E.; Wiedemann, B. Functional expression of a bacterial xylose isomerase in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 2304–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhavan, A.; Tamalampudi, S.; Ushida, K.; Kanai, D.; Katahira, S.; Srivastava, A.; Fukuda, H.; Bisaria, V.S.; Kondo, A. Xylose isomerase from polycentric fungus Orpinomyces: Gene sequencing, cloning, and expression in Saccharomyces cerevisiae for bioconversion of xylose to ethanol. Appl. Microbiol. Biotechnol. 2009, 82, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Cheng, J.S.; Wang, B.L.; Fink, G.R.; Stephanopoulos, G. Xylose isomerase overexpression along with engineering of the pentose phosphate pathway and evolutionary engineering enable rapid xylose utilization and ethanol production by Saccharomyces cerevisiae. Metab. Eng. 2012, 14, 611–622. [Google Scholar] [CrossRef]
- Bamba, T.; Hasunuma, T.; Kondo, A. Disruption of PHO13 improves ethanol production via the xylose isomerase pathway. AMB Express 2016, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Hector, R.E.; Dien, B.S.; Cotta, M.A.; Mertens, J.A. Growth and fermentation of D-xylose by Saccharomyces cerevisiae expressing a novel D-xylose isomerase originating from the bacterium Prevotella ruminicola TC2-24. Biotechnol. Biofuels 2013, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.K.; Tremaine, M.; Parreiras, L.S.; Hebert, A.S.; Myers, K.S.; Higbee, A.J.; Sardi, M.; McIlwain, S.J.; Ong, I.M.; Breuer, R.J.; et al. Directed evolution reveals unexpected epistatic interactions that alter metabolic regulation and enable anaerobic xylose use by Saccharomyces cerevisiae. PLoS Genet. 2016, 12, e1006372. [Google Scholar] [CrossRef] [Green Version]
- Kuyper, M.; Toirkens, M.J.; Diderich, J.A.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Evolutionary engineering of mixed-sugar utilization by a xylose-fermenting Saccharomyces cerevisiae strain. FEMS Yeast Res. 2005, 5, 925–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia Sanchez, R.; Karhumaa, K.; Fonseca, C.; Sanchez Nogue, V.; Almeida, J.R.; Larsson, C.U.; Bengtsson, O.; Bettiga, M.; Hahn-Hägerdal, B.; Gorwa-Grauslund, M.F. Improved xylose and arabinose utilization by an industrial recombinant Saccharomyces cerevisiae strain using evolutionary engineering. Biotechnol. Biofuels 2010, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-M.; Jellison, T.; Alper, H.S. Systematic and evolutionary engineering of a xylose isomerase-based pathway in Saccharomyces cerevisiae for efficient conversion yields. Biotechnol. Biofuels 2014, 7, 122. [Google Scholar] [CrossRef]
- Tomas-Pejo, E.; Ballesteros, M.; Oliva, J.M.; Olsson, L. Adaptation of the xylose fermenting yeast Saccharomyces cerevisiae F12 for improving ethanol production in different fed-batch SSF processes. J. Ind. Microbiol. Biotechnol. 2010, 37, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, X.; Peng, B.; Chen, L.; Hou, J.; Bao, X. An efficient xylose-fermenting recombinant Saccharomyces cerevisiae strain obtained through adaptive evolution and its global transcription profile. Appl. Microbiol. Biotechnol. 2012, 96, 1079–1091. [Google Scholar] [CrossRef]
- Kuyper, M.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Minimal metabolic engineering of Saccharomyces cerevisiae for efficient anaerobic xylose fermentation: A proof of principle. FEMS Yeast Res. 2004, 4, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Shen, Y.; Li, X.; Chen, X.; Hou, J.; Bao, X. Improvement of xylose fermentation in respiratory-deficient xylose-fermenting Saccharomyces cerevisiae. Metab. Eng. 2012, 14, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Sauer, U. Evolutionary engineering of Saccharomyces cerevisiae for anaerobic growth on xylose. Appl. Environ. Microbiol. 2003, 69, 1990–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisselink, H.W.; Toirkens, M.J.; Wu, Q.; Pronk, J.T.; van Maris, A.J. Novel evolutionary engineering approach for accelerated utilization of glucose, xylose, and arabinose mixtures by engineered Saccharomyces cerevisiae strains. Appl. Environ. Microbiol. 2009, 75, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.; van Rensburg, E.; Gorgens, J.F. Simultaneously improving xylose fermentation and tolerance to lignocellulosic inhibitors through evolutionary engineering of recombinant Saccharomyces cerevisiae harbouring xylose isomerase. BMC Biotechnol. 2014, 14, 41. [Google Scholar] [CrossRef] [Green Version]
- Scalcinati, G.; Otero, J.M.; Van Vleet, J.R.; Jeffries, T.W.; Olsson, L.; Nielsen, J. Evolutionary engineering of Saccharomyces cerevisiae for efficient aerobic xylose consumption. FEMS Yeast Res. 2012, 12, 582–597. [Google Scholar] [CrossRef] [Green Version]
- Demeke, M.M.; Dietz, H.; Li, Y.; Foulquie-Moreno, M.R.; Mutturi, S.; Deprez, S.; Den Abt, T.; Bonini, B.M.; Liden, G.; Dumortier, F.; et al. Development of a D-xylose fermenting and inhibitor tolerant industrial Saccharomyces cerevisiae strain with high performance in lignocellulose hydrolysates using metabolic and evolutionary engineering. Biotechnol. Biofuels 2013, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Matsushika, A.; Oguri, E.; Sawayama, S. Evolutionary adaptation of recombinant shochu yeast for improved xylose utilization. J. Biosci. Bioeng. 2010, 110, 102–105. [Google Scholar] [CrossRef]
- Kim, S.R.; Xu, H.; Lesmana, A.; Kuzmanovic, U.; Au, M.; Florencia, C.; Oh, E.J.; Zhang, G.; Kim, K.H.; Jin, Y.S. Deletion of PHO13, encoding haloacid dehalogenase type IIA phosphatase, results in upregulation of the pentose phosphate pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 1601–1609. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.; Laplaza, J.M.; Jeffries, T.W. Transposon mutagenesis to improve the growth of recombinant Saccharomyces cerevisiae on D-xylose. Appl. Environ. Microbiol. 2007, 73, 2061–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.R.; Skerker, J.M.; Kang, W.; Lesmana, A.; Wei, N.; Arkin, A.P.; Jin, Y.S. Rational and Evolutionary Engineering Approaches Uncover a Small Set of Genetic Changes Efficient for Rapid Xylose Fermentation in Saccharomyces cerevisiae. PLoS ONE 2013, 8, e57048. [Google Scholar] [CrossRef] [Green Version]
- Parreiras, L.S.; Breuer, R.J.; Avanasi Narasimhan, R.; Higbee, A.J.; La Reau, A.; Tremaine, M.; Qin, L.; Willis, L.B.; Bice, B.D.; Bonfert, B.L.; et al. Engineering and two-stage evolution of a lignocellulosic hydrolysate-tolerant Saccharomyces cerevisiae strain for anaerobic fermentation of xylose from AFEX pretreated corn stover. PLoS ONE 2014, 9, e107499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, L.V.; Carazzolle, M.F.; Nagamatsu, S.T.; Sampaio, N.M.V.; Almeida, L.D.; Pirolla, R.A.S.; Borelli, G.; Corrêa, T.L.R.; Argueso, J.L.; Pereira, G.A.G. Unraveling the genetic basis of xylose consumption in engineered Saccharomyces cerevisiae strains. Sci. Rep. 2016, 6, 38676. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Tremaine, M.; Place, M.; Liu, L.; Pier, A.; Krause, D.J.; Xie, D.; Zhang, Y.; Landick, R.; Gasch, A.P.; et al. Crabtree/Warburg-like aerobic xylose fermentation by engineered Saccharomyces cerevisiae. Metab. Eng. 2021, 68, 119–130. [Google Scholar] [CrossRef]
- Hou, J.; Jiao, C.; Peng, B.; Shen, Y.; Bao, X. Mutation of a regulator Ask10p improves xylose isomerase activity through up-regulation of molecular chaperones in Saccharomyces cerevisiae. Metab. Eng. 2016, 38, 241–250. [Google Scholar] [CrossRef]
- Papapetridis, I.; Verhoeven, M.D.; Wiersma, S.J.; Goudriaan, M.; van Maris, A.J.A.; Pronk, J.T. Laboratory evolution for forced glucose-xylose co-consumption enables identification of mutations that improve mixed-sugar fermentation by xylose-fermenting Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, foy056. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, M.D.; Lee, M.; Kamoen, L.; van den Broek, M.; Janssen, D.B.; Daran, J.-M.G.; van Maris, A.J.A.; Pronk, J.T. Mutations in PMR1 stimulate xylose isomerase activity and anaerobic growth on xylose of engineered Saccharomyces cerevisiae by influencing manganese homeostasis. Sci. Rep. 2017, 7, 46155. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.; Oh, E.J.; Ko, J.K.; Nam, J.O.; Park, H.S.; Jin, Y.S.; Lee, E.J.; Kim, S.R. Metabolic engineering considerations for the heterologous expression of xylose-catabolic pathways in Saccharomyces cerevisiae. PLoS ONE 2020, 15, e0236294. [Google Scholar] [CrossRef]
- Demeke, M.M.; Foulquie-Moreno, M.R.; Dumortier, F.; Thevelein, J.M. Rapid evolution of recombinant Saccharomyces cerevisiae for Xylose fermentation through formation of extra-chromosomal circular DNA. PLoS Genet. 2015, 11, e1005010. [Google Scholar] [CrossRef] [Green Version]
- Hector, R.E.; Mertens, J.A.; Nichols, N.N. Development and characterization of vectors for tunable expression of both xylose-regulated and constitutive gene expression in Saccharomyces yeasts. New Biotechnol. 2019, 53, 16–23. [Google Scholar] [CrossRef]
- Christianson, T.W.; Sikorski, R.S.; Dante, M.; Shero, J.H.; Hieter, P. Multifunctional yeast high-copy-number shuttle vectors. Gene 1992, 110, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.B.; Lewis, A.L.; Baldwin, K.K.; Resnick, M.A. Lethality induced by a single site-specific double-strand break in a dispensable yeast plasmid. Proc. Natl. Acad. Sci. USA 1993, 90, 5613–5617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hector, R.E.; Dien, B.S.; Cotta, M.A.; Qureshi, N. Engineering industrial Saccharomyces cerevisiae strains for xylose fermentation and comparison for switchgrass conversion. J. Ind. Microbiol. Biotechnol. 2011, 38, 1193–1202. [Google Scholar] [CrossRef]
- Hector, R.E.; Mertens, J.A.; Bowman, M.J.; Nichols, N.N.; Cotta, M.A.; Hughes, S.R. Saccharomyces cerevisiae engineered for xylose metabolism requires gluconeogenesis and the oxidative branch of the pentose phosphate pathway for aerobic xylose assimilation. Yeast 2011, 28, 645–660. [Google Scholar] [CrossRef]
- Hauf, J.; Zimmermann, F.K.; Muller, S. Simultaneous genomic overexpression of seven glycolytic enzymes in the yeast Saccharomyces cerevisiae. Enzym. Microb. Technol. 2000, 26, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.A.; Kelly, A.; Hector, R.E. Screening for inhibitor tolerant Saccharomyces cerevisiae strains from diverse environments for use as platform strains for production of fuels and chemicals from biomass. Bioresour. Technol. Rep. 2018, 3, 154–161. [Google Scholar] [CrossRef]
- Huxley, C.; Green, E.D.; Dunham, I. Rapid assessment of S. cerevisiae mating type by PCR. Trends Genet. 1990, 6, 236. [Google Scholar] [CrossRef]
- Chen, J.; Schedl, T. A simple one-step PCR assay for SNP detection. MicroPubl. Biol. 2021, 1–4. [Google Scholar] [CrossRef]
- Nichols, N.N.; Hector, R.E.; Saha, B.C.; Frazer, S.E.; Kennedy, G.J. Biological abatement of inhibitors in rice hull hydrolyzate and fermentation to ethanol using conventional and engineered microbes. Biomass Bioenergy 2014, 67, 79–88. [Google Scholar] [CrossRef]
- Araya, C.L.; Payen, C.; Dunham, M.J.; Fields, S. Whole-genome sequencing of a laboratory-evolved yeast strain. BMC Genom. 2010, 11, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, R.M.; Rash, B.M.; Hentges, K.E.; Robertson, D.L.; Delneri, D.; Lovell, S.C. Gene duplication and environmental adaptation within yeast populations. Genome Biol. Evol. 2010, 2, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.S.; Ni, H.; Laplaza, J.M.; Jeffries, T.W. Optimal growth and ethanol production from xylose by recombinant Saccharomyces cerevisiae require moderate D-xylulokinase activity. Appl. Environ. Microbiol. 2003, 69, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Johansson, B.; Christensson, C.; Hobley, T.; Hahn-Hagerdal, B. Xylulokinase overexpression in two strains of Saccharomyces cerevisiae also expressing xylose reductase and xylitol dehydrogenase and its effect on fermentation of xylose and lignocellulosic hydrolysate. Appl. Environ. Microbiol. 2001, 67, 4249–4255. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Peña, J.M.; Cid, V.J.; Arroyo, J.; Nombela, C. The YGR194c (XKS1) gene encodes the xylulokinase from the budding yeast Saccharomyces cerevisiae. FEMS Microbiol. Lett. 1998, 162, 155–160. [Google Scholar] [CrossRef]
- Deng, X.X.; Ho, N.W. Xylulokinase activity in various yeasts including Saccharomyces cerevisiae containing the cloned xylulokinase gene. Scientific note. Appl. Biochem. Biotechnol. 1990, 24, 193–199. [Google Scholar] [CrossRef]
- Matsushika, A.; Sawayama, S. Comparative study on a series of recombinant flocculent Saccharomyces cerevisiae strains with different expression levels of xylose reductase and xylulokinase. Enzym. Microb. Technol. 2011, 48, 466–471. [Google Scholar] [CrossRef]
- Krieger, F.; Moglich, A.; Kiefhaber, T. Effect of proline and glycine residues on dynamics and barriers of loop formation in polypeptide chains. J. Am. Chem. Soc. 2005, 127, 3346–3352. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Tatebayashi, K.; Saito, H. Two adjacent docking sites in the yeast Hog1 mitogen-activated protein (MAP) kinase differentially interact with the Pbs2 MAP kinase kinase and the Ptp2 protein tyrosine phosphatase. Mol. Cell. Biol. 2008, 28, 2481–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Posas, F. Response to hyperosmotic stress. Genetics 2012, 192, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, S.M.; Herskowitz, I. Unique and redundant roles for HOG MAPK pathway components as revealed by whole-genome expression analysis. Mol. Biol. Cell 2004, 15, 532–542. [Google Scholar] [CrossRef]
- Aguilera, J.; Rodriguez-Vargas, S.; Prieto, J.A. The HOG MAP kinase pathway is required for the induction of methylglyoxal-responsive genes and determines methylglyoxal resistance in Saccharomyces cerevisiae. Mol. Microbiol. 2005, 56, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Garay-Arroyo, A.; Covarrubias, A.A. Three genes whose expression is induced by stress in Saccharomyces cerevisiae. Yeast 1999, 15, 879–892. [Google Scholar] [CrossRef]
- Träff, K.L.; Otero Cordero, R.R.; van Zyl, W.H.; Hahn-Hägerdal, B. Deletion of the GRE3 aldose reductase gene and its influence on xylose metabolism in recombinant strains of Saccharomyces cerevisiae expressing the xylA and XKS1 genes. Appl. Environ. Microbiol. 2001, 67, 5668–5674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalevsky, A.; Hanson, B.L.; Mason, S.A.; Forsyth, V.T.; Fisher, Z.; Mustyakimov, M.; Blakeley, M.P.; Keen, D.A.; Langan, P. Inhibition of D-xylose isomerase by polyols: Atomic details by joint X-ray/neutron crystallography. Acta Crystallogr. D 2012, 68, 1201–1206. [Google Scholar] [CrossRef]
- Van Vleet, J.H.; Jeffries, T.W.; Olsson, L. Deleting the para-nitrophenyl phosphatase (pNPPase), PHO13, in recombinant Saccharomyces cerevisiae improves growth and ethanol production on D-xylose. Metab. Eng. 2008, 10, 360–369. [Google Scholar] [CrossRef]
- Gutmann, F.; Jann, C.; Pereira, F.; Johansson, A.; Steinmetz, L.M.; Patil, K.R. CRISPRi screens reveal genes modulating yeast growth in lignocellulose hydrolysate. Biotechnol. Biofuels 2021, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.R.; Myers, K.S.; Riley, N.M.; Coon, J.J.; Gasch, A.P. PKA and HOG signaling contribute separable roles to anaerobic xylose fermentation in yeast engineered for biofuel production. PLoS ONE 2019, 14, e0212389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maayan, I.; Beenstock, J.; Marbach, I.; Tabachnick, S.; Livnah, O.; Engelberg, D. Osmostress induces autophosphorylation of Hog1 via a C-terminal regulatory region that is conserved in p38alpha. PLoS ONE 2012, 7, e44749. [Google Scholar] [CrossRef] [PubMed]

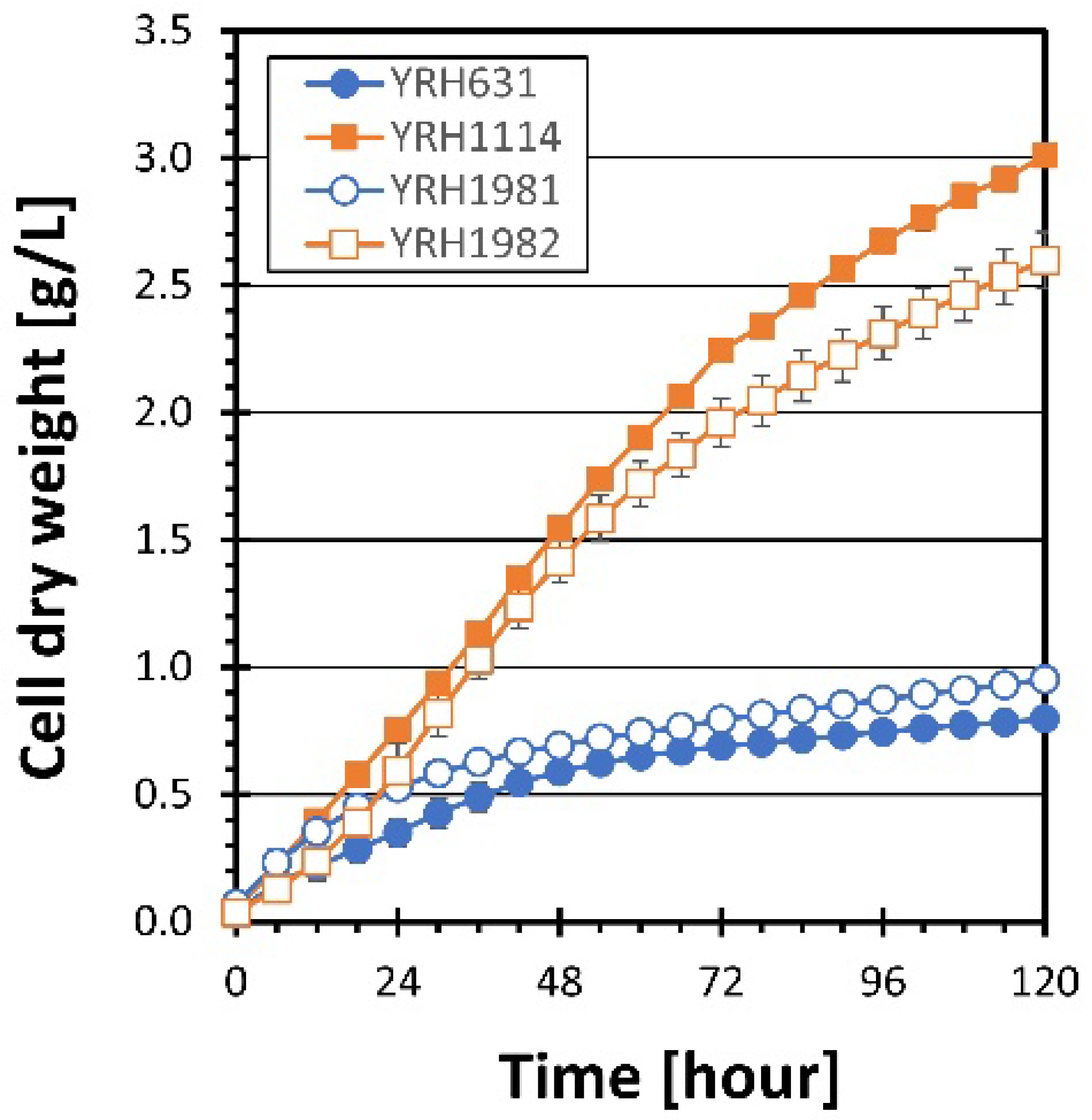
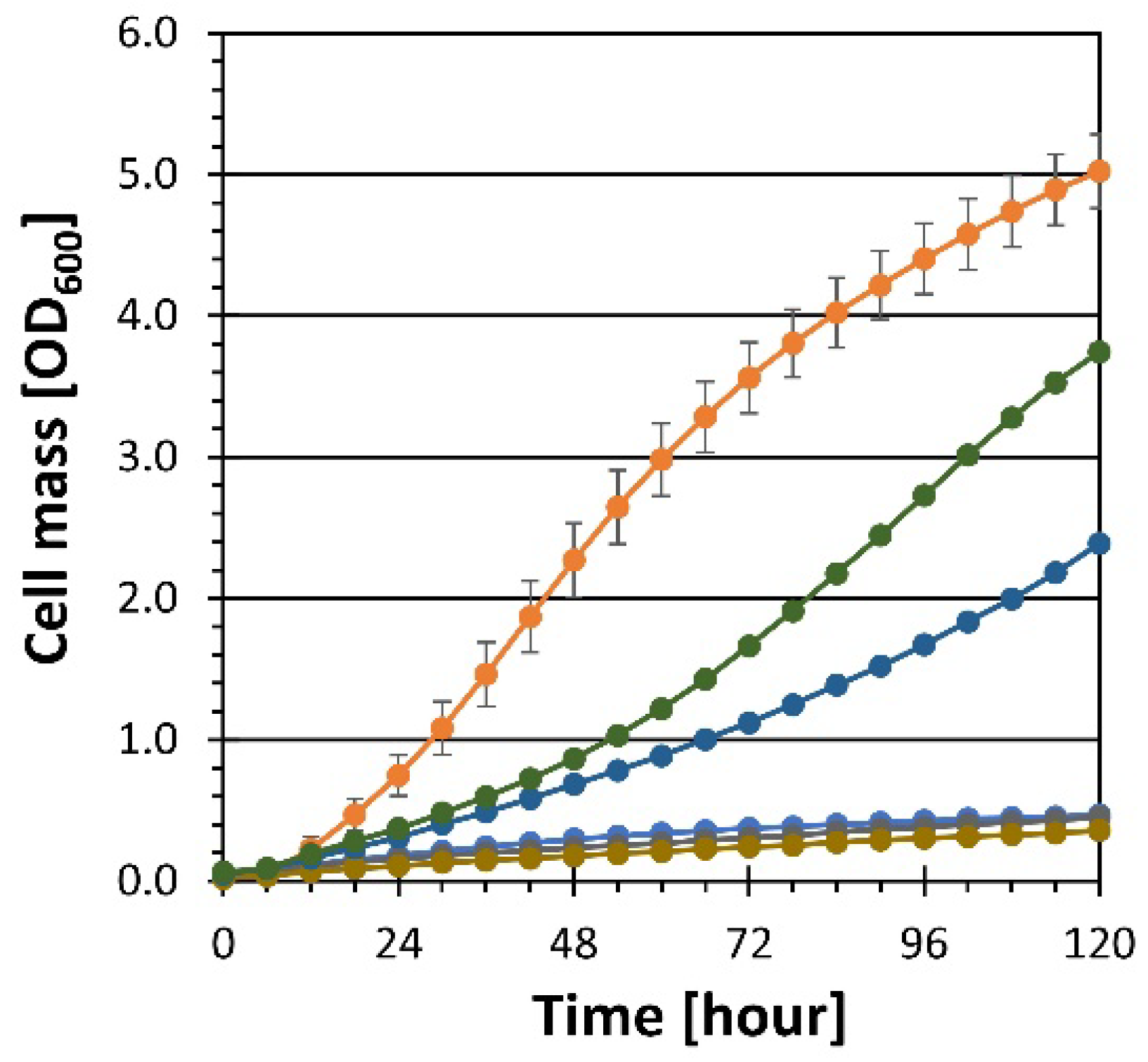
 ) and heterozygous diploid (YRH1981
) and heterozygous diploid (YRH1981  ) strains were grown on YP medium + 50 g/L xylose and compared to the unadapted haploid (YRH631
) strains were grown on YP medium + 50 g/L xylose and compared to the unadapted haploid (YRH631  ) and adapted haploid (YRH1114
) and adapted haploid (YRH1114  ) strains. Cell dry weight was used to measure cell mass instead of OD600 to account for differences in dry weight per OD600 between haploid and diploid strains. Plots are the average values from at least triplicate cultures. Error bars show standard deviations.
) and heterozygous diploid (YRH1981 ) strains were grown on YP medium + 50 g/L xylose and compared to the unadapted haploid (YRH631 ) and adapted haploid (YRH1114 ) strains. Cell dry weight was used to measure cell mass instead of OD600 to account for differences in dry weight per OD600 between haploid and diploid strains. Plots are the average values from at least triplicate cultures. Error bars show standard deviations.
) strains. Cell dry weight was used to measure cell mass instead of OD600 to account for differences in dry weight per OD600 between haploid and diploid strains. Plots are the average values from at least triplicate cultures. Error bars show standard deviations.
) and heterozygous diploid (YRH1981 ) strains were grown on YP medium + 50 g/L xylose and compared to the unadapted haploid (YRH631 ) and adapted haploid (YRH1114 ) strains. Cell dry weight was used to measure cell mass instead of OD600 to account for differences in dry weight per OD600 between haploid and diploid strains. Plots are the average values from at least triplicate cultures. Error bars show standard deviations.

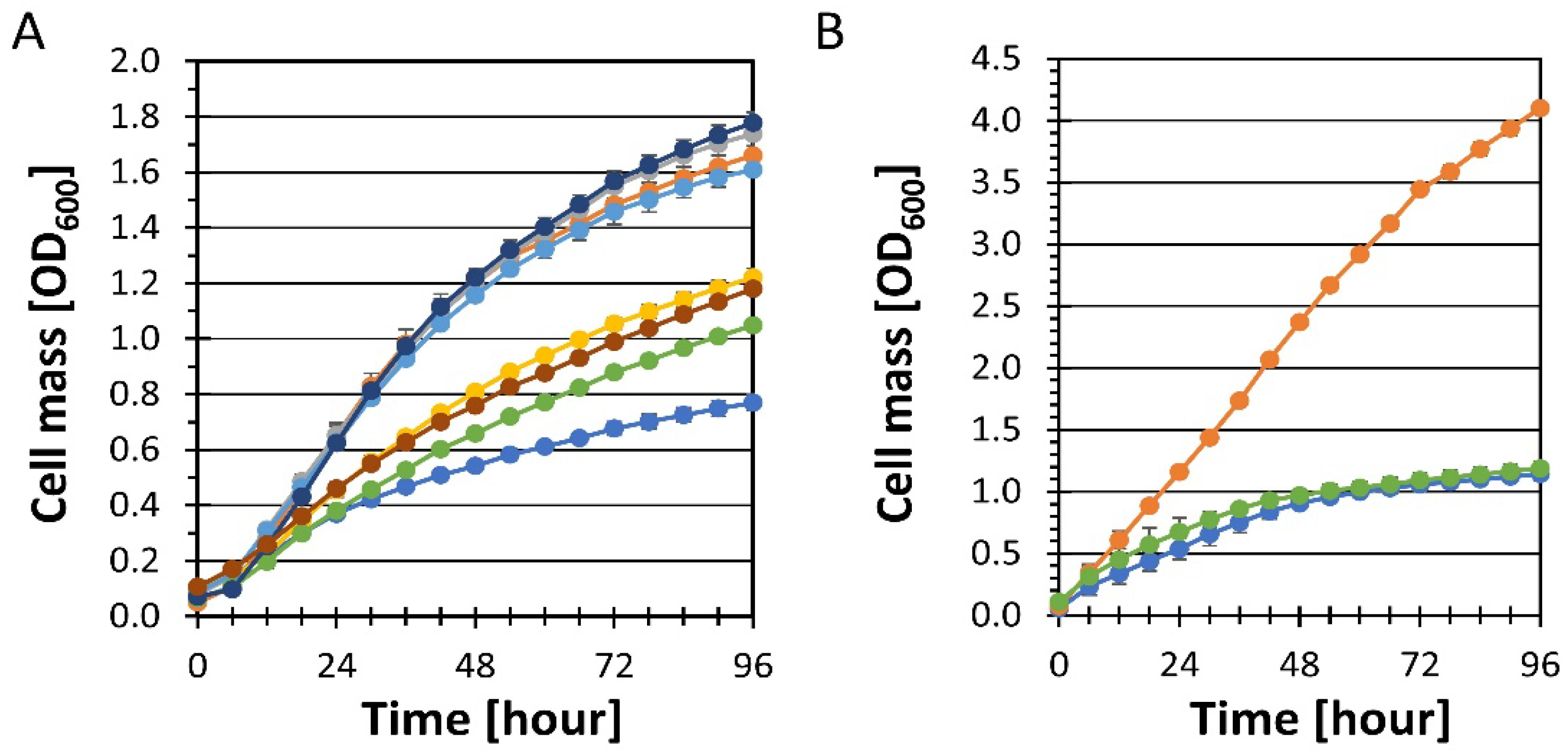
 YRH631 (unadapted strain),
YRH631 (unadapted strain),  YRH1114 (adapted strain); Panel (A)
YRH1114 (adapted strain); Panel (A)  YRH1966 (YRH1114 + empty vector),
YRH1966 (YRH1114 + empty vector),  YRH1968 (YRH1114 + PBS2 plasmid),
YRH1968 (YRH1114 + PBS2 plasmid),  YRH2042 (YRH1114 + empty vector),
YRH2042 (YRH1114 + empty vector),  YRH2043 (YRH1114 + PHO13 plasmid),
YRH2043 (YRH1114 + PHO13 plasmid),  YRH2044 (YRH1114 + empty vectors),
YRH2044 (YRH1114 + empty vectors),  YRH2045 (YRH1114 + PBS2 and PHO13 plasmids); Panel (B)
YRH2045 (YRH1114 + PBS2 and PHO13 plasmids); Panel (B)  YRH1934 (YRH1114 + corrected genomic PBS2).
YRH631 (unadapted strain), YRH1114 (adapted strain); Panel (A) YRH1966 (YRH1114 + empty vector), YRH1968 (YRH1114 + PBS2 plasmid), YRH2042 (YRH1114 + empty vector), YRH2043 (YRH1114 + PHO13 plasmid), YRH2044 (YRH1114 + empty vectors), YRH2045 (YRH1114 + PBS2 and PHO13 plasmids); Panel (B) YRH1934 (YRH1114 + corrected genomic PBS2).
YRH1934 (YRH1114 + corrected genomic PBS2).
YRH631 (unadapted strain), YRH1114 (adapted strain); Panel (A) YRH1966 (YRH1114 + empty vector), YRH1968 (YRH1114 + PBS2 plasmid), YRH2042 (YRH1114 + empty vector), YRH2043 (YRH1114 + PHO13 plasmid), YRH2044 (YRH1114 + empty vectors), YRH2045 (YRH1114 + PBS2 and PHO13 plasmids); Panel (B) YRH1934 (YRH1114 + corrected genomic PBS2).
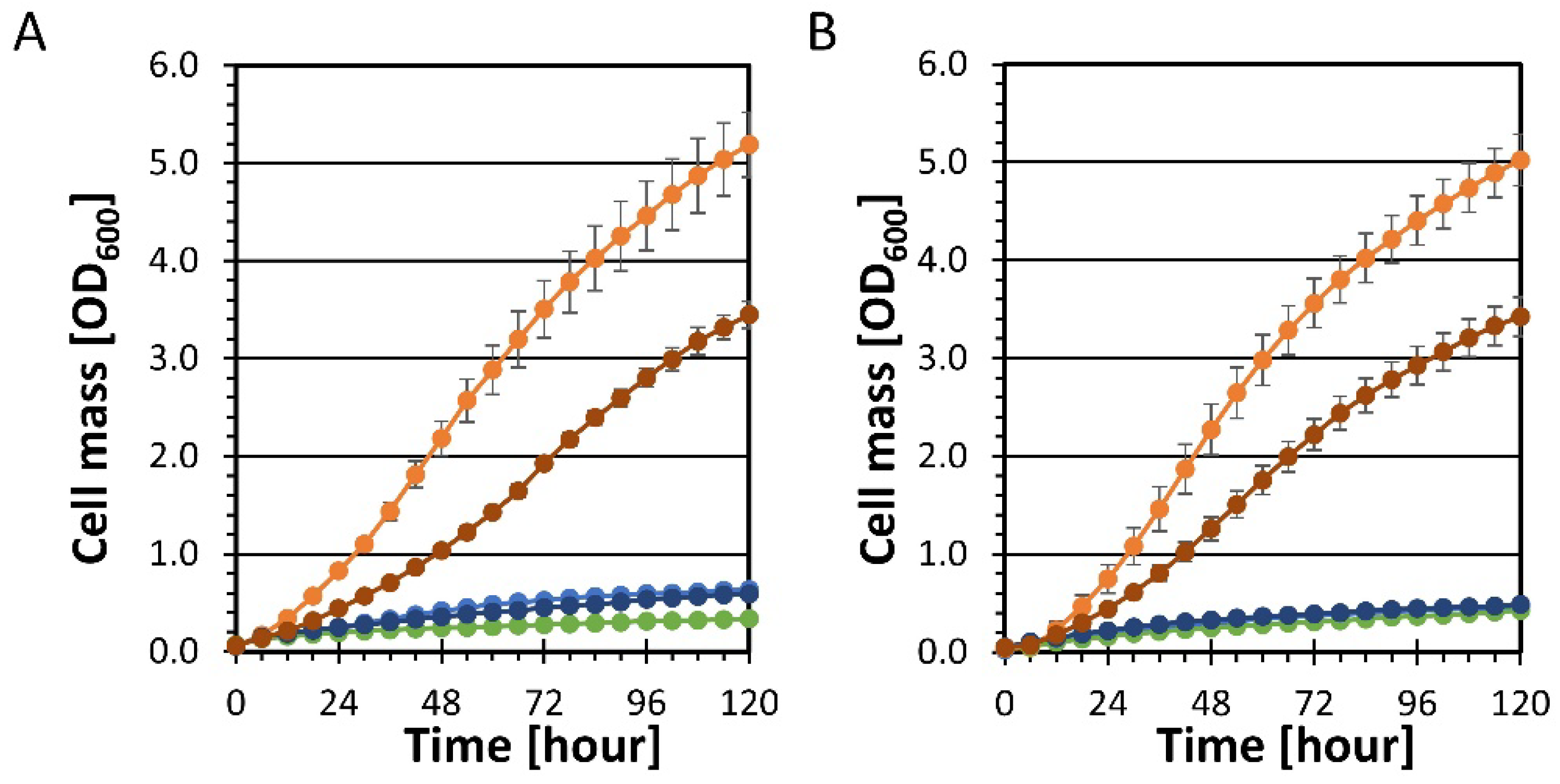
 YRH631 (unadapted strain),
YRH631 (unadapted strain),  YRH1114 (adapted strain); Panel (A),
YRH1114 (adapted strain); Panel (A),  YRH1563 (YRH631 + pho13Δ),
YRH1563 (YRH631 + pho13Δ),  YRH2021 (YRH631 + pbs2Δ),
YRH2021 (YRH631 + pbs2Δ),  YRH2022 (YRH631 + pbs2Δ, pho13Δ); Panel (B),
YRH2022 (YRH631 + pbs2Δ, pho13Δ); Panel (B),  YRH2056 (CEN.PK2-1C + pbs2Δ),
YRH2056 (CEN.PK2-1C + pbs2Δ),  YRH2057 (CEN.PK2-1C + pho13Δ),
YRH2057 (CEN.PK2-1C + pho13Δ),  YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ).
YRH631 (unadapted strain), YRH1114 (adapted strain); Panel (A), YRH1563 (YRH631 + pho13Δ), YRH2021 (YRH631 + pbs2Δ), YRH2022 (YRH631 + pbs2Δ, pho13Δ); Panel (B), YRH2056 (CEN.PK2-1C + pbs2Δ), YRH2057 (CEN.PK2-1C + pho13Δ), YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ).
YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ).
YRH631 (unadapted strain), YRH1114 (adapted strain); Panel (A), YRH1563 (YRH631 + pho13Δ), YRH2021 (YRH631 + pbs2Δ), YRH2022 (YRH631 + pbs2Δ, pho13Δ); Panel (B), YRH2056 (CEN.PK2-1C + pbs2Δ), YRH2057 (CEN.PK2-1C + pho13Δ), YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ).
 YRH1114 (adapted XI strain),
YRH1114 (adapted XI strain),  YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid),
YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid),  YRH2053 (YRH2040 + pbs2Δ),
YRH2053 (YRH2040 + pbs2Δ),  YRH2054 (YRH2040 + pho13Δ),
YRH2054 (YRH2040 + pho13Δ),  YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH1114 (adapted XI strain), YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid), YRH2053 (YRH2040 + pbs2Δ), YRH2054 (YRH2040 + pho13Δ), YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH1114 (adapted XI strain), YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid), YRH2053 (YRH2040 + pbs2Δ), YRH2054 (YRH2040 + pho13Δ), YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
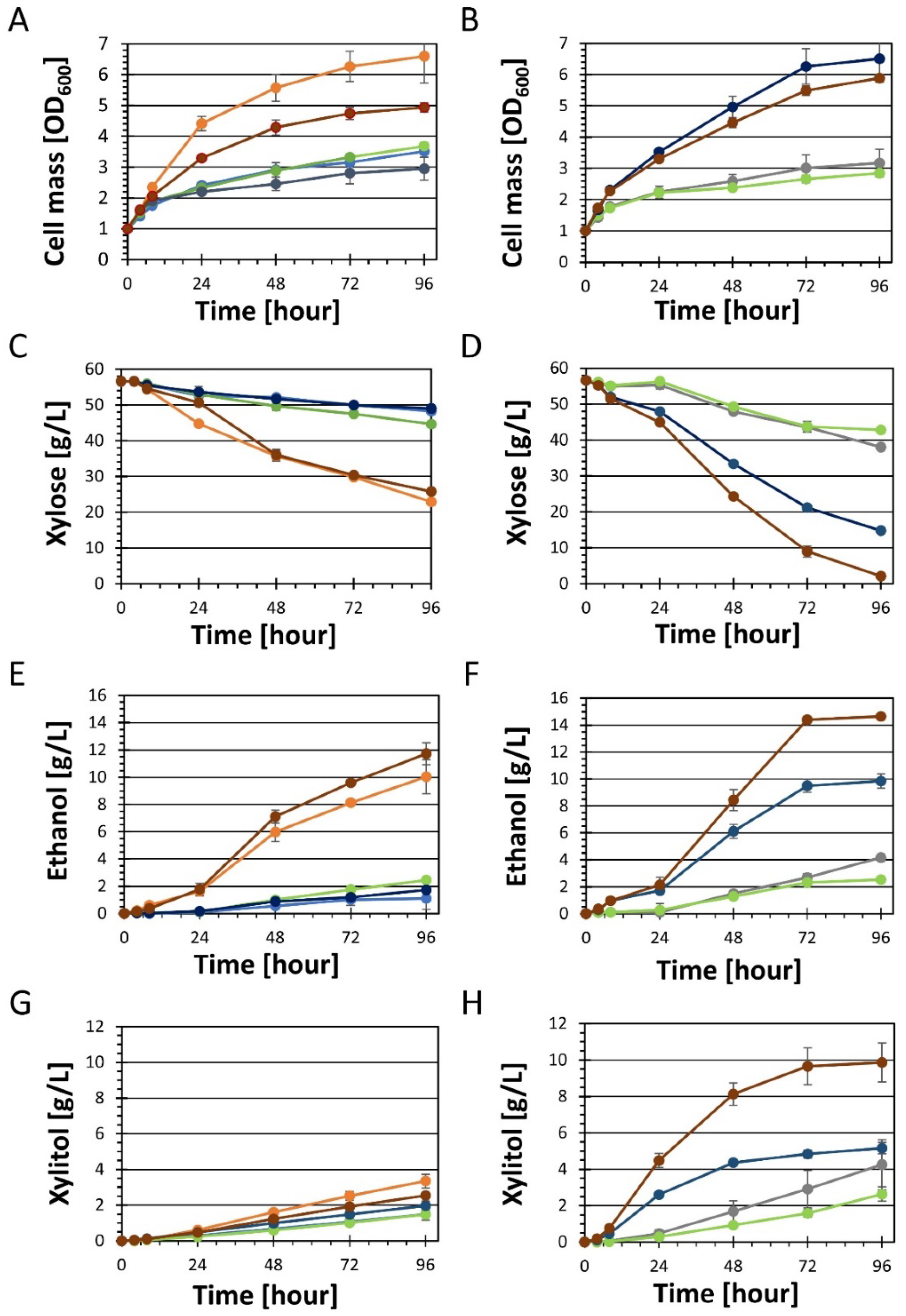
 YRH631 (unadapted XI strain),
YRH631 (unadapted XI strain),  YRH1114 (adapted XI strain),
YRH1114 (adapted XI strain),  YRH2056 (CEN.PK2-1C + pbs2Δ),
YRH2056 (CEN.PK2-1C + pbs2Δ),  YRH2057 (CEN.PK2-1C + pho13Δ),
YRH2057 (CEN.PK2-1C + pho13Δ),  YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ); Panels (B,D,F,H),
YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ); Panels (B,D,F,H),  YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid),
YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid),  YRH2053 (YRH2040 + pbs2Δ),
YRH2053 (YRH2040 + pbs2Δ),  YRH2054 (YRH2040 + pho13Δ),
YRH2054 (YRH2040 + pho13Δ),  YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH631 (unadapted XI strain), YRH1114 (adapted XI strain), YRH2056 (CEN.PK2-1C + pbs2Δ), YRH2057 (CEN.PK2-1C + pho13Δ), YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ); Panels (B,D,F,H), YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid), YRH2053 (YRH2040 + pbs2Δ), YRH2054 (YRH2040 + pho13Δ), YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH2055 (YRH2040 + pbs2Δ, pho13Δ).
YRH631 (unadapted XI strain), YRH1114 (adapted XI strain), YRH2056 (CEN.PK2-1C + pbs2Δ), YRH2057 (CEN.PK2-1C + pho13Δ), YRH2058 (CEN.PK2-1C + pbs2Δ, pho13Δ); Panels (B,D,F,H), YRH2040 (CEN.PK2-1C with XR/XDH/XK plasmid), YRH2053 (YRH2040 + pbs2Δ), YRH2054 (YRH2040 + pho13Δ), YRH2055 (YRH2040 + pbs2Δ, pho13Δ).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid # | Description | Reference |
|---|---|---|
| pRS413 | pBluescript II SK+, HIS3, CEN6, ARSH4 | [43] |
| pRS414 | pBluescript II SK+, TRP1, CEN6, ARSH4 | [43] |
| pRS415 | pBluescript II SK+, LEU2, CEN6, ARSH4 | [43] |
| pRS416 | pBluescript II SK+, URA3, CEN6, ARSH4 | [43] |
| pRS426 | pBluescript II SK+, URA3, 2μ origin | [43] |
| pGALHOT | PGAL10HO, TRP1, CEN4 | [44] |
| pRH164 A | pRS414 + PHXT7—MCS—THXT7 | [45] |
| pRH167 A | pRS426 + PHXT7—MCS—THXT7 | [45] |
| pRH274 B | pRS416 + PPGK1-SsXYL1-TPGK1; PADH1-SsXYL2-TADH1; PHXT7-ScXKS1-THXT7 | [46] |
| pRH384 | pRH167 + Xylose Isomerase C cloned into the MCS | [16] |
| pRH385 | pRH164 + Xylulokinase C cloned into the MCS | [16] |
| pRH544 | Xylulokinase C plasmid pRH385 rescued from YRH1137 | This study |
| pRH545 | Xylose isomerase C plasmid pRH384 rescued from YRH1138 | This study |
| pRH1000 | pRS415 + PPBS2-PBS2-TPBS2 | This study |
| pRH1011 | pRS413 + PPHO13-PHO13-TPHO13 | This study |
| Strain # | Genotype/Description | Reference |
|---|---|---|
| NEB 5-α | E. coli fhuA2Δ(argF-lacZ)U169 phoA glnV44 f80Δ(lacZ)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17 | NEB |
| CEN.PK2-1C | S. cerevisiae MATa, ura3-52, trp1-289, leu2-3,112, his3Δ 1, MAL2-8c; SUC2 | Euroscarf |
| YRH631 | CEN.PK2-1C + [pRH384, pRH385] [also carries ste24L418F] | [16] |
| YRH1114 | YRH631 microaerobically adapted on xylose medium | [16] |
| YRH1136 | YRH1114 without pRH384 and pRH385 | [16] |
| YRH1137 | YRH1114 with only pRH385 (xylulokinase plasmid) | This study |
| YRH1138 | YRH1114 with only pRH384 (xylose isomerase plasmid) | This study |
| YRH1169 | YRH1136 + [pRH544, pRH545] | This study |
| YRH1170 | YRH1136 + [pRH384, pRH385] | This study |
| YRH1563 | YRH631 with pho13Δ::HIS3 | This study |
| YRH1188 | CEN.PK2-1C + [pRH384, pRH385], remade to correct STE24 | This study |
| YRH1911 | YRH1136 + [pGAL-HOT] (HO expression vector for switching mating type) | This study |
| YRH1932 | YRH1114 with fixed genomic PBS2 | This study |
| YRH1934 | YRH1114 with fixed genomic PBS2 + [pRH384, pRH385] | This study |
| YRH1954 | YRH1136 with mating type switched to MATα | This study |
| YRH1955 | YRH1954 + [pRS413] (empty HIS3 vector for diploid selection) | This study |
| YRH1966 | YRH1114 + [pRS415] (empty LEU2 vector control) | This study |
| YRH1968 | YRH1114 + [pRH1000] (PBS2 expression vector) | This study |
| YRH1981 | YRH1955 X YRH631 (heterozygous diploid): MATa/MATα, PBS2/pbs2, SAS3/sas3, PHO13/pho13, PHO81/pho81, HSP104/hsp104, ste24/ste24 + [pRH384, pRH385] | This study |
| YRH1982 | YRH1955 X YRH1114 (homozygous diploid): MATa/MATα, pbs2/pbs2, sas3/sas3, pho13/pho13, pho81/pho81, hsp104/hsp104, ste24/ste24 + [pRH384, pRH385] | This study |
| YRH1994 | YRH1981 tetrad #1 spore A | This study |
| YRH1995 | YRH1981 tetrad #1 spore B | This study |
| YRH1996 | YRH1981 tetrad #1 spore C | This study |
| YRH1997 | YRH1981 tetrad #1 spore D | This study |
| YRH2002 | YRH1981 tetrad #3 spore A | This study |
| YRH2003 | YRH1981 tetrad #3 spore B | This study |
| YRH2004 | YRH1981 tetrad #3 spore C | This study |
| YRH2005 | YRH1981 tetrad #3 spore D | This study |
| YRH2006 | YRH1981 tetrad #4 spore A | This study |
| YRH2007 | YRH1981 tetrad #4 spore B | This study |
| YRH2008 | YRH1981 tetrad #4 spore C | This study |
| YRH2009 | YRH1981 tetrad #4 spore D | This study |
| YRH2010 | YRH1981 tetrad #5 spore A | This study |
| YRH2011 | YRH1981 tetrad #5 spore B | This study |
| YRH2012 | YRH1981 tetrad #5 spore C | This study |
| YRH2013 | YRH1981 tetrad #5 spore D | This study |
| YRH2014 | YRH1981 tetrad #6 spore A | This study |
| YRH2015 | YRH1981 tetrad #6 spore B | This study |
| YRH2016 | YRH1981 tetrad #6 spore C | This study |
| YRH2017 | YRH1981 tetrad #6 spore D | This study |
| YRH2021 | YRH631 with pbs2Δ::LEU2 | This study |
| YRH2022 | YRH631 with pbs2Δ::LEU2, pho13Δ::HIS3 | This study |
| YRH2040 | CEN.PK2-1C + [pRH274] (XR/XDH/XK plasmid) | This study |
| YRH2042 | YRH1114 + [pRS413] (empty HIS3 vector control) | This study |
| YRH2043 | YRH1114 + [pRH1011] (PHO13 expression vector) | This study |
| YRH2044 | YRH1114 + [pRS413, pRS415] (empty HIS3, LEU2 vector controls) | This study |
| YRH2045 | YRH1114 + [pRH1000, pRH1011] (PHO13 and PBS2 expression vectors) | This study |
| YRH2053 | CEN.PK2-1C with pbs2Δ::LEU2 + [pRH274] (XR/XDH/XK plasmid) | This study |
| YRH2054 | CEN.PK2-1C with pho13Δ::HIS3 + [pRH274] (XR/XDH/XK plasmid) | This study |
| YRH2055 | CEN.PK2-1C with pbs2Δ::LEU2, pho13Δ::HIS3 + [pRH274] (XR/XDH/XK plasmid) | This study |
| YRH2056 | CEN.PK2-1C with pbs2Δ::LEU2 + [pRH384, pRH385] | This study |
| YRH2057 | CEN.PK2-1C with pho13Δ::HIS3 + [pRH384, pRH385] | This study |
| YRH2058 | CEN.PK2-1C with pbs2Δ::LEU2, pho13Δ::HIS3 + [pRH384, pRH385] | This study |
| GENE | SNP | ORF | Chromosome | Function * |
|---|---|---|---|---|
| SAS3 | G733 to T | E245X | 2-L | Histone acetyltransferase subunit of NuA3 complex that catalyzes acetylation of histone H3 |
| YBR225W | C1839 to T | N631(Silent) | 2-R | Protein whose biological role and cellular location are unknown |
| PHO13 | G622 to T | G208C | 4-L | Alkaline phosphatase specific for p-nitrophenyl phosphate |
| PHO81 | C3182 to T | P1061L | 7-R | Cyclin-dependent protein serine/threonine kinase inhibitor involved in phosphate metabolism |
| PBS2 | T1008 to A | L363X | 10-L | MAP kinase kinase of the HOG signaling pathway; activated under severe osmotic stress |
| STE24 | C1252 to T | L418F | 10-R | Highly conserved zinc metalloprotease; component of the ER quality control mechanism that removes faulty proteins clogging translocation channels |
| HSP104 | T1892 to C | L631S | 12-L | Adenosine-binding protein chaperone involved in protein folding |
| Strain | Tetrad | MAT | Growth * | PBS2 ** | PHO13 | SAS3 | HSP104 | PHO81 | STE24 |
|---|---|---|---|---|---|---|---|---|---|
| YRH 1994 | 1A | α | +++ | − | − | + | − | + | − |
| YRH 1995 | 1B | a | − | + | + | − | + | − | − |
| YRH 1996 | 1C | α | +++ | − | − | + | − | + | − |
| YRH 1997 | 1D | a | − | + | + | − | + | + | − |
| YRH 2002 | 3A | a | − | + | − | + | + | − | − |
| YRH 2003 | 3B | α | − | + | + | − | + | + | − |
| YRH 2004 | 3C | a | + | − | + | − | − | − | − |
| YRH 2005 | 3D | α | +++ | − | − | + | − | + | − |
| YRH 2006 | 4A | a | + | + | + | + | − | − | − |
| YRH 2007 | 4B | a | + | − | + | − | − | + | − |
| YRH 2008 | 4C | α | + | + | − | − | + | + | − |
| YRH 2009 | 4D | α | +++ | − | − | + | + | − | − |
| YRH 2010 | 5A | α | +++ | − | − | + | + | + | − |
| YRH 2011 | 5B | α | ++ | + | − | − | + | − | − |
| YRH 2012 | 5C | a | + | + | + | + | − | − | − |
| YRH 2013 | 5D | a | + | − | + | − | − | − | − |
| YRH 2014 | 6A | a | − | − | + | − | + | + | − |
| YRH 2015 | 6B | α | ++ | + | − | − | − | − | − |
| YRH 2016 | 6C | a | − | + | − | + | + | + | − |
| YRH 2017 | 6D | α | + | − | + | + | − | − | − |
| Strain | Genotype | Xylose Pathway Utilized | Xylose Consumed [g/L] | Final Concentration [g/L] | Specific Consumption or Production Rate [g/gCDW/h] | Ethanol Yield [gethanol/gxylose] | ||
|---|---|---|---|---|---|---|---|---|
| Xylitol | Ethanol | Xylose | Ethanol | |||||
| YRH631 | PHO13, PBS2 | XI/XK A | 8.3 ± 0.81 | 1.5 ± 0.33 | 2.3 ± 0.81 | 0.04 ± 0.003 | 0.012 ± 0.0066 | 0.18 ± 0.053 |
| YRH1114 | pho13G208C, pbs2L363X | XI/XK | 33.7 ± 0.60 | 3.4 ± 0.38 | 10.0 ± 1.25 | 0.08 ± 0.011 | 0.025 ± 0.0064 | 0.30 ± 0.036 |
| YRH2056 | PHO13, pbs2Δ | XI/XK | 12.0 ± 0.15 | 1.5 ± 0.04 | 2.5 ± 0.07 | 0.05 ± 0.001 | 0.011 ± 0.0004 | 0.20 ± 0.007 |
| YRH2057 | pho13Δ, PBS2 | XI/XK | 7.6 ± 0.18 | 2.0 ± 0.25 | 1.7 ± 0.19 | 0.04 ± 0.005 | 0.007 ± 0.0051 | 0.23 ± 0.018 |
| YRH2058 | pho13Δ, pbs2Δ | XI/XK | 30.8 ± 0.57 | 2.5 ± 0.01 | 11.7 ± 0.81 | 0.10 ± 0.013 | 0.033 ± 0.0075 | 0.36 ± 0.036 |
| YRH2040 | PHO13, PBS2 | XR/XDH/XK B | 18.6 ± 0.30 | 4.3 ± 1.37 | 4.2 ± 0.27 | 0.09 ± 0.012 | 0.021 ± 0.0030 | 0.22 ± 0.011 |
| YRH2053 | PHO13, pbs2Δ | XR/XDH/XK | 13.8 ± 0.54 | 2.6 ± 0.39 | 2.5 ± 0.16 | 0.08 ± 0.004 | 0.014 ± 0.0010 | 0.18 ± 0.005 |
| YRH2054 | pho13Δ, PBS2 | XR/XDH/XK | 41.8 ± 0.48 | 5.2 ± 0.31 | 9.8 ± 0.52 | 0.10 ± 0.013 | 0.025 ± 0.0040 | 0.24 ± 0.013 |
| YRH2055 | pho13Δ, pbs2Δ | XR/XDH/XK | 54.5 ± 0.62 | 9.9 ± 1.06 | 14.7 ± 0.56 | 0.15 ± 0.004 | 0.037 ± 0.0037 | 0.25 ± 0.027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hector, R.E.; Mertens, J.A.; Nichols, N.N. Identification of Mutations Responsible for Improved Xylose Utilization in an Adapted Xylose Isomerase Expressing Saccharomyces cerevisiae Strain. Fermentation 2022, 8, 669. https://doi.org/10.3390/fermentation8120669
Hector RE, Mertens JA, Nichols NN. Identification of Mutations Responsible for Improved Xylose Utilization in an Adapted Xylose Isomerase Expressing Saccharomyces cerevisiae Strain. Fermentation. 2022; 8(12):669. https://doi.org/10.3390/fermentation8120669
Chicago/Turabian StyleHector, Ronald E., Jeffrey A. Mertens, and Nancy N. Nichols. 2022. "Identification of Mutations Responsible for Improved Xylose Utilization in an Adapted Xylose Isomerase Expressing Saccharomyces cerevisiae Strain" Fermentation 8, no. 12: 669. https://doi.org/10.3390/fermentation8120669
APA StyleHector, R. E., Mertens, J. A., & Nichols, N. N. (2022). Identification of Mutations Responsible for Improved Xylose Utilization in an Adapted Xylose Isomerase Expressing Saccharomyces cerevisiae Strain. Fermentation, 8(12), 669. https://doi.org/10.3390/fermentation8120669