
Untargeted Metabolomics Combined with Metabolic Flux Analysis Reveals the Mechanism of Sodium Citrate for High S-Adenosyl-Methionine Production by Pichia pastoris

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Cultivations
2.2. Analysis Methods
2.3. Sample Preparation and LC-MS/MS Analysis
2.4. Metabolic Flux Analysis
2.5. Data Processing and Statistical Analysis
3. Results and Discussion
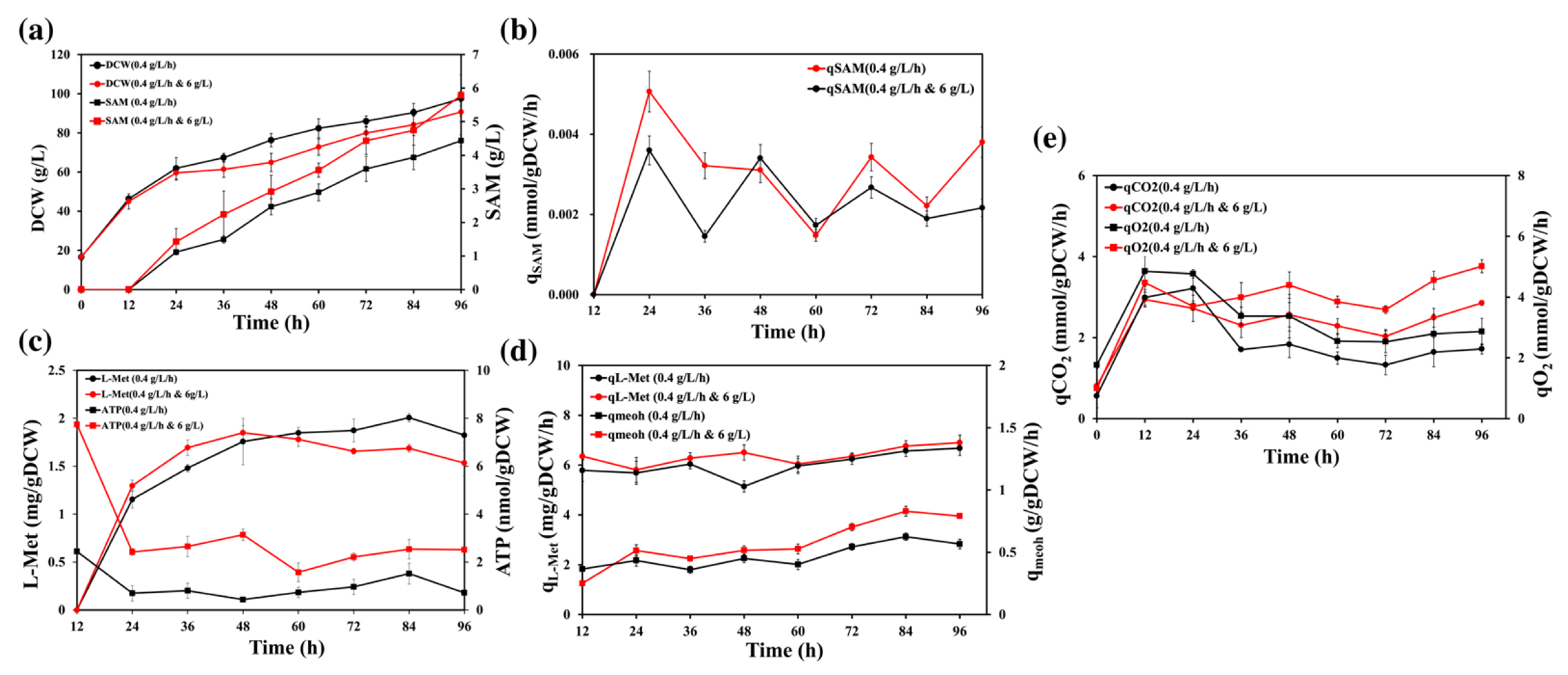
3.1. Comparison of Physiology under Different L-methionine Feeding Rate
3.2. Comparison of Physiology under Sodium Citrate Supplement
3.3. Exploring the Effects of Sodium Citrate Supplementation on Cellular Metabolism through Metabolomics
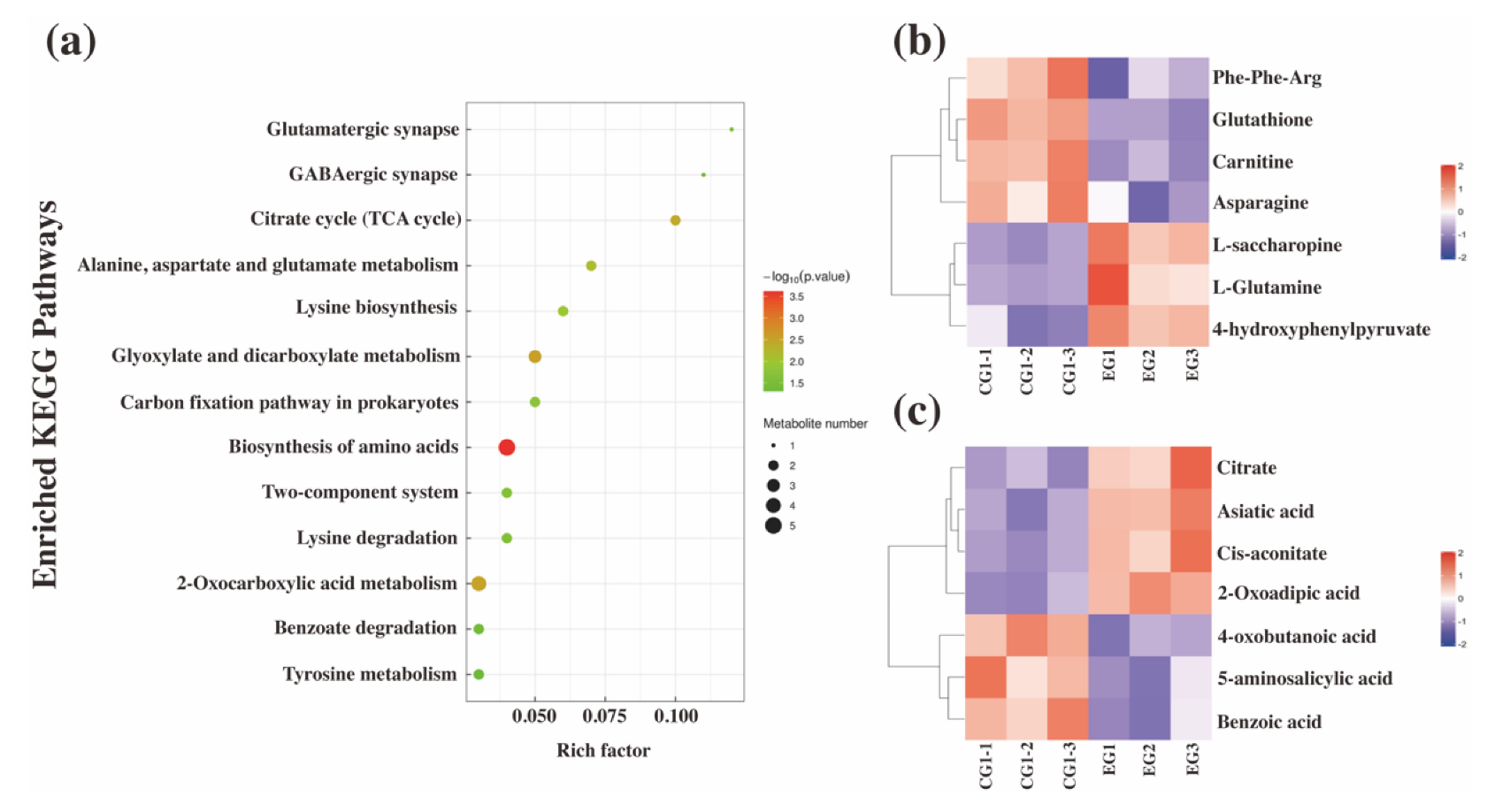
3.3.1. Differential Metabolite Screening and Pathway Annotation
3.3.2. Clustering Analysis of Differential Metabolites Related to Amino Acid Metabolism
3.3.3. Clustering Analysis of Differential Metabolites Related to Organic Acid Metabolism
3.4. Metabolic Flux Analysis
3.5. Analysis of Metabolic Regulatory Mechanisms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mischoulon, D.; Alpert, J.E.; Arning, E.; Bottiglieri, T.; Fava, M.; Papakostas, G.I. Bioavailability of S-adenosyl methionine and impact on response in a randomized, double-blind, placebo-controlled trial in major depressive disorder. J. Clin. Psychiat. 2012, 73, 843–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.W.; Lou, S.Y.; Fan, L.H.; Zhang, X.; Tan, T.W. Control of ATP concentration in Escherichia coli using synthetic small regulatory RNAs for enhanced S-adenosylmethionine production. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.L.; Wang, Z.L.; Cai, H.B.; Zhou, C.L. Progress in the microbial production of S-adenosyl-L-methionine. World J. Microbiol. Biotechnol. 2016, 32, 153. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Gou, X.; Hu, H.; Xu, Q.; Lu, Y.; Cheng, J. Enhanced S-adenosyl-l-methionine production in Saccharomyces cerevisiae by spaceflight culture, overexpressing methionine adenosyltransferase and optimizing cultivation. J. Appl. Microbiol. 2012, 112, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Parashar, S.; Cheishvili, D.; Arakelian, A.; Hussain, Z.; Tanvir, I.; Khan, H.A.; Szyf, M.; Rabbani, S.A. S-adenosylmethionine blocks osteosarcoma cells proliferation and invasion in vitro and tumor metastasis in vivo: Therapeutic and diagnostic clinical applications. Cancer Med. 2015, 4, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Qian, J.C.; Zhuang, Y.P.; Zhang, S.L.; Li, Y.R. Progress in the research of S-adenosyl-L-methionine production. Appl. Microbiol. Biotechnol. 2013, 97, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Macauley-patrick, S.; Fazenda, M.L.; Mcneil, B.; Harvey, L.M. Heterologous protein production using the Pichia pastoris expression system. Yeast 2005, 22, 249–270. [Google Scholar] [CrossRef]
- Hayakawa, K.; Matsuda, F.; Shimizu, H. Metabolome analysis of Saccharomyces cerevisiae and optimization of culture medium for S-adenosyl-L-methionine production. AMB Express. 2016, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Wang, D.H.; Wei, G.Y.; Wang, C.F. Improved co-production of S-adenosylmethionine and glutathione using citrate as an auxiliary energy substrate. Bioresour. Technol. 2013, 131, 28–32. [Google Scholar] [CrossRef]
- Qin, X.L.; Lu, J.J.; Zhang, Y.; Wu, X.L.; Qiao, X.F.; Wang, Z.P.; Chu, J.; Qian, J.C. Engineering Pichia pastoris to improve S-adenosyl-l-methionine production using systems metabolic strategies. Biotechnol. Bioeng. 2020, 117, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Q.; Chu, J.; Zhang, S.L.; Zhuang, Y.P.; Wang, Y.H.; Zhu, S.; Zhu, Z.G.; Yuan, Z. A novel feeding strategy during the production phase for enhancing the enzymatic synthesis of S-adenosyl-L-methionine by methylotrophic Pichia pastoris. Enzyme Microb. Technol. 2007, 40, 669–674. [Google Scholar] [CrossRef]
- Hu, H.; Qian, J.C.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. Optimization of L-methionine feeding strategy for improving S-adenosyl-L-methionine production by methionine adenosyltransferase overexpressed Pichia pastoris. Appl. Microbiol. Biot. 2009, 83, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Ke, X.; Hong, M.; Huang, M.Z.; Chen, C.C.; Tian, X.W.; Hang, H.F.; Chu, J. Exploring the metabolic fate of propanol in industrial erythromycin-producing strain via 13C labeling experiments and enhancement of erythromycin production by rational metabolic engineering of Saccharopolyspora erythraea. Biochem. Bioph. Res. Commun. 2021, 542, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Z.; Liu, X.Y.; Huang, M.Z.; Xia, J.Y.; Chu, J.; Zhuang, Y.P.; Zhang, S.L.; Noorman, H. Integrated isotope-assisted metabolomics and 13C metabolic flux analysis reveals metabolic flux redistribution for high glucoamylase production by Aspergillus niger. Microb. Cell Fact. 2015, 14, 147. [Google Scholar] [CrossRef] [Green Version]
- Abdelrazig, S.; Ortori, C.A.; Doherty, M.; Valdes, A.M.; Chapman, V.; Barrett, D.A. Metabolic signatures of osteoarthritis in urine using liquid chromatography-high resolution tandem mass spectrometry. Metabolomics 2021, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Huang, M.Z.; Guo, M.L.; Qian, J.C.; Lin, W.L.; Chu, J.; Zhuang, Y.P.; Zhang, S.L. Combined 13C-assisted metabolomics and metabolic flux analysis reveals the impacts of glutamate on the central metabolism of high β-galactosidase-producing Pichia pastoris. Bioresour. Bioprocess. 2016, 3, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.S.; Huang, M.Z.; Lu, J.J.; Qian, J.C.; Lin, W.L.; Chu, J.; Zhuang, Y.P.; Zhang, S.L. Impacts of high β-galactosidase expression on central metabolism of recombinant Pichia pastoris GS115 using glucose as sole carbon source via (13)C metabolic flux analysis. J. Biotechnol. 2014, 187, 124–134. [Google Scholar] [CrossRef]
- Jordà, J.; Suarez, C.; Carnicer, M.; ten Pierick, A.; Heijnen, J.J.; van Gulik, W.; Ferrer, P.; Albiol, J.; Wahl, A. Glucose-methanol co-utilization in Pichia pastoris studied by metabolomics and instationary ¹³C flux analysis. BMC Syst. Biol. 2013, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Heyland, J.; Fu, J.N.; Blank, L.M.; Schmid, A. Quantitative physiology of Pichia pastoris during glucose-limited high-cell density fed-batch cultivation for recombinant protein production. Biotechnol. Bioeng. 2010, 107, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Heyland, J.; Fu, J.N.; Blank, L.M.; Schmid, A. Carbon metabolism limits recombinant protein production in Pichia pastoris. Biotechnol. Bioeng. 2011, 108, 1942–1953. [Google Scholar] [CrossRef]
- Xu, F.; Lu, J.; Ke, X.; Shao, M.H.; Huang, M.Z.; Chu, J. Reconstruction of the Genome-Scale Metabolic Model of Saccharopolyspora erythraea and Its Application in the Overproduction of Erythromycin. Metabolites 2022, 12, 509. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Huang, M.Z.; Lu, H.Z.; Qian, J.C.; Lin, W.L.; Chu, J.; Zhuang, Y.P.; Zhang, S.L. Comprehensive reconstruction and evaluation of Pichia pastoris genome-scale metabolic model that accounts for 1243 ORFs. Bioresour. Bioprocess. 2017, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soberon, M.; Olamendi, J.; Rodriguez, L.; González, A. Role of glutamine aminotransferase in glutamine catabolism by Saccharomyces cerevisiae under microaerophilic conditions. J. Gen. Microbiol. 1989, 135, 2693–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayyad-Kazan, M.; Feller, A.; Bodo, E.; Boeckstaens, M.; Marini, A.M.; Dubois, E.; Georis, I. Yeast nitrogen catabolite repression is sustained by signals distinct from glutamine and glutamate reservoirs. Mol. Microbiol. 2016, 99, 360–379. [Google Scholar] [CrossRef]
- Ding, M.Z.; Wang, X.; Yang, Y.; Yuan, Y.J. Metabolomic study of interactive effects of phenol, furfural, and acetic acid on Saccharomyces cerevisiae. OMICS 2011, 15, 647–653. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- He, J.Y.; Deng, J.J.; Zheng, Y.H.; Gu, J. A synergistic effect on the production of S-adenosyl-L-methionine in Pichia pastoris by knocking in of S-adenosyl-L-methionine synthase and knocking out of cystathionine-beta synthase. J. Biotechnol. 2006, 126, 519–527. [Google Scholar] [CrossRef]
- Sulheim, S.; Kumelj, T.; van Dissel, D.; Salehzadeh-Yazdi, A.; Du, C.; van Wezel, G.P.; Nieselt, K.; Almaas, E.; Wentzel, A.; Kerkhoven, E.J. Enzyme-Constrained Models and Omics Analysis of Streptomyces coelicolor Reveal Metabolic Changes that Enhance Heterologous Production. iScience 2020, 23, 101525. [Google Scholar] [CrossRef]
- Pekala, J.; Patkowska-Sokoła, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochyński, S.; Librowski, T. L-carnitine--metabolic functions and meaning in humans life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef]
- Pook, C.; Diep, T.T.; Yoo, M.J.Y. Simultaneous Quantification of Organic Acids in Tamarillo (Solanum betaceum) and Untargeted Chemotyping Using Methyl Chloroformate Derivatisation and GC-MS. Molecules 2022, 27, 1314. [Google Scholar] [CrossRef]
- Shibata, K.; Yasui, M.; Sano, M.; Fukuwatari, T. Fluorometric determination of 2-oxoadipic acid, a common metabolite of tryptophan and lysine, by high-performance liquid chromatography with pre-chemical derivatization. Biosci. Biotechnol. Biochem. 2011, 75, 185–187. [Google Scholar] [CrossRef]
- Koning, W.; Gleeson, M.A.G.; Harder, W.; Dijkhuizen, L. Regulation of methanol metabolism in the yeast Hansenula polymorpha. Arch. Microbiol. 1987, 147, 375–382. [Google Scholar] [CrossRef]
- Gosalvez, M.; Pérez-García, J.; Weinhouse, S. Competition for ADP between pyruvate kinase and mitochondrial oxidative phosphorylation as a control mechanism in glycolysis. Eur. J. Biochem. 1974, 46, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.; Mach, R.L.; Mach-Aigner, A.R. The pentose phosphate pathway in industrially relevant fungi: Crucial insights for bioprocessing. Appl. Microbiol. Biotechnol. 2021, 105, 4017–4031. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Zhou, L.S.; Zhao, Y.Z.; Wang, S.W.; Chen, L.L.; Liu, L.X.; Ling, Z.Q.; Hu, F.J.; Sun, Y.P.; Zhang, J.Y.; et al. Regulation of G6PD acetylation by SIRT2 and KAT9 modulates NADPH homeostasis and cell survival during oxidative stress. EMBO J. 2014, 33, 1304–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.L.; Wang, Z.; Wang, Z.L.; Dou, J.; Zhou, C.L. Improving methionine and ATP availability by MET6 and SAM2 co-expression combined with sodium citrate feeding enhanced SAM accumulation in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2016, 32, 56. [Google Scholar] [CrossRef]
- Anderson, D.H.; Duckworth, H.W. In vitro mutagenesis of Escherichia coli citrate synthase to clarify the locations of ligand binding sites. J. Biol. Chem. 1988, 263, 2163–2169. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Parameters | Control Group | Experimental Group |
|---|---|---|
| qMeOH (mmol/gDCW/h) | 9.25 ± 0.09 | 11.24 ± 0.15 |
| qSAM (mmol/gDCW/h) | 0.0015 ± 0.0002 | 0.0032 ± 0.0003 |
| qTri.Ci (mmol/gDCW/h) | 0 | 0.160 |
| qL-Met (mmol/gDCW/h) | 0.041 ± 0.0029 | 0.035 ± 0.0021 |
| qCO2 (mmol/gDCW/h) | 9.29 ± 0.31 | 12.04 ± 0.28 |
| qO2 (mmol/gDCW/h) | 19.49 ± 0.15 | 25.62 ± 0.36 |
| Carbon recoveries 1 (%) | 98.58 ± 0.08 | 97.70 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Xu, F.; Song, W.; Dong, L.; Qian, J.; Huang, M. Untargeted Metabolomics Combined with Metabolic Flux Analysis Reveals the Mechanism of Sodium Citrate for High S-Adenosyl-Methionine Production by Pichia pastoris. Fermentation 2022, 8, 681. https://doi.org/10.3390/fermentation8120681
Xu W, Xu F, Song W, Dong L, Qian J, Huang M. Untargeted Metabolomics Combined with Metabolic Flux Analysis Reveals the Mechanism of Sodium Citrate for High S-Adenosyl-Methionine Production by Pichia pastoris. Fermentation. 2022; 8(12):681. https://doi.org/10.3390/fermentation8120681
Chicago/Turabian StyleXu, Wentao, Feng Xu, Weijing Song, Le Dong, Jiangchao Qian, and Mingzhi Huang. 2022. "Untargeted Metabolomics Combined with Metabolic Flux Analysis Reveals the Mechanism of Sodium Citrate for High S-Adenosyl-Methionine Production by Pichia pastoris" Fermentation 8, no. 12: 681. https://doi.org/10.3390/fermentation8120681
APA StyleXu, W., Xu, F., Song, W., Dong, L., Qian, J., & Huang, M. (2022). Untargeted Metabolomics Combined with Metabolic Flux Analysis Reveals the Mechanism of Sodium Citrate for High S-Adenosyl-Methionine Production by Pichia pastoris. Fermentation, 8(12), 681. https://doi.org/10.3390/fermentation8120681