
Qualitative Screening of Yeast Biodiversity for Hydrolytic Enzymes Isolated from the Gastrointestinal Tract of a Coprophage “Gymnopleurus sturmi” and Dung of Ruminants
 , ,
, ,  ,
,
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Yeasts | Applications | References |
|---|---|---|---|
| Cellulases | Aureobasidium pullulans 98 | Food, chemical, textile, paper, and biofuel industries | [17,18] |
| Pectinase | Kluveromyces marxianus Metschnikowia pulcherrima | Wine, cider, and fruit juice industries | [19,20] |
| Lipase | Aureobasidium pullulans HN2.3 | Food, wastewater treatment, cosmetics, pharmaceuticals, leather processing, and biofuel industries | [21] |
| ß-glucosidase | Guehomyces pullulans 17-1 | Food industry | [22] |
| Catalase | Saccharomyces cerevisiae | Food industry | [23] |
| Inulinase | Pichia guilliermondii | Food, beverage, and biofuel industries | [24] |
| Urease | Cryptococcus gattii | Diagnostic kits, beverages, and animal feed | [25] |
| Gelatinase | Trichosporon pullulans | Food, chemical, and medical industries | [26] |
| Protease | Rhodotorula mucilaginosa L7 | Clinical applications, food, beverage, and leather processing industries | [27] |
| Amylase | Aureobasidium pullulans N13dSaccharomyces cerevisiae | Food, textile, paper, detergent industry, medical, and pharmaceutical industries | [28,29] |
| Tannase | Kluyveromyces marxianus | Feed, food, beverages, brewing, pharmaceutical, chemical, cosmetic, and leather industries | [30] |
2. Materials and Methods
2.1. Sampling
2.2. Qualitative Screening of Isolates for Hydrolytic Enzymes
2.2.1. Oxidation of Phenolic Substrates
2.2.2. Cellulase Activity
2.2.3. Amylase Activity
2.2.4. Inulinase Activity
2.2.5. β-Glucosidase Activity
2.2.6. Pectinase Activity
2.2.7. Lipase Activity
2.2.8. Gelatinase Activity
2.2.9. Urease Activity
2.2.10. Protease Activity
2.2.11. Catalase Activity
2.2.12. Tannase Activity
2.3. Study of Physiological Characteristics of Cellulolytic Isolates
2.3.1. Thermotolerance
2.3.2. pH Tolerance
2.3.3. Utilization of Carbon Sources
2.3.4. Glucose Tolerance of Yeasts
2.3.5. Tolerance to Ethanol
2.4. Molecular Identification
2.5. Multiple Sequence Alignment and Phylogenetic Tree
3. Results and Discussion
3.1. Distribution of Isolated Yeasts
3.2. Oxidation of Phenolic Substrates
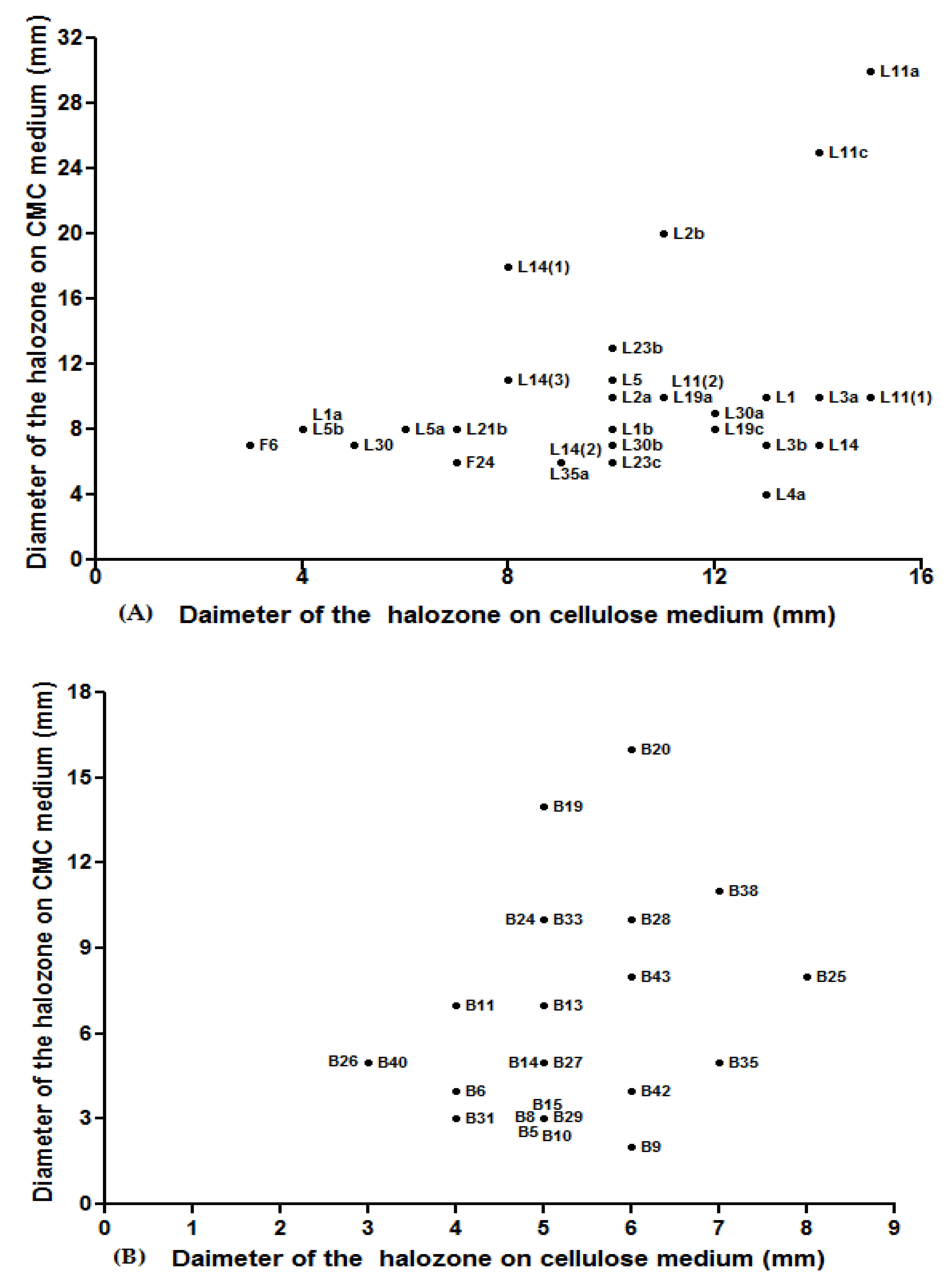
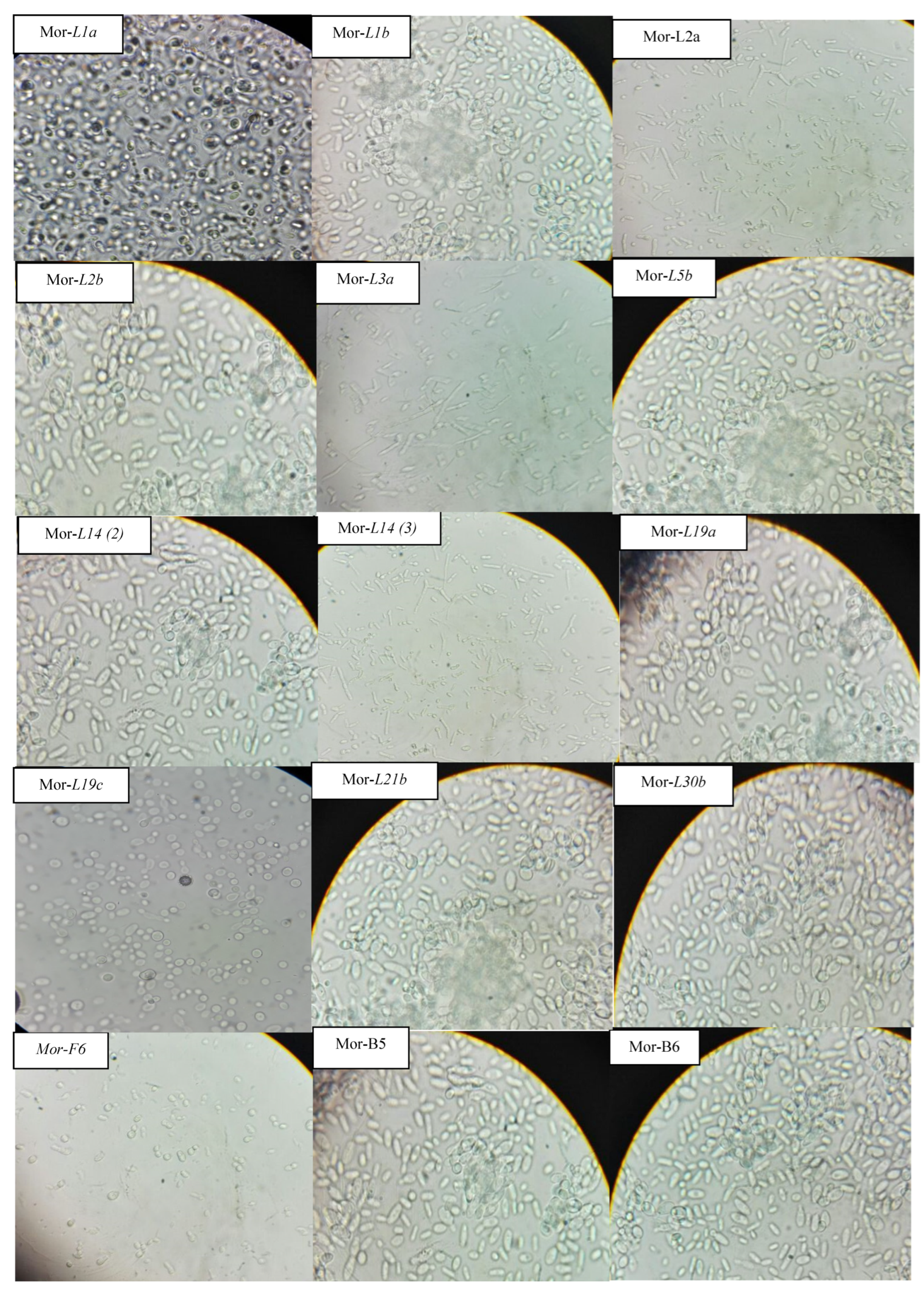
3.3. Screening of Cellulolytic Isolates
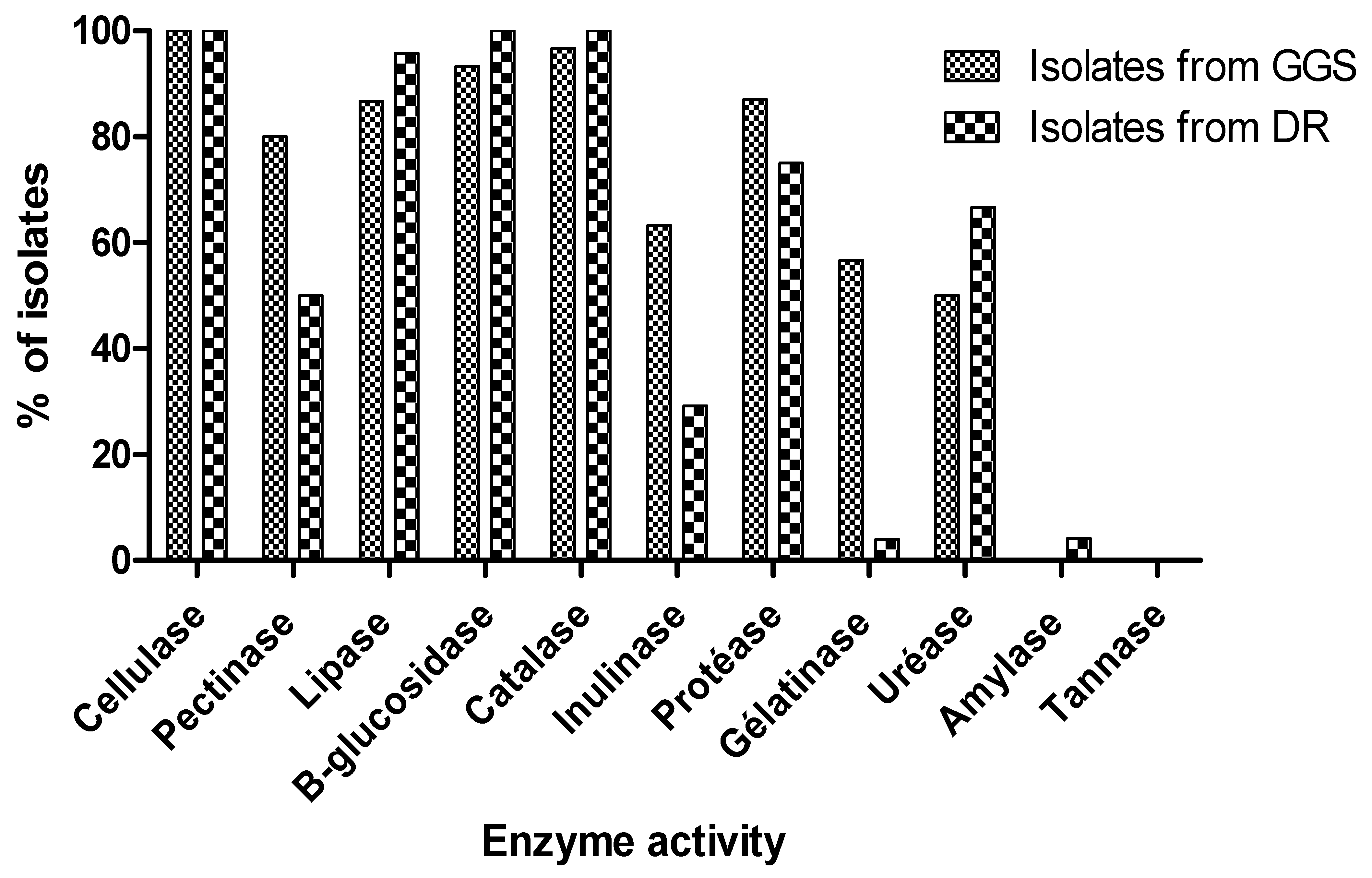
3.4. Hydrolytic Enzymes
3.5. Study of Physiological Characteristics
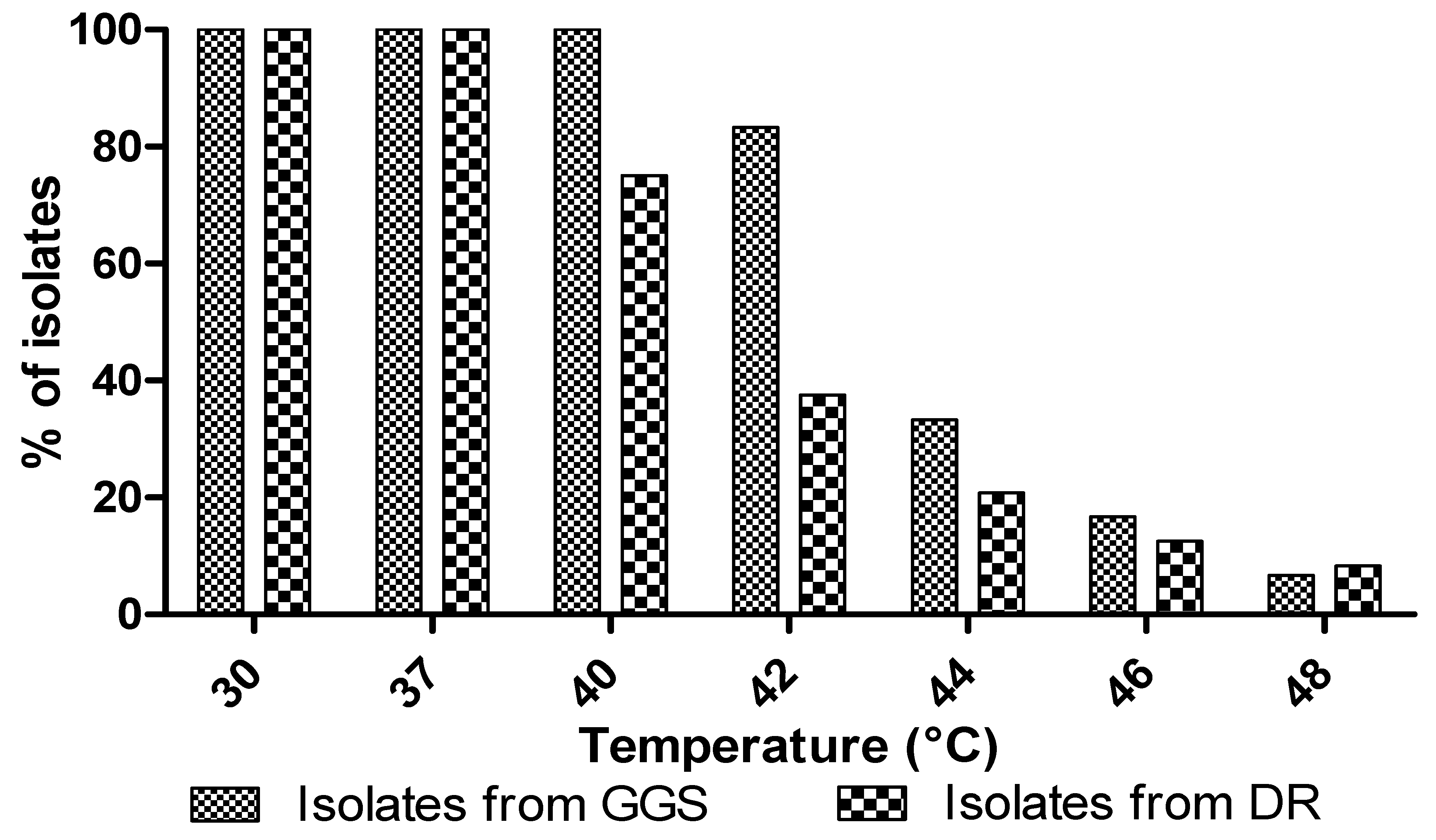
3.5.1. Temperature
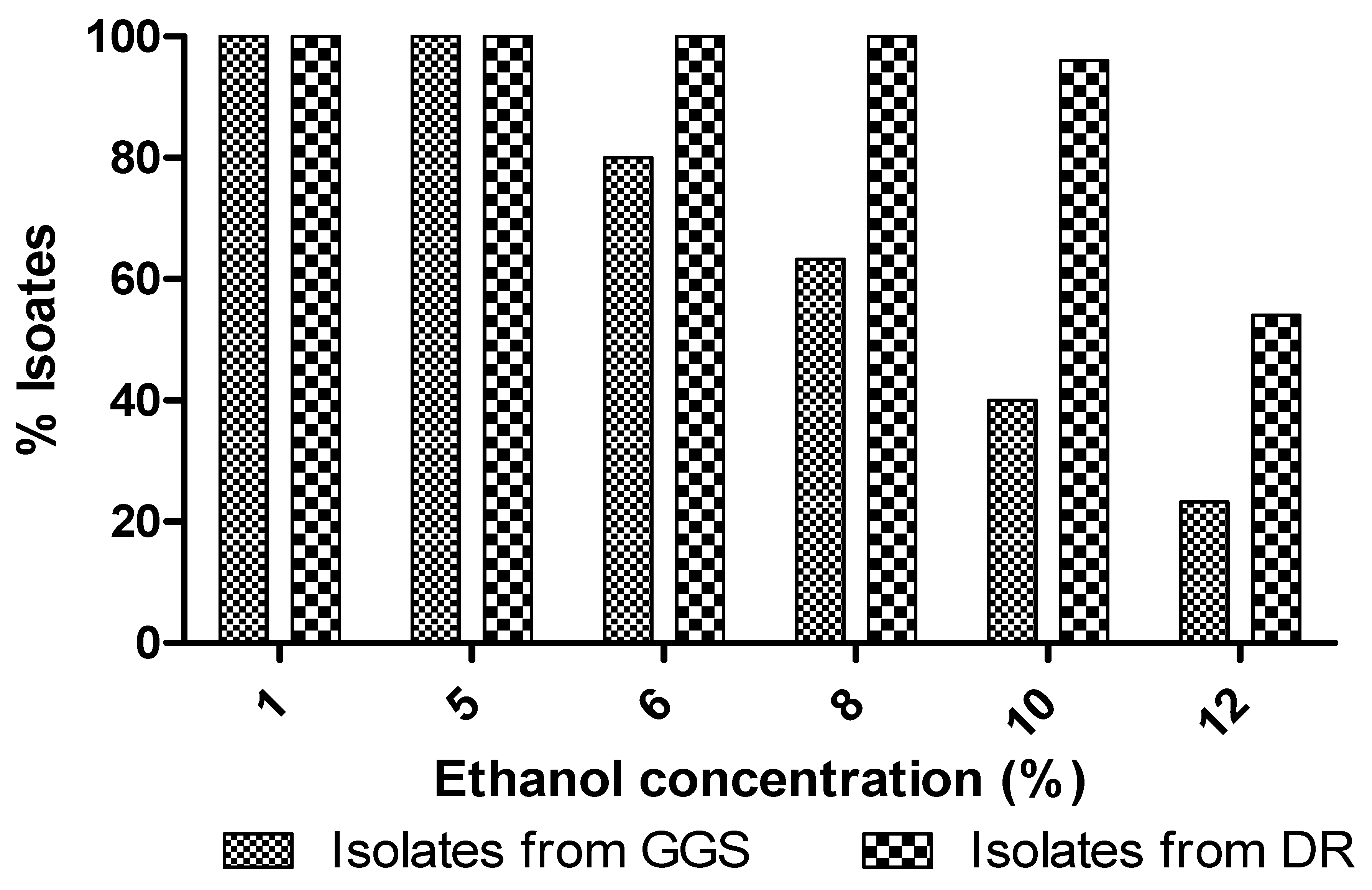
3.5.2. Exogenous Ethanol
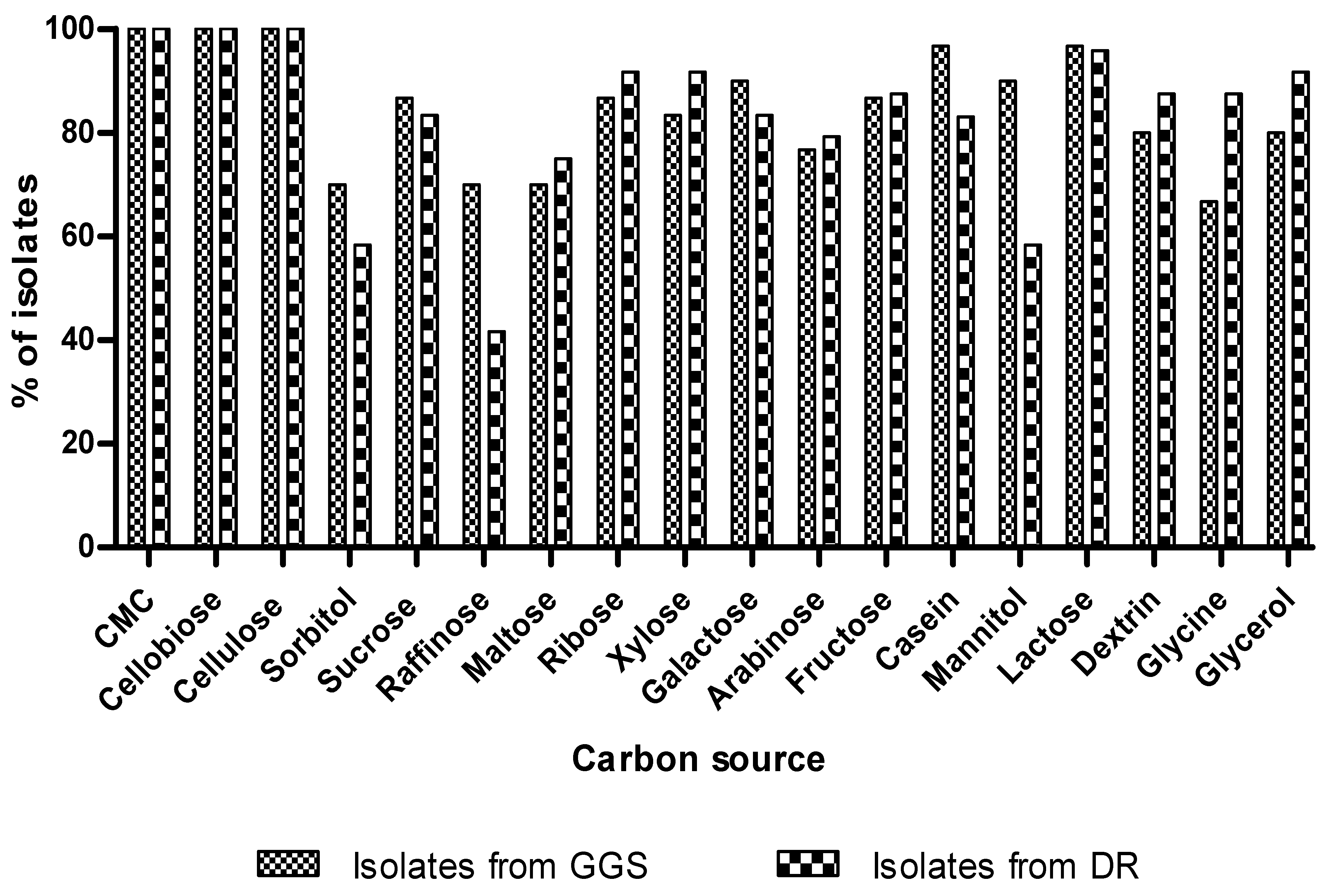
3.5.3. Utilization of Carbon Sources
3.5.4. Osmotolerance
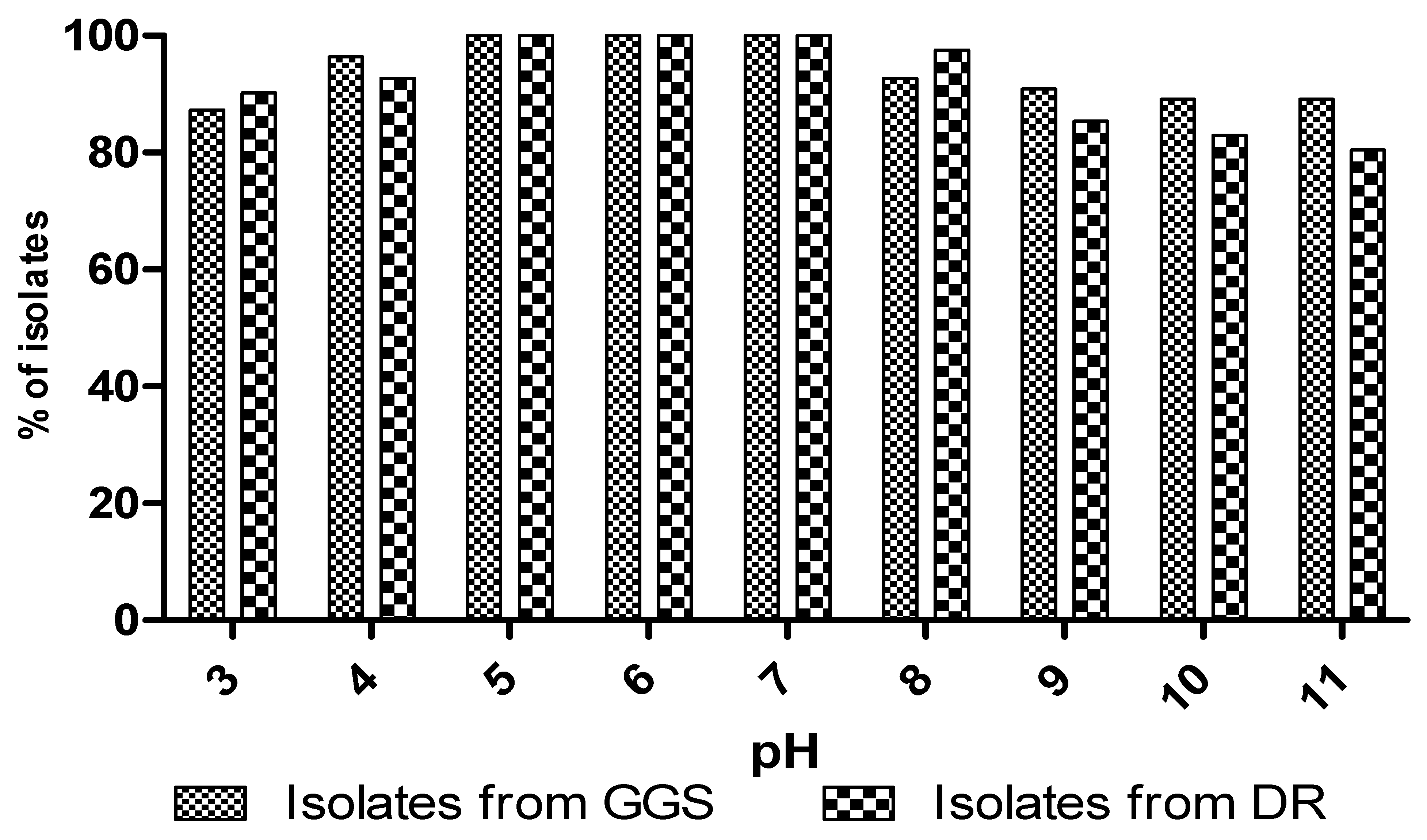
3.5.5. Effect of pH
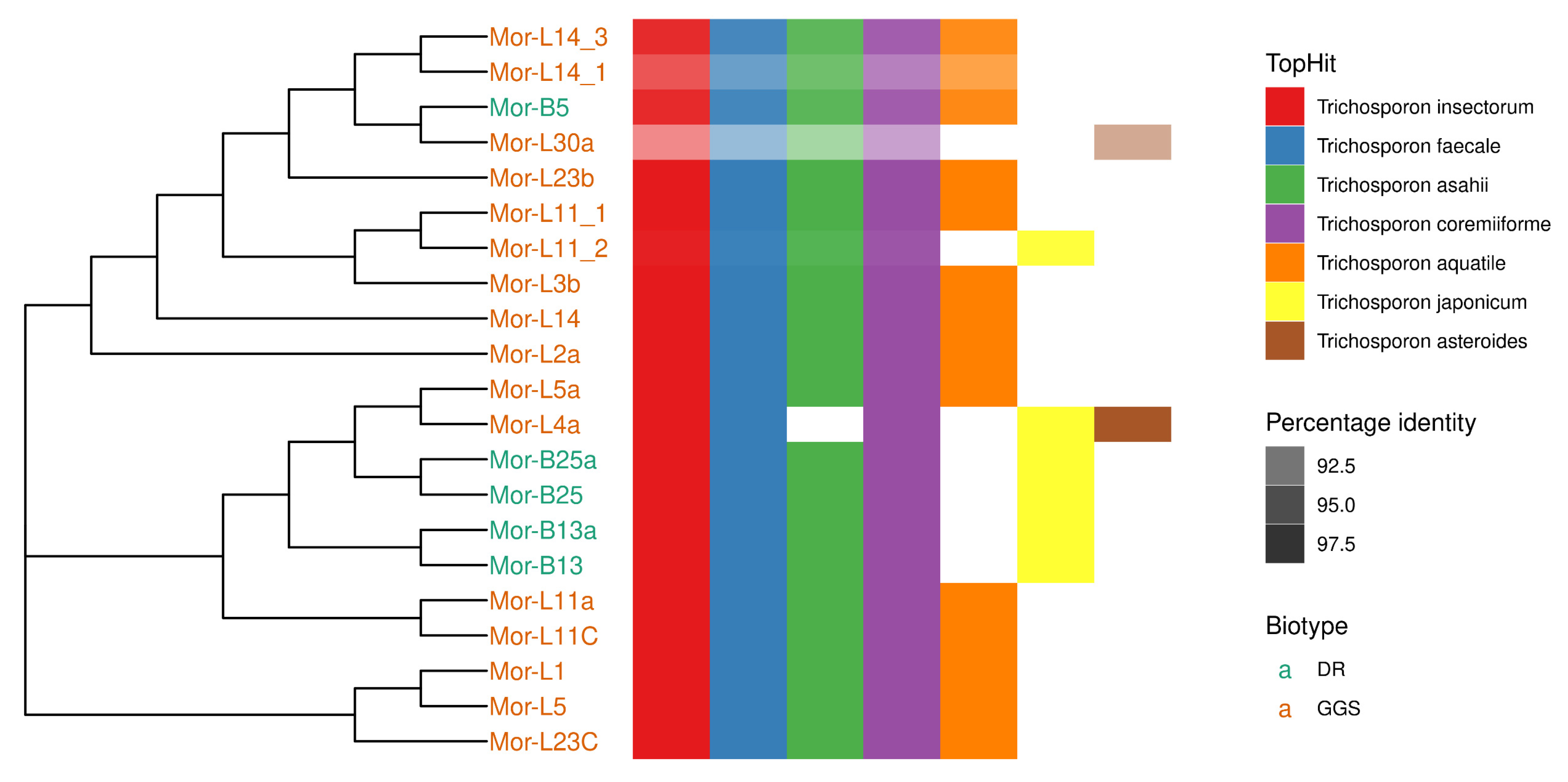
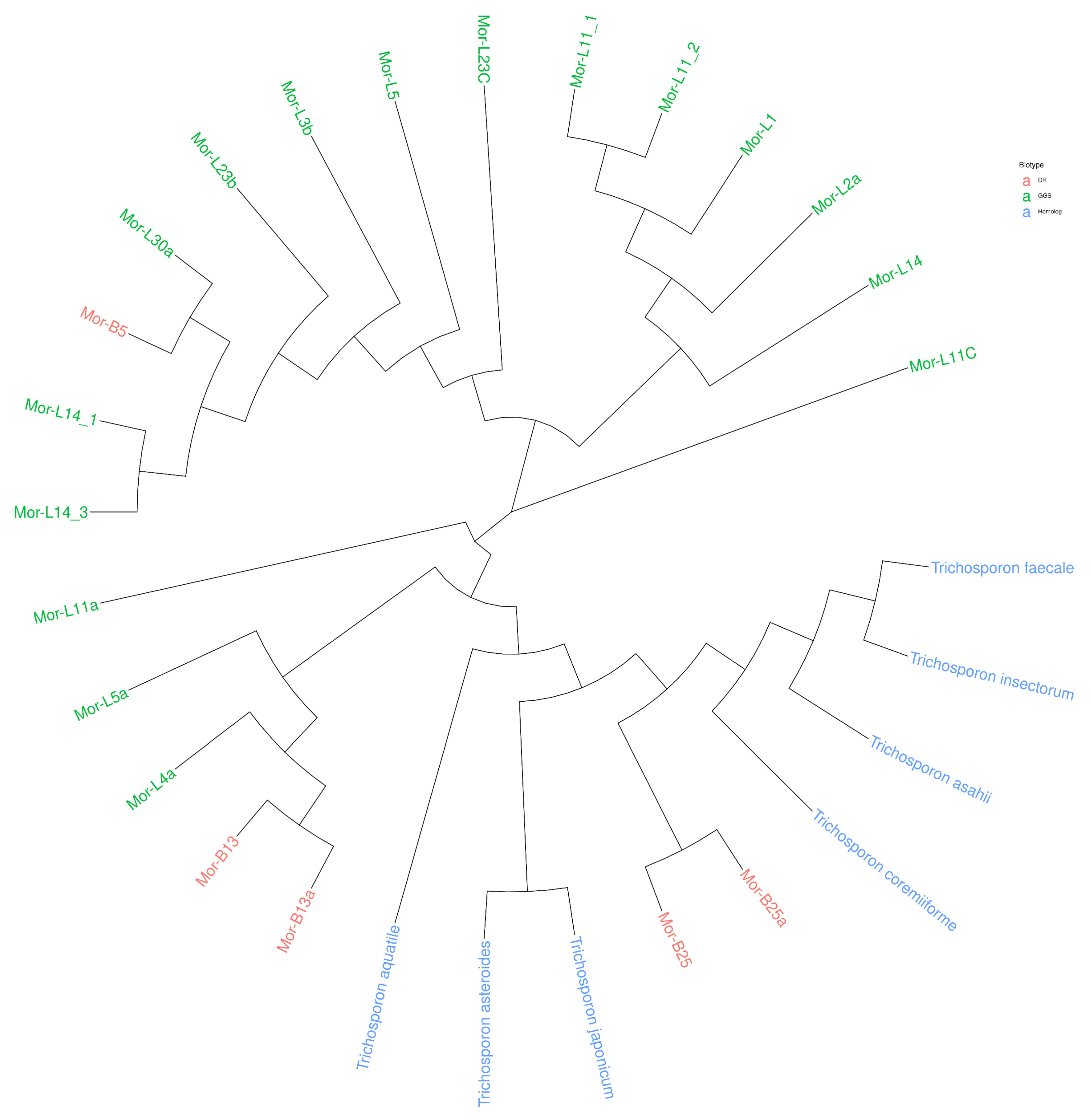
3.6. Molecular Identification
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R. The ecology of insect–yeast relationships and its relevance to human industry. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef] [Green Version]
- Willaert, R.G. Yeast Biotechnology 4.0. Fermentation 2021, 7, 69. [Google Scholar] [CrossRef]
- Satyanarayana, T.; Kunze, G. Yeast Biotechnology: Diversity and Applications; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Chabasse, P.; Bouchara, D.; de Gentile, J.P.; Brun, L.; Cimon, S.; Penn, B. Les Moisissures D’Intérêt Médical. Bioforma 2002, 161. [Google Scholar]
- Lee, Y.-J.; Choi, Y.-R.; Lee, S.-Y.; Park, J.-T.; Shim, J.-H.; Park, K.-H.; Kim, J.-W. Screening Wild Yeast Strains for Alcohol Fermentation from Various Fruits Screening Wild Yeast Strains for Alcohol Fermentation from Various Fruits. Mycobiology 2018, 39, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanini, I. Yeast-insect associations: It takes guts Irene. Yeast 2018, 35, 315–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandi, R.; Velu, G.; Devi, P.; Dananjeyan, B. Isolation and Screening of Soil Yeasts for Plant Growth Promoting Traits. Madras Agric. J. 2019, 106, 143–148. [Google Scholar] [CrossRef]
- Quan, A.S.; Eisen, B.M. The Ecology of the Drosophila-Yeast Mutualism in Wineries. PLoS ONE 2018, 13, e0196440. [Google Scholar] [CrossRef] [Green Version]
- Saberi, S.; Cliff, M.A.; Van Vuuren, H.J.J. Impact of Mixed S. Cerevisiae Strains on the Production of Volatiles and Estimated Sensory pro Fi Les of Chardonnay Wines. Food Res. Int. 2012, 48, 725–735. [Google Scholar] [CrossRef]
- Mitsui, R.; Nishikawa, R.; Yamada, R.; Matsumoto, T.; Ogino, H. Construction of yeast producing patchoulol by global metabolic engineering strategy. Biotechnol. Bioeng. 2020, 117, 1348–1356. [Google Scholar] [CrossRef]
- Romo-sánchez, S.; Alves-baf, M.; Arévalo-villena, M.; Úbeda-iranzo, J.; Briones-pérez, A. Yeast Biodiversity from Oleic Ecosystems: Study of Their Biotechnological Properties. Food Microbiol. 2010, 27, 487–492. [Google Scholar] [CrossRef]
- Bhat, M.K. Cellulases and related enzymes in biotechnology. Biotechnol. Adv. 2000, 18, 355–383. [Google Scholar] [CrossRef]
- Lumaret, J.-P.; Kirk, A. Ecology of dung beetles in the French Mediterranean region (Coleoptera: Scarabaeinae). Acta Zool. Mex. 1987, 24, 1–55. [Google Scholar]
- Yun, J.-H.; Roh, S.W.; Tae, W.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect Gut Bacterial Diversity Determined by Environmental Habitat, Diet, Developmental Stage, and Phylogeny of Host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [Green Version]
- Darwesh, O.M.; El-Maraghy, S.H.; Abdel-Rahman, H.M.; Zaghloul, R.A. Improvement of paper wastes conversion to bioethanol using novel cellulose degrading fungal isolate. Fuel 2020, 262, 116518. [Google Scholar] [CrossRef]
- Rosyidaa, V.T.; Indrianingsiha, W.; Maryanaa, R.; Wahono, S.K. Effect of Temperature and Fermentation Time of Crude Cellulase Production by Trichoderma reesei on Straw Substrate. Energy Procedia 2015, 65, 368–371. [Google Scholar] [CrossRef] [Green Version]
- Rong, Y.; Zhang, L.; Chi, Z.; Wang, X. A Carboxymethyl Cellulase from a Marine Yeast (Aureobasidium Pullulans 98): Its Purification, Characterization, Gene Cloning and Carboxymethyl Cellulose Digestion. J. Ocean Univ. China 2015, 14, 913–921. [Google Scholar] [CrossRef]
- Thongekkaew, J.; Kongsanthia, J. Screening and Identification of Cellulase Producing Yeast from Rongkho Forest, Ubon Ratchathani University. Bioeng. Biosci. 2016, 4, 29–33. [Google Scholar] [CrossRef]
- Da Silva, E.G.; Borges, M.D.F.; Medina, C.; Hilsdorf Piccoli, R.; Freitas Schwan, R. Pectinolytic Enzymes Secreted by Yeasts from Tropical Fruits. FEMS Yeast Res. 2005, 5, 859–865. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Conchillo, L.B.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Selection and Use of Pectinolytic Yeasts for Improving Clarification and Phenolic Extraction in Winemaking. Int. J. Food Microbiol. 2016, 223, 1–8. [Google Scholar] [CrossRef]
- Liu, Z.; Chi, Z.; Wang, L.; Li, J. Production, Purification and Characterization of an Extracellular Lipase from Aureobasidium Pullulans HN2.3 with Potential Application for the Hydrolysis of Edible Oils. Biochem. Eng. J. 2008, 40, 445–451. [Google Scholar] [CrossRef]
- Song, C.; Liu, G.L.; Xu, J.L.; Chi, Z.M. Purification and Characterization of Extracellular β-Galactosidase from the Psychrotolerant Yeast Guehomyces Pullulans 17-1 Isolated from Sea Sediment in Antarctica. Process Biochem. 2010, 45, 954–960. [Google Scholar] [CrossRef]
- D’Souza, S.F.; Nadkarni, G.B. Immobilized catalase-containing yeast cells: Preparation and enzymatic properties. Biotechnol. Bioeng. 1980, 22, 2191–2205. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Zhang, T.; Chi, Z.; Sheng, J.; Li, J.; Wang, X. Purification and Characterization of Extracellular Inulinase from a Marine Yeast Pichia Guilliermondii and Inulin Hydrolysis by the Purified Inulinase. Biotechnol. Bioprocess Eng. 2008, 13, 533–539. [Google Scholar] [CrossRef]
- Feder, V.; Kmetzsch, L.; Christian Staats, C.; Vidal-Figueiredo, N.; Ligabue-Braun, R.; Carlini, C.R.; Vainstein, M.H. Cryptococcus gattii urease as a virulence factor and the relevance of enzymatic activity in cryptococcosis pathogenesis. FEBS J. 2015, 282, 1406–1418. [Google Scholar] [CrossRef]
- Loperena, L.; Soria, V.; Varela, H.; Lupo, S.; Bergalli, A.; Guigou, M.; Pellegrino, A.; Bernardo, A.; Calvino, A.; Rivas, F.; et al. Extracellular Enzymes Produced by Microorganisms Isolated from Maritime Antarctica. World J. Microbiol. Biotechnol. 2012, 28, 2249–2256. [Google Scholar] [CrossRef]
- Lilao, J.; Mateo, J.J.; Maicas, S. Biotechnological Activities from Yeasts Isolated from Olive Oil Mills. Eur. Food Res. Technol. 2015, 240, 357–365. [Google Scholar] [CrossRef]
- Li, H.; Chi, Z.; Wang, X.; Ma, C. Amylase Production by the Marine Yeast Aureobasidium Pullulans N13d. J. Ocean Univ. China 2007, 6, 60–65. [Google Scholar] [CrossRef]
- Cripwell, R.A.; Rose, S.H.; Viljoen-Bloom, M.; Van Zyl, W.H. Improved Raw Starch Amylase Production by Saccharomyces Cerevisiae Using Codon Optimisation Strategies. FEMS Yeast Res. 2019, 19, foy127. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.E.; Fathy, S.A.; Rashad, M.M.; Ezz, M.K.; Mohammed, A.T. Purification and Characterization of a Novel Tannase Produced by Kluyveromyces Marxianus Using Olive Pomace as Solid Support, and Its Promising Role in Gallic Acid Production. Int. J. Biol. Macromol. 2018, 107, 2342–2350. [Google Scholar] [CrossRef]
- Patel, M.A.; Ou, M.S.; Harbrucker, R.; Aldrich, H.C.; Buszko, M.L.; Ingram, L.O.; Shanmugam, K.T. Isolation and Characterization of Acid-Tolerant, Thermophilic Bacteria for Effective Fermentation of Biomass-Derived Sugars to Lactic Acid. Appl. Environ. Microbiol. 2006, 72, 3228–3235. [Google Scholar] [CrossRef] [Green Version]
- Gohel, H.R.; Contractor, C.N.; Ghosh, S.K.; Braganza, V.J.A. Comparative Study of Various Staining Techniques for Determination of Extra Cellular Cellulase Activity on Carboxy Methyl Cellulose (CMC) Agar Plates. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 261–266. [Google Scholar]
- Mishra, S.; Behera, N. Amylase Activity of a Starch Degrading Bacteria Isolated from Soil Receiving Kitchen Wastes. Afr. J. Biotechnol. 2008, 7, 3326–3331. [Google Scholar] [CrossRef]
- Gao, L.; Chi, Z.; Sheng, J.; Wang, L.; Li, J.; Gong, F. Inulinase-Producing Marine Yeasts: Evaluation of Their Diversity and Inulin Hydrolysis by Their Crude Enzymes. Microb. Ecol. 2007, 54, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Villena, M.A.; Úbeda Iranzo, J.F.; Briones Pérez, A.I. β-Glucosidase Activity in Wine Yeasts: Application in Enology. Enzym. Microb. Technol. 2007, 40, 420–425. [Google Scholar] [CrossRef]
- Hankin, L.; Zucker, M.; Sands, D.C. Improved Solid Medium for the Detection and Enumeration of Pectolytic Bacteria. Appl. Microbiol. 1971, 22, 205–209. [Google Scholar] [CrossRef]
- Dhiman, S.; Chapadgaonkar, S. Optimization of Lipase Production Medium for a Bacterial Isolate. Int. J. Chem. Technol. Res. 2013, 5, 2837–2843. [Google Scholar]
- Balan, S.S.; Nethaji, R.; Sankar, S.; Jayalakshmi, S. Production of Gelatinase Enzyme from Bacillus Spp Isolated from the Sediment Sample of Porto Novo Coastal Sites. Asia Pacif. J. Trop. Biomed. 2012, 2, S1811–S1816. [Google Scholar] [CrossRef]
- Geweely, N.S.I. Purification and characterization of intracellular urease enzyme isolated from Rhizopus oryzae. Biotechnology 2006, 5, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; Van Rensburg, P. Screening for the Production of Extracellular Hydrolytic Enzymes by Non-Saccharomyces Wine Yeasts. J. Appl. Microb. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Hernandez, A.; Martin, A.; Aranda, E.; Perez-Nevado, F.; Cordoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–351. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Mostafa, A.M.; Mwafy, E.A.; Darwesh, O.M. Eco-friendly Cellulose Nano Fibers via First Reported Egyptian Humicola Fuscoatra Egyptia X4: Isolation and Characterization. Environ. Nanotechnol. Monit. Manag. 2018, 10, 409–418. [Google Scholar] [CrossRef]
- Thongekkaew, J.; Ikeda, H.; Masaki, K.; Iefuji, H. An acidic and thermostable carboxymethyl cellulase from the yeast Cryptococcus sp. S-2: Purification, characterization and improvement of its recombinant enzyme production by high cell-density fermentation of Pichia pastoris. Protein Expr. Purif. 2008, 60, 140–146. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Darwesh, O.M.; Matter, I.A.; El-Saied, H. Isolation and characterization of non-cellulolytic Aspergillus flavus EGYPTA5 exhibiting selective ligninolytic potential. Biocatal. Agric. Biotechnol. 2019, 17, 160–167. [Google Scholar] [CrossRef]
- Brown, J.; Scholtz, C.H.; Janeau, J.L.; Grellier, S.; Podwojewski, P. Dung beetles (Coleoptera: Scarabaeidae) can improve soil hydrological properties. Appl. Soil Ecol. 2010, 46, 9–16. [Google Scholar] [CrossRef]
- Gonzalo, G.; Colpa, D.I.; Habib, M.H.M.; Fraaijeb, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínkova, L.; Kotik, M.; Markova, E.; Homolka, L. Biodegradation of phenolic compounds by Basidiomycota and its phenol oxidases: A review. Chemosphere 2016, 149, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Tsioulpas, A.; Dimou, D.; Iconomou, D.; Aggelis, G. Phenolic removal in olive oil mill wastewater by strains of Pleurotus spp. in respect to their phenol oxidase (laccase) activity. Bioresour. Technol. 2002, 84, 251–257. [Google Scholar] [CrossRef]
- Garrido Hoyos, S.E.; Martinez Nieto, L.; Camacho Rubio, F.; Ramos Cormenzana, A. Kinetics of aerobic treatment of olive-mill wastewater (OMW) with Aspergillus terreus. Process Biochem. 2002, 37, 1169–1176. [Google Scholar] [CrossRef]
- Fadil, K.; Chahlaoui, A.; Ouahbi, A.; Zaid, A.; Borja, R. Aerobic biodegradation and detoxification of wastewaters from the olive oil industry. Int. Biodeterior. Biodegrad. 2003, 51, 37–41. [Google Scholar] [CrossRef]
- Chi, Z.; Liu, G.-l.; Lu, Y.; Jiang, H.; Chi, Z.-M. Bio-Products Produced by Marine Yeasts and Their Potential Applications. Bioresour. Technol. 2016, 202, 244–252. [Google Scholar] [CrossRef]
- Rai, A.K.; Kumari, R.; Sanjukta, S.; Sahoo, D. Production of Bioactive Protein Hydrolysate Using the Yeasts Isolated from Soft Chhurpi. Bioresour. Technol. 2016, 219, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Jay-Robert, P.; Errouissi, F.; Lumaret, J.P. Temporal Coexistence of Dung-Dweller and Soil-Digger Dung Beetles (Coleoptera, Scarabaeoidea) in Contrasting Mediterranean Habitats. Bull. Èntomol. Res. 2008, 98, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Bautista-gallego, J.; Rodríguez-gómez, F.; Barrio, E.; Querol, A. International Journal of Food Microbiology Exploring the Yeast Biodiversity of Green Table Olive Industrial Fermentations for Technological Applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef]
- Aissam, H.; Errachidi, F.; Merzouki, M.; Benlemlih, M. Identification of the yeasts isolated from the Olive Mill Wastewater and study of catalase activity. Biotechnol. Bioeng. 2002, 7, 23–30. [Google Scholar] [CrossRef]
- Babavalian, H.; Ali Amoozegar, M.; Zahraei, S.; Rohban, R.; Shakeri, F.; Mehrdad Moosazadeh, M. Comparison of Bacterial Biodiversity and Enzyme Production in Three Hypersaline Lakes; Urmia, Howz-Soltan and Aran-Bidgol. Indian J. Microbiol. 2014, 54, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.N.; Khan, M.M. Screening, Identification and Characterization of Alcohol Tolerant Potential Bioethanol Producing yeasts. Curr. Res. Microbiol. Biotechnol. 2016, 2, 316–324. [Google Scholar]
- Milala, M.; Shugaba, A.; Gidado, A.; Ene, A.; Wafar, J. Studies on the Use of Agricultural Wastes for Cellulase Enzyme Production by Aspegillus Niger. Res. J. Agric. Biol. Sci. 2005, 1, 325–328. [Google Scholar]
- Arroyo, F.N.; Durán-Quintana, M.C.; Garrido-Fernández, A. Evaluation of primary models to describe the growth of Pichia anomala and study by response surface methodology of the effects of temperature, NaCl and pH on its biological parameters. J. Food Prot. 2005, 68, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.; Libkind, D.; Sampaio, J.P.; Van Broock, M.R. Yeast Diversity in the Acidic Rio Agrio-Lake Caviahue Volcanic Environment (Patagonia, Argentina). FEMS Microb. Ecol. 2008, 65, 415–424. [Google Scholar] [CrossRef]










| Isolation Medium | Sampling Period | Total Counts of Yeasts (CFU/g) | Number of Isolates |
|---|---|---|---|
| Isolates from GGS | 2015 | 7.9 × 109 | 24 |
| 2016 | 6.52 × 109 | 31 | |
| Isolates from DR | 2015 | 16.8 × 106 | 19 |
| 2016 | 11 × 106 | 22 |
| Isolation Medium | Positive Isolates | Total Number of Isolates |
|---|---|---|
| Isolates from GGS | 22 (40%) | 55 |
| Isolates from DR | 17 (41.5%) | 41 |
| Enzyme | Current Study | 1 | 2 | 3 | 4 | 5 | |
|---|---|---|---|---|---|---|---|
| a | b | ||||||
| Cellulase | 30 | 24 | - | 11 | - | 38 | - |
| Pectinase | 24 | 12 | - | 9 | 62 | 4 | 23 |
| Lipase | 26 | 23 | 4 | - | 134 | - | 6 |
| β-glucosidase | 28 | 24 | 10 | - | 142 | 13 | - |
| Catalase | 29 | 24 | 16 | - | - | - | 41 |
| Inulinase | 19 | 7 | - | - | - | - | - |
| Urease | 15 | 16 | - | - | - | - | - |
| Gelatinase | 17 | 1 | - | - | - | - | - |
| Protease | 26 | 18 | 6 | 10 | 49 | - | - |
| Amylase | 0 | 1 | - | 9 | - | - | - |
| Tannase | 0 | 0 | - | - | - | - | - |
| Number of isolates | 30 | 24 | 30 | 216 | 42 | 83 | |
| Glucose Concentration (g/L) | NaCl Concentration (g/L) | ||||||
|---|---|---|---|---|---|---|---|
| Concentration | 50 | 100 | 150 | 200 | 250 | 300 | 100 |
| Isolates from (GGS) (%) | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Isolates from (DR) (%) | 100 | 100 | 100 | 100 | 100 | 100 | 95.83 |
| Coded | Biotope | Homologous Strains | Accession Numbers |
|---|---|---|---|
| Mor-L1 | GGS | Trichosporon insectorum | ON810624.1 |
| Mor-L2a | GGS | Trichosporon insectorum | ON810627.1 |
| Mor-L3b | GGS | Trichosporon insectorum | ON810628.1 |
| Mor-L4a | GGS | Trichosporon japonicum | ON810629.1 |
| Mor-L5 | GGS | Trichosporon insectorum | ON810630.1 |
| Mor-L5a | GGS | Trichosporon insectorum | ON862731.1 |
| Mor-L11a | GGS | Trichosporon insectorum | ON810622.1 |
| Mor-L11 (1) | GGS | Trichosporon insectorum | ON862728.1 |
| Mor-L11 (2) | GGS | Trichosporon insectorum | ON862729.1 |
| Mor-L11C | GGS | Trichosporon insectorum | ON810621.1 |
| Mor-L14 | GGS | Trichosporon insectorum | ON810623.1 |
| Mor-L14 (1) | GGS | Trichosporon insectorum | ON862731.1 |
| Mor-L14 (3) | GGS | Trichosporon insectorum | ON862732.1 |
| Mor-L23b | GGS | Trichosporon insectorum | ON810625.1 |
| Mor-L23C | GGS | Trichosporon insectorum | ON810626.1 |
| Mor-L30a | GGS | Trichosporon coremiiforme | ON810631.1 |
| Mor-B13 | DR | Trichosporon insectorum | ON810633.1 |
| Mor-B25 | DR | Trichosporon insectorum | ON810632.1 |
| Mor-B13a | DR | Trichosporon insectorum | ON862726.1 |
| Mor-B25a | DR | Trichosporon insectorum | ON862727.1 |
| Mor-B5 | DR | Trichosporon insectorum | ON862730.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanane, T.; Najoua, B.; Salsabil, H.; Abdellatif, J.I.; Dalila, B.; Ahmad, I.; BukharI, S.A.R.; Irfan, M.; Chen, L.; Hicham, B. Qualitative Screening of Yeast Biodiversity for Hydrolytic Enzymes Isolated from the Gastrointestinal Tract of a Coprophage “Gymnopleurus sturmi” and Dung of Ruminants. Fermentation 2022, 8, 692. https://doi.org/10.3390/fermentation8120692
Hanane T, Najoua B, Salsabil H, Abdellatif JI, Dalila B, Ahmad I, BukharI SAR, Irfan M, Chen L, Hicham B. Qualitative Screening of Yeast Biodiversity for Hydrolytic Enzymes Isolated from the Gastrointestinal Tract of a Coprophage “Gymnopleurus sturmi” and Dung of Ruminants. Fermentation. 2022; 8(12):692. https://doi.org/10.3390/fermentation8120692
Chicago/Turabian StyleHanane, Touijer, Benchemsi Najoua, Hamdi Salsabil, Janati Idrissi Abdellatif, Bousta Dalila, Irfan Ahmad, Sayyad Ali Raza BukharI, Muhammad Irfan, Lijing Chen, and Bekkari Hicham. 2022. "Qualitative Screening of Yeast Biodiversity for Hydrolytic Enzymes Isolated from the Gastrointestinal Tract of a Coprophage “Gymnopleurus sturmi” and Dung of Ruminants" Fermentation 8, no. 12: 692. https://doi.org/10.3390/fermentation8120692
APA StyleHanane, T., Najoua, B., Salsabil, H., Abdellatif, J. I., Dalila, B., Ahmad, I., BukharI, S. A. R., Irfan, M., Chen, L., & Hicham, B. (2022). Qualitative Screening of Yeast Biodiversity for Hydrolytic Enzymes Isolated from the Gastrointestinal Tract of a Coprophage “Gymnopleurus sturmi” and Dung of Ruminants. Fermentation, 8(12), 692. https://doi.org/10.3390/fermentation8120692