
Analysis of the Microbial Community Structure and Volatile Metabolites of JIUYAO in Fangxian, China
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. DNA Extraction, PCR Amplification, and Illumina MiSeq Sequencing
2.3. Determination of Volatile Flavor Compounds
2.4. Statistical and Bioinformatic Analysis
3. Results
3.1. Sequencing the JIUYAO Samples
3.2. α-Diversity Analysis of the JIUYAO Samples
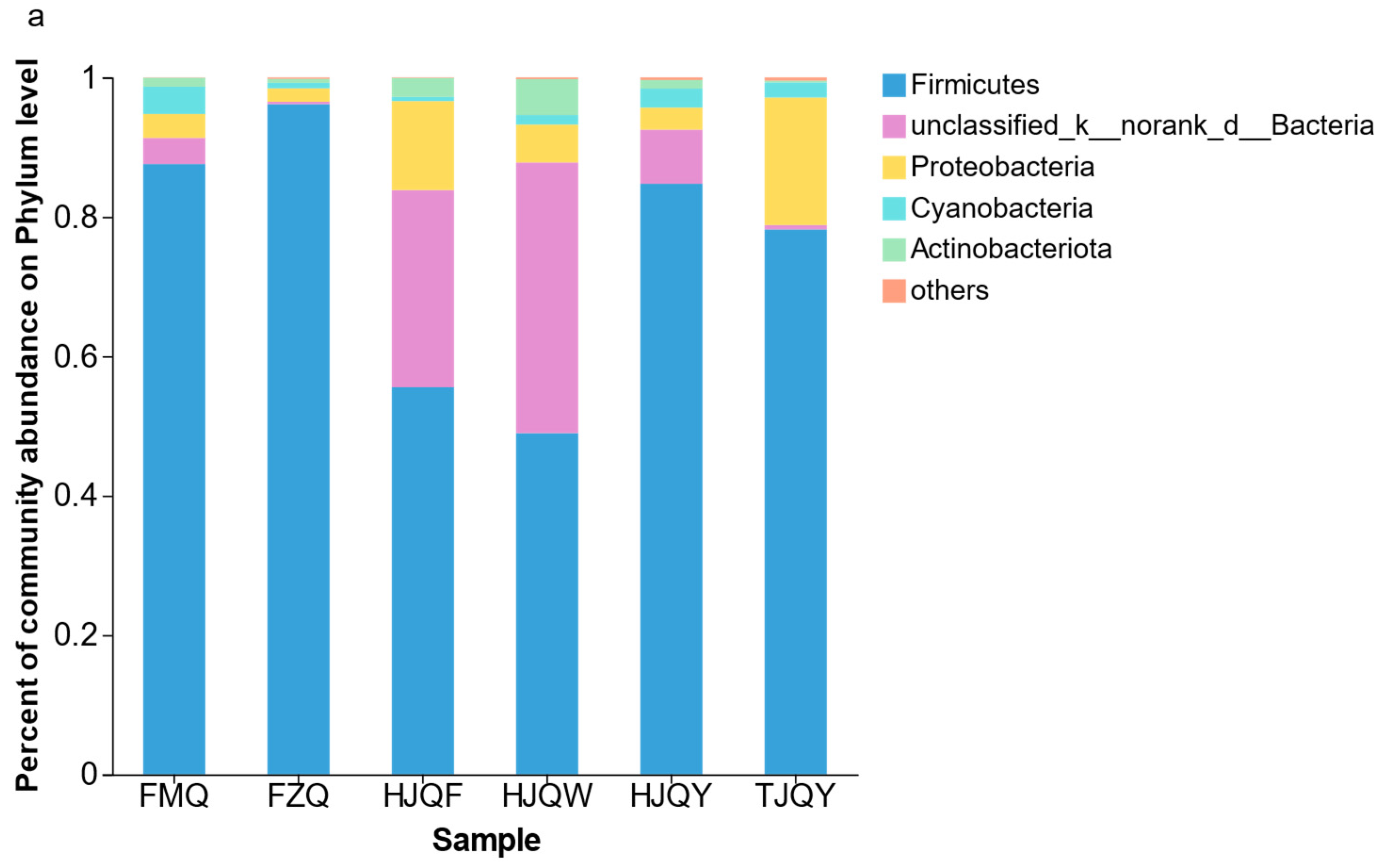
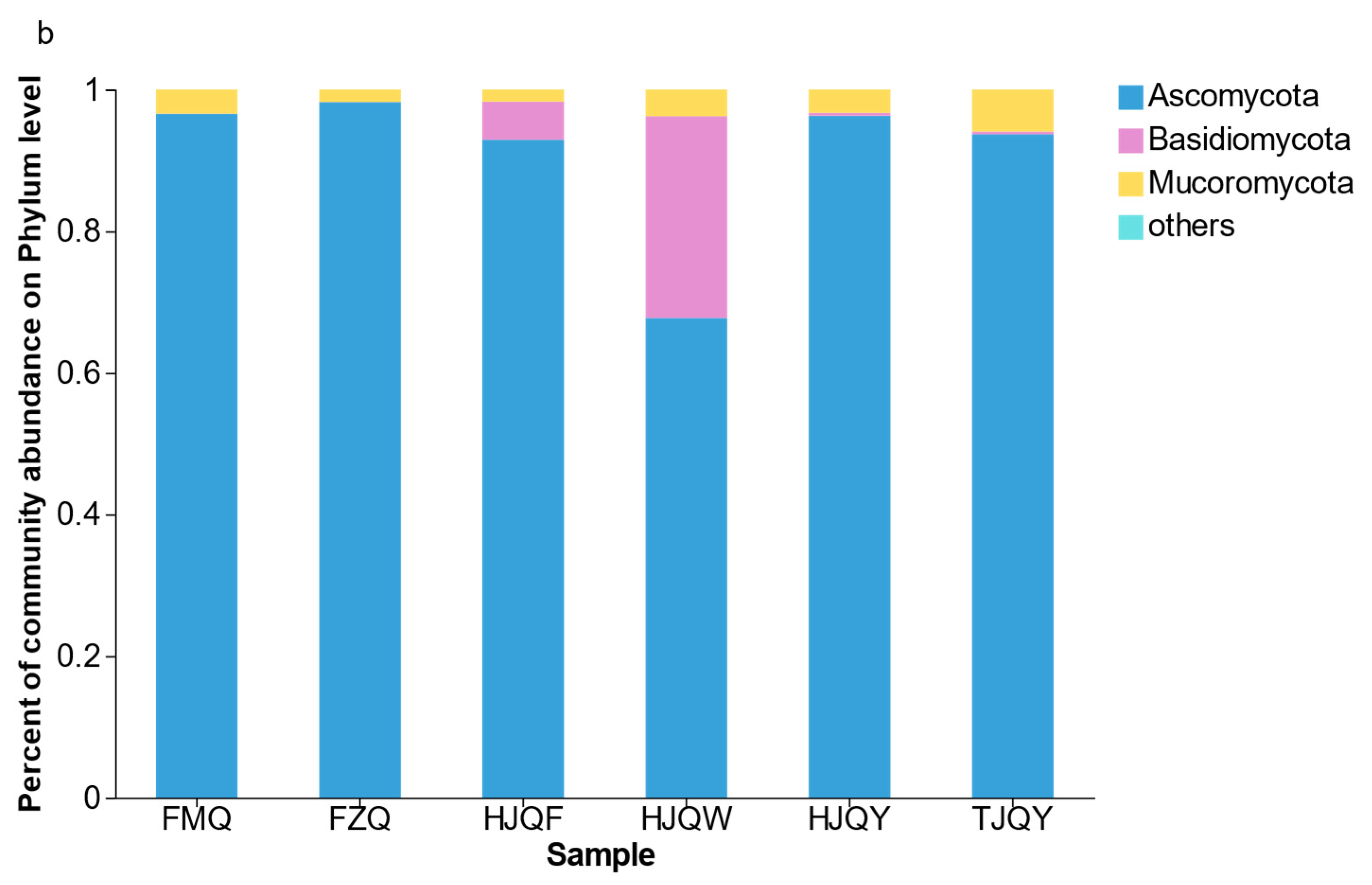
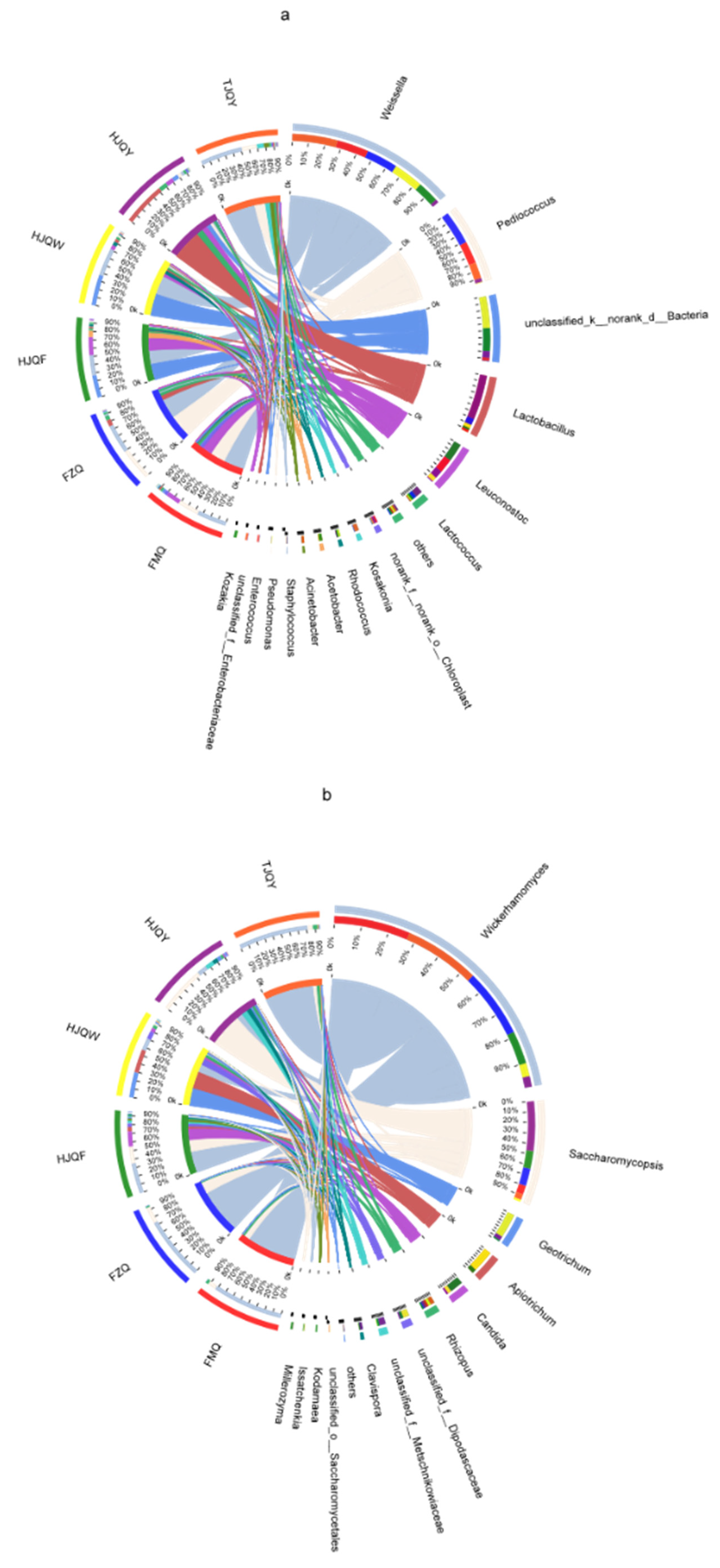
3.3. Microbial Composition Analysis in JIUYAO
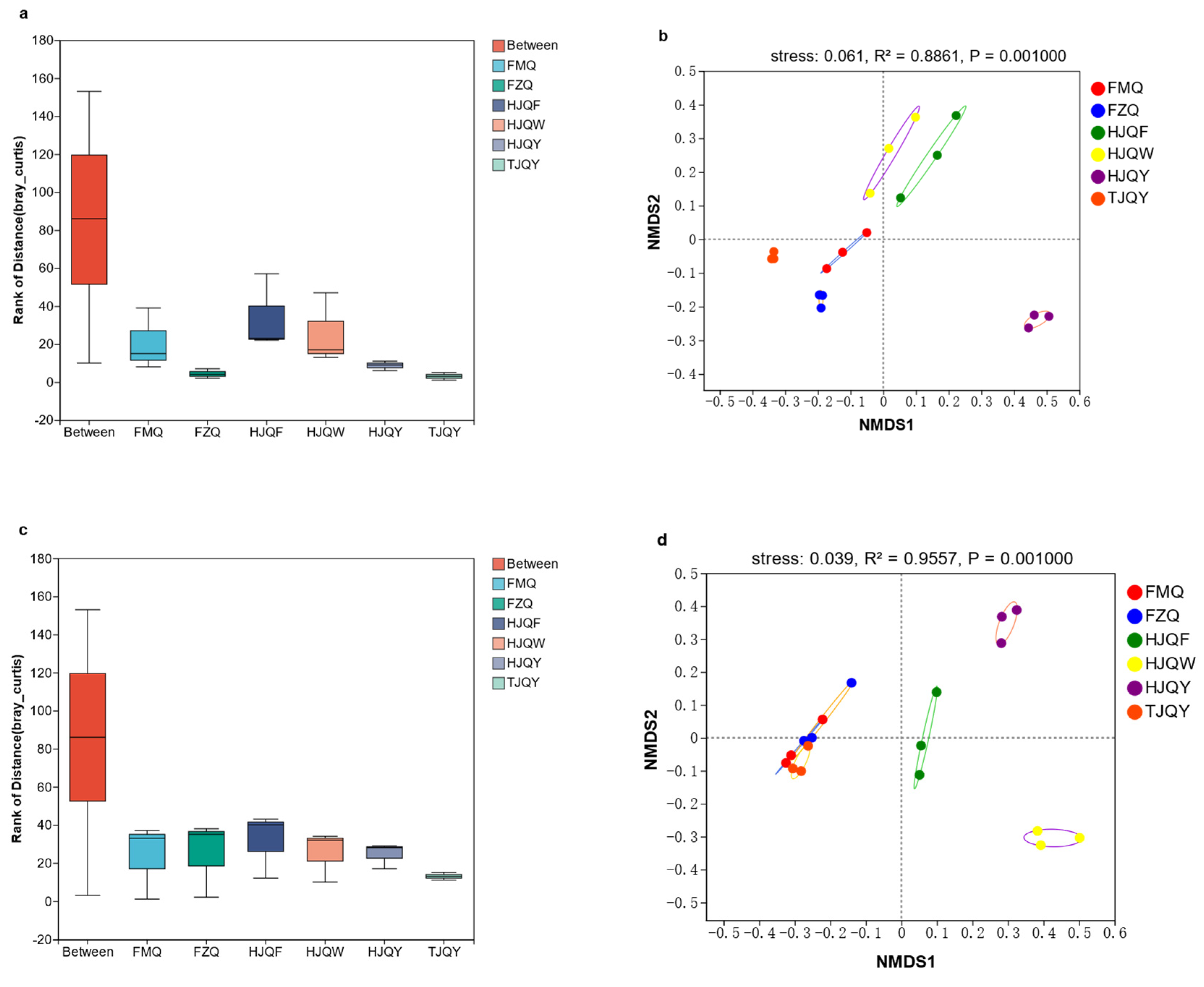
3.4. β-Diversity Analysis of JIUYAO
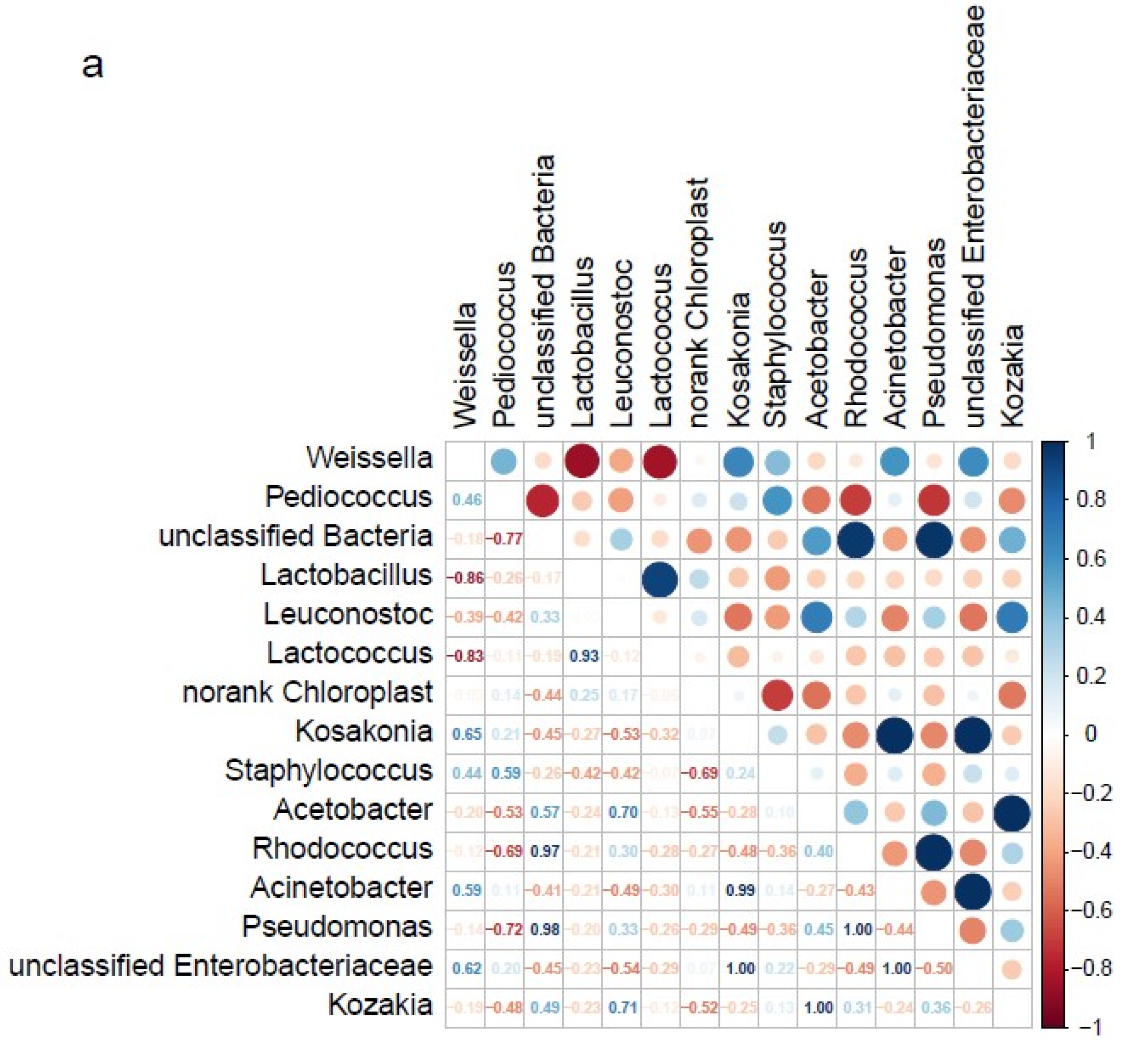
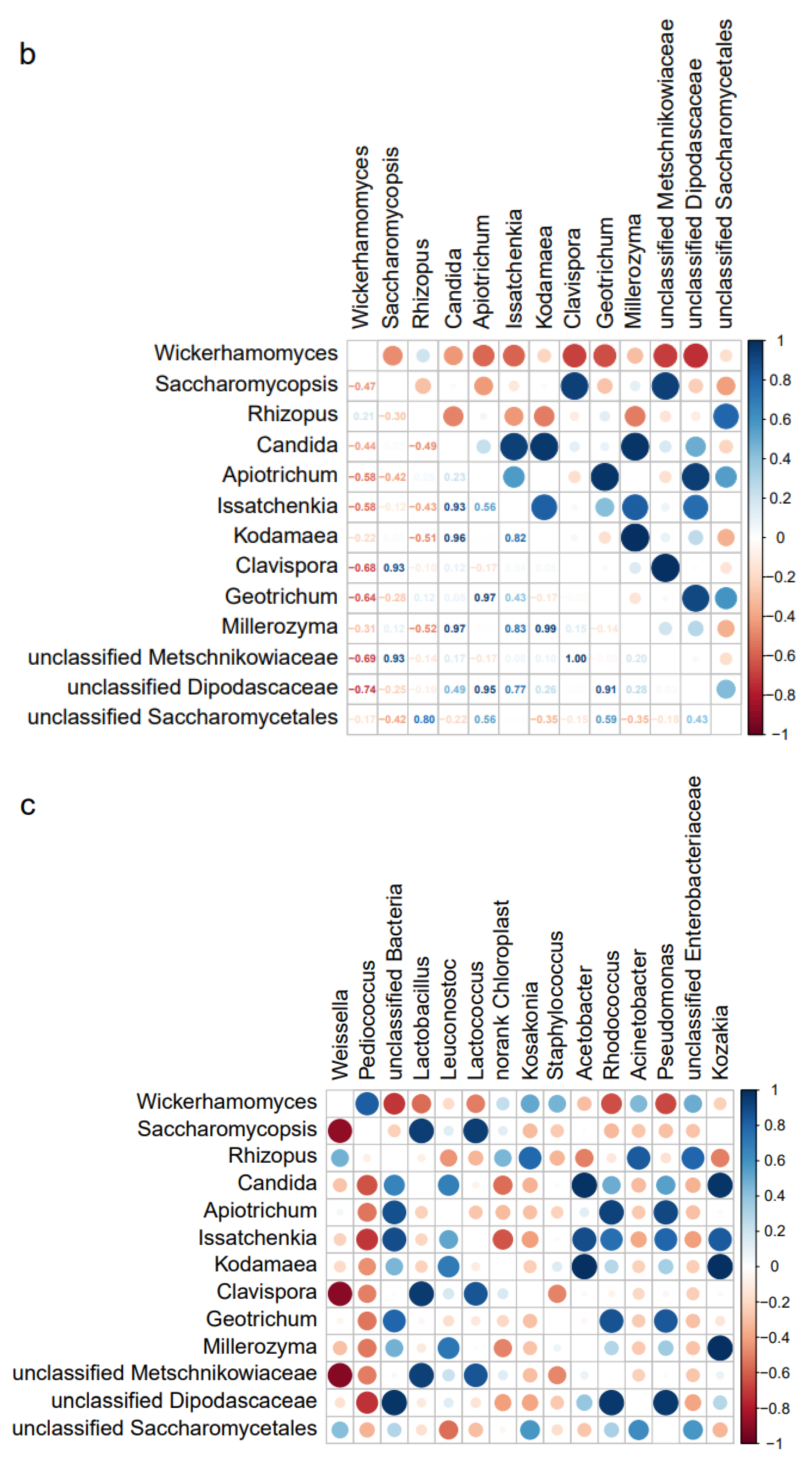
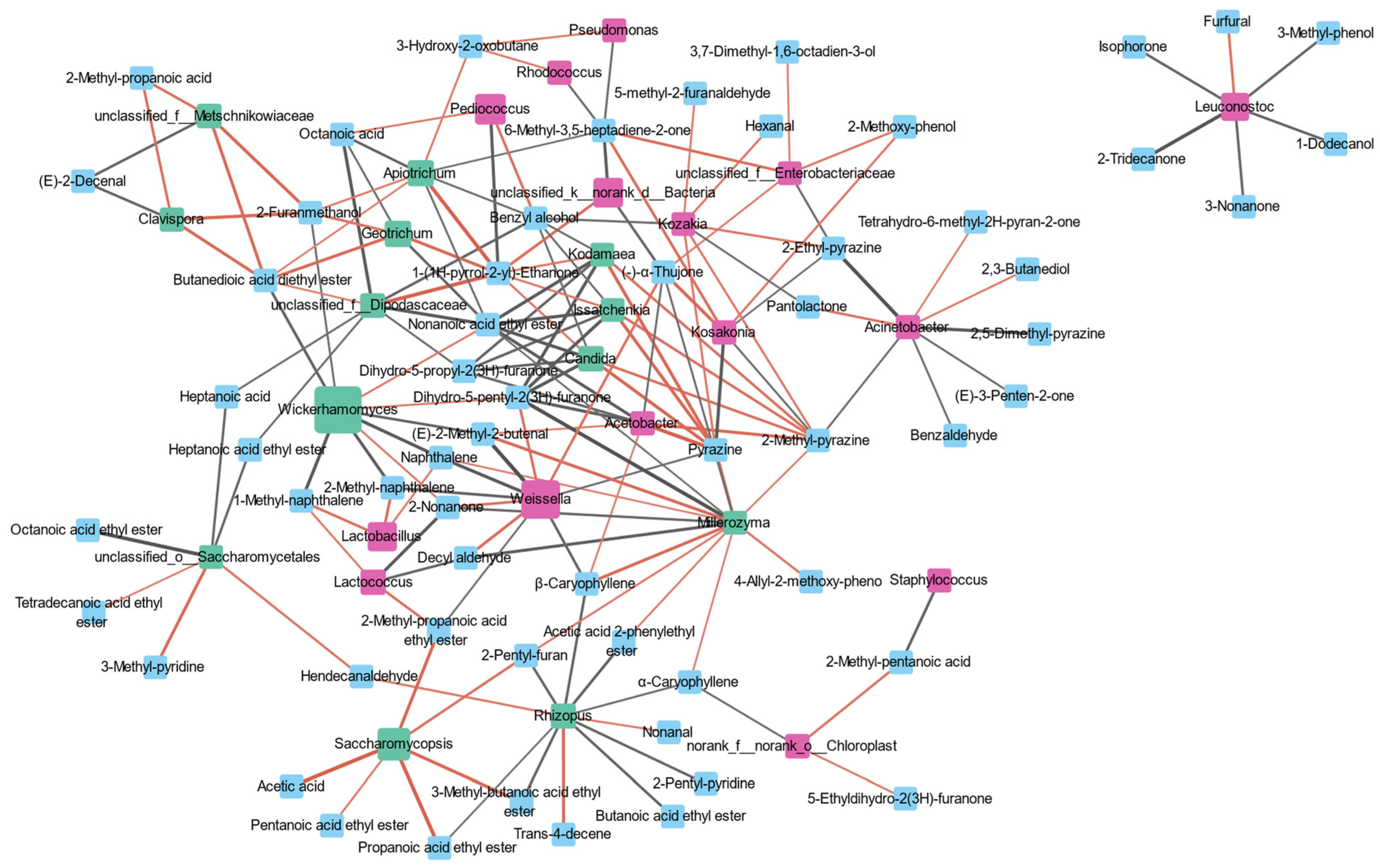
3.5. Analysis of Microbe Interactions in JIUYAO
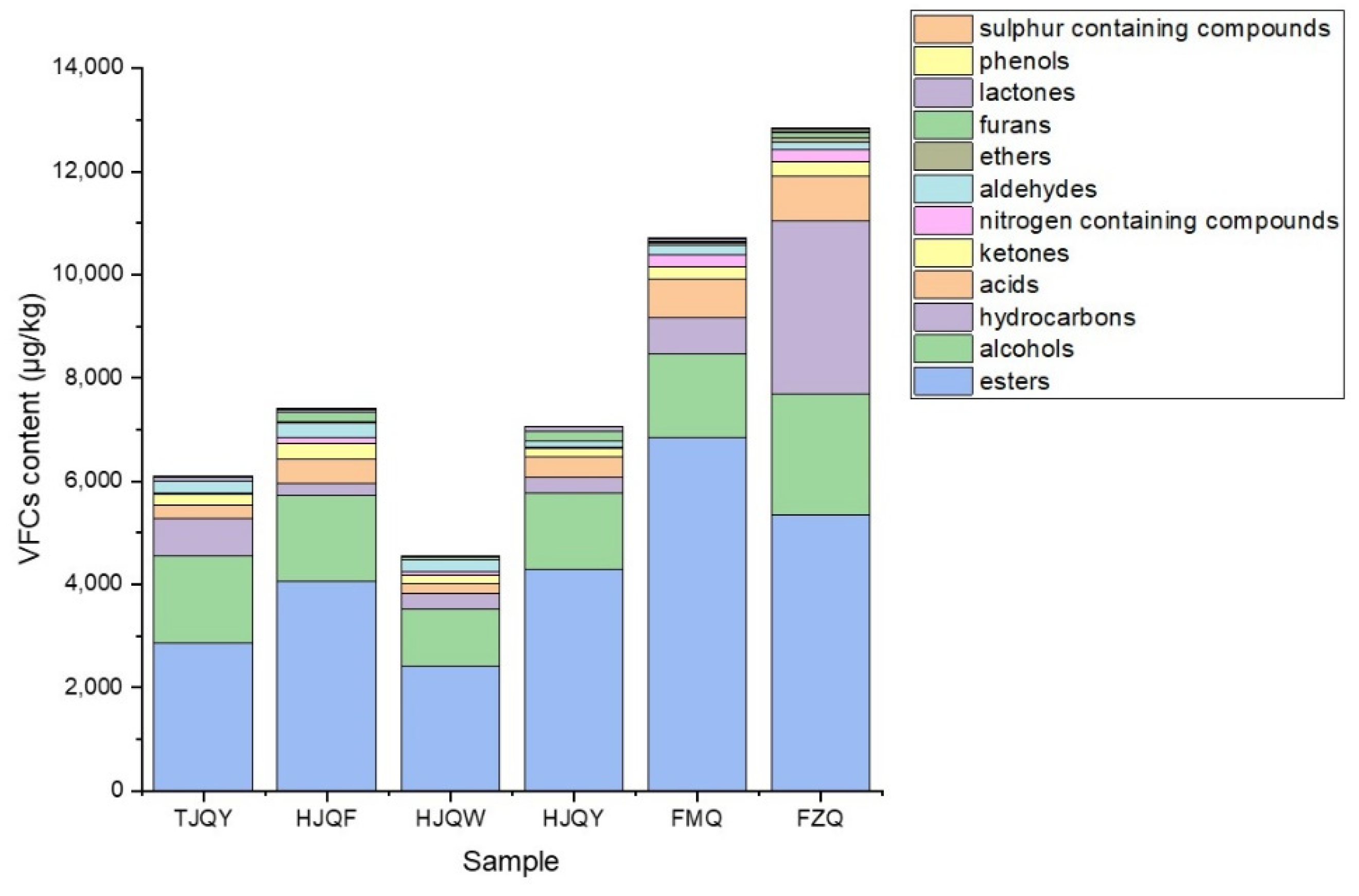
3.6. Analysis of Volatile Flavor Compounds in JIUYAO
3.7. Correlation between Micro-Organisms and Volatile Flavor Compounds
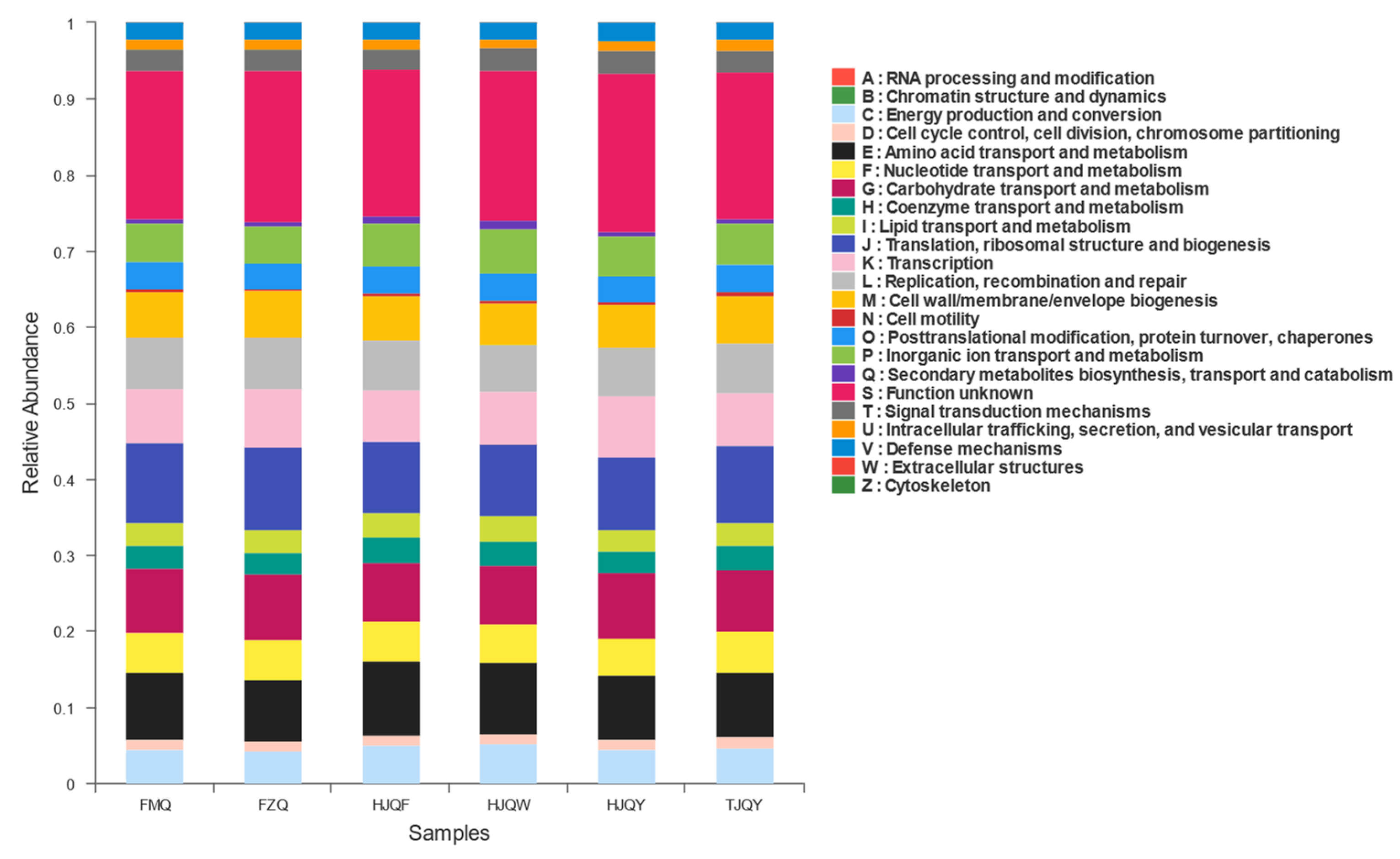
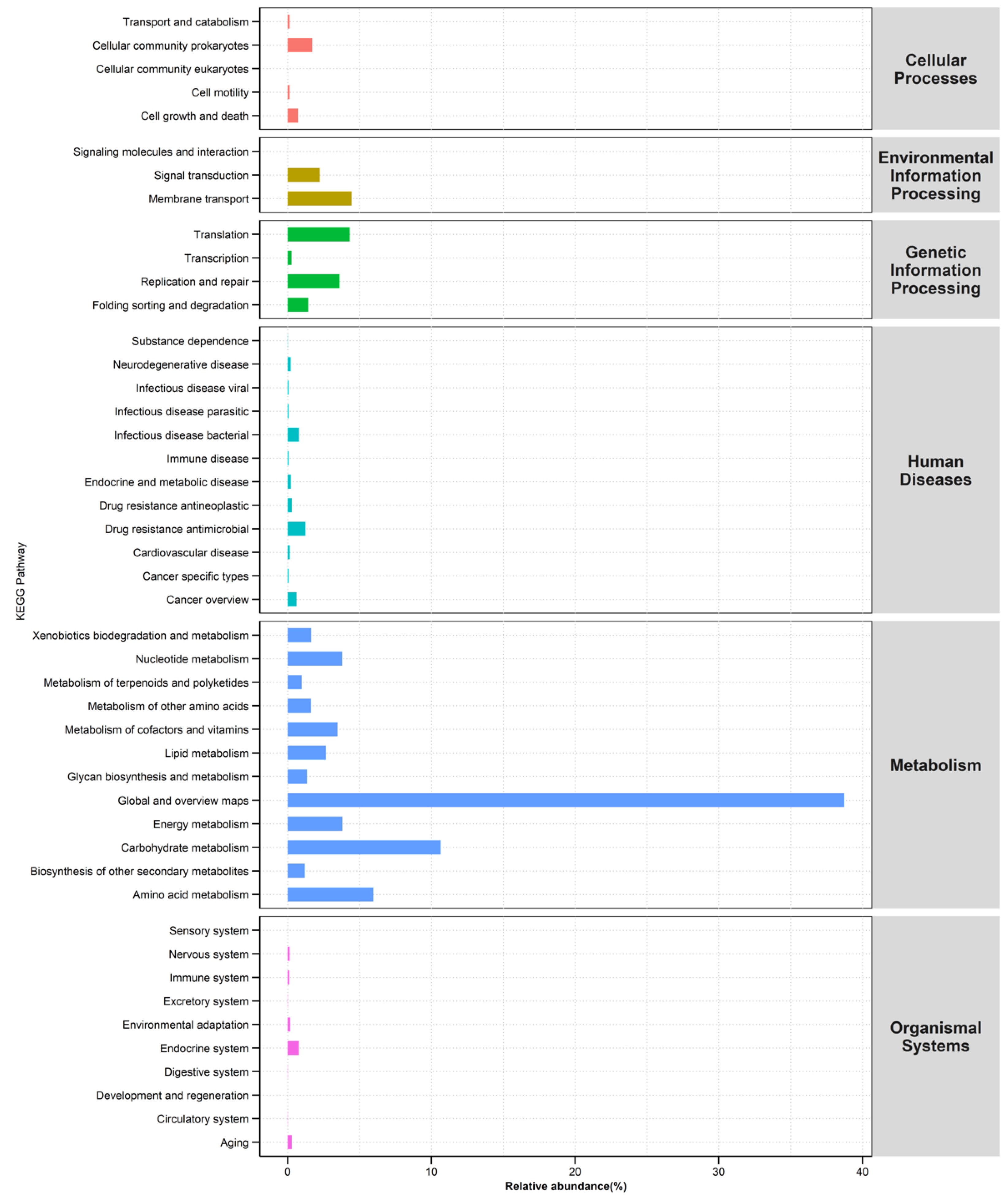
3.8. Functional Microbial Metabolic Pathways
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, C.; Liu, Y.; Tian, H.; Ai, L.; Yu, H. Metagenomic analysis reveals the impact of JIUYAO microbial diversity on fermentation and the volatile profile of Shaoxing-jiu. Food Microbiol. 2020, 86, 103326. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xiang, F.; Wang, L.; Zhang, X.; Zhang, J. Effect of Polygonum hydropiper on Yeast and Its Application in the Production of Rice Wine Starter. Food Res. Dev. 2017, 38, 117–120. [Google Scholar]
- Shen, B. Preliminary study on the role of Polygonum spicum in traditional Shaoxing liquor. Huaxia Wine News, 7 May 2010; p. 3. [Google Scholar]
- Lu, B.; Li, X. Study on the Effects of Polygonum hydropiper on Xiaoqu Quality. Liquor-Mak. Sci. Technol. 2006, 11, 42–43. [Google Scholar]
- Yang, Y.; Hu, W.; Xia, Y.; Mu, Z.; Tao, L.; Song, X.; Zhang, H.; Ni, B.; Ai, L. Flavor Formation in Chinese Rice Wine (Huangjiu): Impacts of the Flavor-Active Microorganisms, Raw Materials, and Fermentation Technology. Front. Microbiol. 2020, 11, 580247. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xia, Y.; Wang, G.; Yu, J.; Ai, L. Effect of mixed yeast starter on volatile flavor compounds in Chinese rice wine during different brewing stages. LWT 2017, 78, 373–381. [Google Scholar] [CrossRef]
- Liu, S.; Hu, J.; Xu, Y.; Xue, J.; Zhou, J.; Han, X.; Ji, Z.; Mao, J. Combined use of single molecule real-time DNA sequencing technology and culture-dependent methods to analyze the functional microorganisms in inoculated raw wheat Qu. Food Res. Int. 2020, 132, 109062. [Google Scholar] [CrossRef]
- Yu, P.; Du, J.; Cao, C.; Cai, G.; Sun, J.; Wu, D.; Lu, J. Development of a novel multi-strain wheat Qu with high enzyme activities for Huangjiu fermentation. J. Sci. Food Agric. 2021, 101, 4808–4817. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, C.; Gao, X.; Kang, Y.; Huang, M.; Wu, J.; Liu, Y.; Zhang, J.; Li, H.; Zhang, Y. Characterization of key aroma compounds in Huangjiu from northern China by sensory-directed flavor analysis. Food Res. Int. 2020, 134, 109238. [Google Scholar] [CrossRef]
- Chen, L.; Ren, L.; Li, D.; Ma, X. Analysis of microbiomes in three traditional starters and volatile components of the Chinese rice wines. Food Sci. Biotechnol. 2021, 30, 87–96. [Google Scholar] [CrossRef]
- Wang, Z.M.; Wang, C.T.; Shen, C.H.; Wang, S.T.; Mao, J.Q.; Li, Z.; Gänzle, M.; Mao, J. Microbiota stratification and succession of amylase-producing Bacillus in traditional Chinese Jiuqu (fermentation starters). J. Sci. Food Agric. 2020, 100, 3544–3553. [Google Scholar] [CrossRef]
- Wang, P.; Mao, J.; Meng, X.; Li, X.; Liu, Y.; Feng, H. Changes in flavour characteristics and bacterial diversity during the traditional fermentation of Chinese rice wines from Shaoxing region. Food Control 2014, 44, 58–63. [Google Scholar] [CrossRef]
- Huang, X.; Fan, Y.; Meng, J.; Sun, S.; Wang, X.; Chen, J.; Han, B.-Z. Laboratory-scale fermentation and multidimensional screening of lactic acid bacteria from Daqu. Food Biosci. 2021, 40, 100853. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, T.; Zhang, Q.; Luo, J.; Cai, C.; Mao, J. Microbial diversity and chemical analysis of the starters used in traditional Chinese sweet rice wine. Food Microbiol. 2018, 73, 319–326. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, S.; Sun, H.; Jiang, Z.; Xu, Y.; Mao, J.; Qian, B.; Wang, L.; Mao, J. Metagenomics-based insights into the microbial community profiling and flavor development potentiality of baijiu Daqu and huangjiu wheat Qu. Food Res. Int. 2022, 152, 110707. [Google Scholar] [CrossRef]
- Ren, Q.; Sun, L.; Wu, H.; Wang, Y.; Wang, Z.; Zheng, F.; Lu, X.; Xu, J. The changes of microbial community and flavor compound in the fermentation process of Chinese rice wine using Fagopyrum tataricum grain as feedstock. Sci. Rep. 2019, 9, 3365. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.-R.; Guo, W.-L.; Zhou, W.-B.; Li, L.; Xu, J.-X.; Hong, J.-L.; Liu, H.-P.; Zeng, F.; Bai, W.-D.; Liu, B.; et al. Microbial communities and volatile metabolites in different traditional fermentation starters used for Hong Qu glutinous rice wine. Food Res. Int. 2019, 121, 593–603. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, L.; Zhan, P.; Tian, H.; Liu, J. Characterization of the aroma compounds of Millet Huangjiu at different fermentation stages. Food Chem. 2022, 366, 130691. [Google Scholar] [CrossRef]
- Wang, J.; Yi, X.; Huang, M.; Liu, Y.; Meng, W.; Yang, Q.; Huo, J.; Zhang, Q.; Luo, X.; Wu, J.; et al. Studies on the key odorants in Maopu buckwheat finished Baijiu and the effect of tartary buckwheat extract on its flavor. LWT 2022, 154, 112650. [Google Scholar] [CrossRef]
- Yang, Y.; Ai, L.; Mu, Z.; Liu, H.; Yan, X.; Ni, L.; Zhang, H.; Xia, Y. Flavor compounds with high odor activity values (OAV > 1) dominate the aroma of aged Chinese rice wine (Huangjiu) by molecular association. Food Chem. 2022, 383, 132370. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Z.; Sun, J.; Ni, L. The dynamics of volatile compounds and their correlation with the microbial succession during the traditional solid-state fermentation of Gutian Hong Qu glutinous rice wine. Food Microbiol. 2020, 86, 103347. [Google Scholar] [CrossRef]
- Hu, Y.; Lei, X.; Zhang, X.; Guan, T.; Wang, L.; Zhang, Z.; Yu, X.; Tu, J.; Peng, N.; Liang, Y.; et al. Characteristics of the Microbial Community in the Production of Chinese Rice-Flavor Baijiu and Comparisons with the Microflora of Other Flavors of Baijiu. Front. Microbiol. 2021, 12, 673670. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Su, W.; Mu, Y.; Jiang, L.; Mu, Y. Correlations between microbiota with physicochemical properties and volatile flavor components in black glutinous rice wine fermentation. Food Res. Int. 2020, 138, 109800. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.-N.; Chen, C.; Huang, X.-N.; Yan, Y.-Z.; Chen, J.-Y.; Han, B.-Z. Influence of indigenous lactic acid bacteria on the volatile flavor profile of light-flavor Baijiu. LWT 2021, 147, 111540. [Google Scholar] [CrossRef]
- Zhou, W.; Shu, Q.; Zhang, X.; Chen, Q. Application of mixed-culture with Lactobacillus brevis and Saccharomyces cerevisiae to Chinese yellow rice wine brewing for ethyl carbamate regulation. Food Control 2021, 122, 107821. [Google Scholar] [CrossRef]
- Fan, G.; Teng, C.; Xu, D.; Fu, Z.; Minhazul, K.A.; Wu, Q.; Liu, P.; Yang, R.; Li, X. Enhanced production of ethyl acetate using co-culture of Wickerhamomyces anomalus and Saccharomyces cerevisiae. J. Biosci. Bioeng. 2019, 128, 564–570. [Google Scholar] [CrossRef]
- Yan, S.; Xiangsong, C.; Xiang, X. Improvement of the aroma of lily rice wine by using aroma-producing yeast strain Wickerhamomyces anomalus HN006. AMB Express 2019, 9, 89. [Google Scholar] [CrossRef]
- Wu, J.; He, W.; Wang, J.; Yang, Y.; Lan, C.; Liu, S.; Yang, T. Optimization of preparation technology of flavor rice starter by response surface methodology. China Brew. 2020, 39, 120–124. [Google Scholar] [CrossRef]
- Xiang, W.; Xu, Q.; Zhang, N.; Rao, Y.; Zhu, L.; Zhang, Q. Mucor indicus and Rhizopus oryzae co-culture to improve the flavor of Chinese turbid rice wine. J. Sci. Food Agric. 2019, 99, 5577–5585. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Q.; Xu, Y.; Sun, B. Specific Volumetric Weight-Driven Shift in Microbiota Compositions With Saccharifying Activity Change in Starter for Chinese Baijiu Fermentation. Front. Microbiol. 2018, 9, 2349. [Google Scholar] [CrossRef] [Green Version]
- Ling, M. Study on the Key Indicators Screening and Quality Evaluation Method of Shaoxing Huangjiu Wheat Qu. Master’s Thesis, Jiangnan University, Wuxi, China, 2019; p. 73.
- Li, W.; Cui, D.-Y.; Wang, J.-H.; Liu, X.-E.; Xu, J.; Zhou, Z.; Zhang, C.-Y.; Chen, Y.-F.; Xiao, D.-G. Overexpression of different alcohol acetyltransferase genes with BAT2 deletion in Saccharomyces cerevisiae affects acetate esters and higher alcohols. Eur. Food Res. Technol. 2018, 244, 555–564. [Google Scholar] [CrossRef]
- Kruis, A.J.; Levisson, M.; Mars, A.E.; van der Ploeg, M.; Daza, F.G.; Ellena, V.; Kengen, S.W.; van der Oost, J.; Weusthuis, R.A. Ethyl acetate production by the elusive alcohol acetyltransferase from yeast. Metab. Eng. 2017, 41, 92–101. [Google Scholar] [CrossRef]
- Van Laere, S.D.M.; Saerens, S.M.G.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Flavour formation in fungi: Characterisation of KlAtf, the Kluyveromyces lactis orthologue of the Saccharomyces cerevisiae alcohol acetyltransferases Atf1 and Atf2. Appl. Microbiol. Biotechnol. 2008, 78, 783–792. [Google Scholar] [CrossRef]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Hou, H.; Lu, X.; Chen, X.; Fang, D.; Hu, Q.; Zhao, L. Production of an innovative mixed Qu (fermentation starter) for waxy maize brewing and comparison of the quality of different waxy maize wines. J. Sci. Food Agric. 2021, 101, 2328–2336. [Google Scholar] [CrossRef]
- Mo, X.; Xu, Y.; Fan, W. Characterization of Aroma Compounds in Chinese Rice Wine Qu by Solvent-Assisted Flavor Evaporation and Headspace Solid-Phase Microextraction. J. Agric. Food Chem. 2010, 58, 2462–2469. [Google Scholar] [CrossRef]
- Yang, Z.; Sakai, M.; Sayama, H.; Shimeno, T.; Yamaguchi, K.; Watanabe, N. Elucidation of the biochemical pathway of 2-phenylethanol from shikimic acid using isolated protoplasts of rose flowers. J. Plant Physiol. 2009, 166, 887–891. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Bai, M.; Zhou, J.; Jin, Z.; Xu, Y.; Yang, Q.; Zhou, J.; Zhang, S.; Mao, J. Analysis of genes from Saccharomyces cerevisiae HJ01 participating in aromatic alcohols biosynthesis during huangjiu fermentation. LWT 2022, 154, 112705. [Google Scholar] [CrossRef]
- Bai, M. The Characteristics of Prephenate Dehydratase for Chinese Huangjiu Yeast in 2-Phenylethanol Synthetic Pathway. Master’s Thesis, Jiangnan University, Wuxi, China, 2018. [Google Scholar]
- Chen, S.; Xu, Y.; Qian, M.C. Aroma Characterization of Chinese Rice Wine by Gas Chromatography–Olfactometry, Chemical Quantitative Analysis, and Aroma Reconstitution. J. Agric. Food Chem. 2013, 61, 11295–11302. [Google Scholar] [CrossRef]
- Yan, Y.; Sun, L.; Xing, X.; Wu, H.; Lu, X.; Zhang, W.; Xu, J.; Ren, Q. Microbial succession and exploration of higher alcohols-producing core bacteria in northern Huangjiu fermentation. AMB Express 2022, 12, 79. [Google Scholar] [CrossRef]
- Son, E.Y.; Lee, S.M.; Kim, M.; Seo, J.A.; Kim, Y.S. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef]
- Park, M.K.; Kim, Y.-S. Distinctive Formation of Volatile Compounds in Fermented Rice Inoculated by Different Molds, Yeasts, and Lactic Acid Bacteria. Molecules 2019, 24, 2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Chen, S.; Zhou, Z. Characterization of volatile organic compounds as potential aging markers in Chinese rice wine using multivariable statistics. J. Sci. Food Agric. 2019, 99, 6444–6454. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, C.; Qian, M.; Li, Z.; Xu, Y. Characterization of the Key Aroma Compounds in Aged Chinese Rice Wine by Comparative Aroma Extract Dilution Analysis, Quantitative Measurements, Aroma Recombination, and Omission Studies. J. Agric. Food Chem. 2019, 67, 4876–4884. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, B.; Wu, Q.; Jiang, X.Y.; Liu, H.J.; Wang, C.Z.; Huang, M.Q.; Wu, J.H.; Zhang, J.L.; Yu, Y.G. Sensomics-assisted flavor decoding of coarse cereal Huangjiu. Food Chem. 2022, 381, 132296. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Carrascón, V.; Ferreira, V. Release and Formation of Oxidation-Related Aldehydes during Wine Oxidation. J. Agric. Food Chem. 2016, 64, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, Y.; Gao, X.; Jiang, X.; Huang, M.; Ye, H.; Wu, J.; Zhang, J.; Sun, X.; Wu, Q. Succession patterns of aroma components during brewing process of broomcorn millet (Panicum miliaceum L.) Huangjiu. Food Res. Int. 2022, 154, 110982. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Hussain, R.; Khan, Q.F.; Zhang, H. Functional microbiota in Chinese traditional Baijiu and Mijiu Qu (starters): A review. Food Res. Int. 2020, 138, 109830. [Google Scholar] [CrossRef]
- Van Lancker, F.; Adams, A.; De Kimpe, N. Impact of the N-Terminal Amino Acid on the Formation of Pyrazines from Peptides in Maillard Model Systems. J. Agric. Food Chem. 2012, 60, 4697–4708. [Google Scholar] [CrossRef]
- Zhu, B.-F.; Xu, Y.; Fan, W.-L. High-yield fermentative preparation of tetramethylpyrazine by Bacillus sp. using an endogenous precursor approach. J. Ind. Microbiol. Biotechnol. 2010, 37, 179–186. [Google Scholar] [CrossRef]
- Wang, P.; Wu, Q.; Jiang, X.; Wang, Z.; Tang, J.; Xu, Y. Bacillus licheniformis affects the microbial community and metabolic profile in the spontaneous fermentation of Daqu starter for Chinese liquor making. Int. J. Food Microbiol. 2017, 250, 59–67. [Google Scholar] [CrossRef]
- Liu, Y.; Qian, X.; Xing, J.; Li, N.; Li, J.; Su, Q.; Chen, Y.; Zhang, B.; Zhu, B. Accurate Determination of 12 Lactones and 11 Volatile Phenols in Nongrape Wines through Head-space-Solid-Phase Microextraction (HS-SPME) Combined with High-Resolution Gas Chromatography-Orbitrap Mass Spec-trometry (GC-Orbitrap-MS). J. Agric. Food Chem. 2022, 70, 1971–1983. [Google Scholar] [CrossRef]
- Qian, X.; Lan, Y.; Han, S.; Liang, N.; Zhu, B.; Shi, Y.; Duan, C. Comprehensive investigation of lactones and furanones in ice wines and dry wines using gas chromatography-triple quadrupole mass spectrometry. Food Res. Int. 2020, 137, 109650. [Google Scholar] [CrossRef]
- Harris, V.; Ford, C.M.; Jiranek, V.; Grbin, P.R. Survey of enzyme activity responsible for phenolic off-flavour production by Dekkera and Brettanomyces yeast. Appl. Microbiol. Biotechnol. 2009, 81, 1117–1127. [Google Scholar] [CrossRef]
- Wen, D.; Li, C.; Di, H.; Liao, Y.; Liu, H. A Universal HPLC Method for the Determination of Phenolic Acids in Compound Herbal Medicines. J. Agric. Food Chem. 2005, 53, 6624–6629. [Google Scholar] [CrossRef]
- Wang, C.; Li, C.; Zhang, X.; Rao, J.; Xin, P.; Yin, J.; Song, Q. Determination of terpenoids in yam wine by liquid-liquid extraction coupled with gas chromatography-mass spectrometry. Food Ferment. Ind. 2019, 45, 213–217. [Google Scholar]
- Ma, L.; Gao, W.; Chen, F.; Meng, Q. HS-SPME and SDE combined with GC–MS and GC-O for characterization of flavor compounds in Zhizhonghe Wujiapi medicinal liquor. Food Res. Int. 2020, 137, 109590. [Google Scholar] [CrossRef]
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Zheng, H.; Wei, P.; Zhang, G.; Xu, W.; Li, Y. The impact of different Saccharomyces cerevisiae strains on microbial composition and quality of Chinese rice wine fermentations. Yeast 2021, 38, 147–156. [Google Scholar] [CrossRef]
- Chen, Q. Study on the Flavor Formation and Functional Microorganism of Shaoxing Mechanized Huangjiu. Master’s Thesis, Jiangnan University, Wuxi, China, 2018; p. 97. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Terpenoids | TJQY (μg/kg) | HJQF (μg/kg) | HJQW (μg/kg) | HJQY (μg/kg) | FMQ (μg/kg) | FZQ (μg/kg) |
|---|---|---|---|---|---|---|---|
| 1 | α-Pinene | - | - | - | - | - | 646.10 ± 32.50 a |
| 2 | Camphene | - | - | 0.89 ± 0.09 a | - | - | 1202.16 ± 143.23 b |
| 3 | 4-Methylene-1-(1-methylethyl)-bicyclo[3.1.0]hexane | 3.45 ± 0.47 b | - | 0.96 ± 0.11 ab | 2.20 ± 0.29 ab | - | 40.22 ± 3.93 c |
| 4 | 7-Methyl-3-methylene-1,6-octadiene | 26.90 ± 1.70 c | - | 0.67 ± 0.05 a | 5.94 ± 0.55 b | - | 66.83 ± 5.50 d |
| 5 | Limonene | 61.80 ± 2.70 b | - | - | - | 22.16 ± 0.64 a | 185.87 ± 15.53 c |
| 6 | γ-Terpinene | 1.87 ± 0.26 ab | 1.65 ± 0.17 ab | 2.29 ± 0.26 ab | 2.78 ± 0.42 b | - | 85.09 ± 3.38 c |
| 7 | 3,7-Dimethyl-1,3,6-octatriene | 0.27 ± 0.04 a | - | - | - | - | 3.50 ± 0.55 b |
| 8 | 1-Dethyl-4-(1-methylethylidene)-cyclohexene | 0.73 ± 0.06 ab | 0.93 ± 0.10 abc | 1.36 ± 0.16 bc | 1.87 ± 0.21 c | - | 11.28 ± 1.36 d |
| 9 | (-)-α-Thujone | 16.91 ± 0.90 c | - | - | - | 2.13 ± 0.25 a | 3.31 ± 0.06 b |
| 10 | Copaene | 9.56 ± 0.34 a | - | - | - | - | 34.06 ± 3.56 b |
| 11 | δ-Camphor | 5.30 ± 0.15 a | 14.01 ± 0.42 cd | 14.76 ± 1.08 d | 12.77 ± 1.04 c | 7.61 ± 0.10 b | 24.88 ± 1.09 e |
| 12 | 3,7-Dimethyl-1,6-octadien-3-ol | 6.53 ± 0.19 e | 2.45 ± 0.09c | 1.08 ± 0.07 a | 1.62 ± 0.18 b | 1.59 ± 0.06 b | 4.32 ± 0.46 d |
| 13 | Isophorone | 3.95 ± 0.28 c | - | 0.61 ± 0.02 a | - | - | 1.06 ± 0.07 b |
| 14 | β-Caryophyllene | 12.3 ± 0.93 a | 143.04 ± 4.95 d | 33.85 ± 3.42 b | 56.91 ± 3.27 c | 16.62 ± 0.33 a | 39.22 ± 3.25 b |
| 15 | Terpinen-4-ol | 3.22 ± 0.27 bc | 1.85 ± 0.10 ab | 4.77 ± 0.48 cd | 5.56 ± 0.49 d | 0.78 ± 0.15 a | 25.93 ± 2.14 e |
| 16 | Isoborneol | - | 0.17 ± 0.02 a | - | - | 0.55 ± 0.05 b | 2.94 ± 0.10 c |
| 17 | α-Caryophyllene | 1.03 ± 0.12 a | 5.73 ± 0.03 e | 1.84 ± 0.27 b | 3.14 ± 0.10 c | - | 3.55 ± 0.23 d |
| 18 | 2-Borneol | - | - | - | - | - | 156.93 ± 11.33 a |
| 19 | Carvone | 0.30 ± 0.02 a | - | - | - | - | 0.78 ± 0.09 b |
| 20 | 6,10-Dimethyl-5,9-undecadien-2-one | 2.83 ± 0.21 c | 1.28 ± 0.10 a | 1.16 ± 0.17 a | 1.40 ± 0.11 a | 1.69 ± 0.13 b | 1.21 ± 0.19 a |
| 21 | trans-β-Ionone | - | - | - | - | - | 0.41 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Ren, Q.; Wang, Z.; Liu, H.; Huang, M.; Wu, J.; Sun, B. Analysis of the Microbial Community Structure and Volatile Metabolites of JIUYAO in Fangxian, China. Fermentation 2022, 8, 754. https://doi.org/10.3390/fermentation8120754
Zhang W, Ren Q, Wang Z, Liu H, Huang M, Wu J, Sun B. Analysis of the Microbial Community Structure and Volatile Metabolites of JIUYAO in Fangxian, China. Fermentation. 2022; 8(12):754. https://doi.org/10.3390/fermentation8120754
Chicago/Turabian StyleZhang, Wendi, Qing Ren, Zhongchao Wang, Huijie Liu, Mingquan Huang, Jihong Wu, and Baoguo Sun. 2022. "Analysis of the Microbial Community Structure and Volatile Metabolites of JIUYAO in Fangxian, China" Fermentation 8, no. 12: 754. https://doi.org/10.3390/fermentation8120754
APA StyleZhang, W., Ren, Q., Wang, Z., Liu, H., Huang, M., Wu, J., & Sun, B. (2022). Analysis of the Microbial Community Structure and Volatile Metabolites of JIUYAO in Fangxian, China. Fermentation, 8(12), 754. https://doi.org/10.3390/fermentation8120754