
Single Cell Protein Production through Multi Food-Waste Substrate Fermentation

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate
2.2. Microorganism
2.3. Experimental Set-Up
2.4. Chemicals
2.5. Crude Protein, True Protein, Moisture, Ash and Lignin Determinations
2.6. Alcohol Insoluble Residues (AIR)
2.7. Sugar Analysis
2.8. Crude Fat and Fatty Acid Determination
2.9. Statistical Analysis
3. Results and Discussion
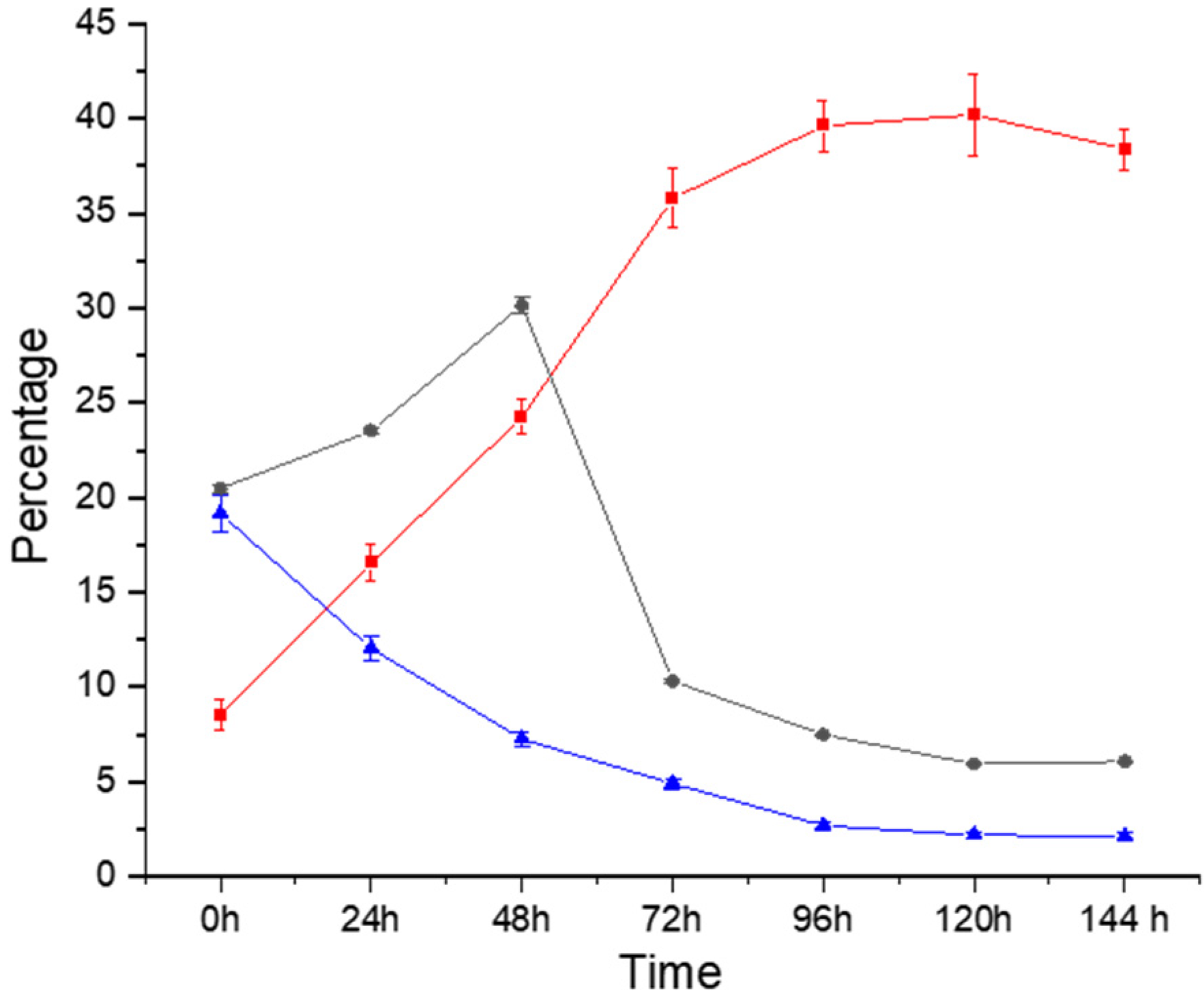
3.1. Proximate Composition
3.2. Fatty Acid Composition
3.3. Cell Wall Insoluble and Soluble Sugars
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Papargyropoulou, E.; Lozano, R.; Steinberger, J.K.; Wright, N.; bin Ujang, Z. The food waste hierarchy as a framework for the management of food surplus and food waste. J. Clean. Prod. 2014, 76, 106–115. [Google Scholar] [CrossRef]
- Elijah, A.I.; Edem, V.E. Value addition to Food and Agricultural wastes: A Biotechnological approach. Nig. J. Agric. Food Environ. 2017, 13, 139–154. [Google Scholar]
- United Nations Environment Programme. Food Waste Index Report; United Nations Environment Programme: Nairobi, Kenya, 2021; ISBN 978-92-807-3868-1. [Google Scholar]
- Aureli, V.; Scalvedi, M.L.; Rossi, L. FoodWaste of Italian Families: Proportion in Quantity and Monetary Value of Food Purchases. Foods 2021, 10, 1920. [Google Scholar] [CrossRef]
- Yazid, N.A.; Barrena, R.; Komilis, D.; Sánchez, A. Solid-State Fermentation as a Novel Paradigm for Organic Waste Valorization: A Review. Sustainability 2017, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Shivamathi, C.S.; Gunaseelan, S.; Soosai, M.R.; Vignesh, N.S.; Varalakshmi, P.; Kumar, R.S.; Karthikumar, S.; Kumar, R.V.; Baskar, R.; Rigby, S.P.; et al. Process optimization and characterization of pectin derived from underexploited pineapple peel biowaste as a value-added product. Food Hydrocoll. 2022, 123, 107141. [Google Scholar] [CrossRef]
- Lo Turco, V.; Potortì, A.G.; Tropea, A.; Dugo, G.; Di Bella, G. Element analysis of dried figs (Ficuscarica L.) from the Mediterranean areas. J. Food Compos. Anal. 2020, 90, 103503. [Google Scholar] [CrossRef]
- Potortί, A.G.; Lo Turco, V.; Saitta, M.; Bua, G.D.; Tropea, A.; Dugo, G.; Di Bella, G. Chemometric analysis of minerals and trace elements in Sicilian wines from two different grape cultivars. Nat. Prod. Res. 2017, 31, 1000–1005. [Google Scholar] [CrossRef]
- Tuttolomondo, T.; Dugo, G.; Leto, C.; Cicero, N.; Tropea, A.; Virga, G.; Leone, R.; Licata, M.; La Bella, S. Agronomical and chemical characterisation of Thymbra capitata (L.) Cav. biotypes from Sicily, Italy. Nat. Prod. Res. 2015, 29, 1289–1299. [Google Scholar] [CrossRef]
- La Torre, G.L.; Potortì, A.G.; Saitta, M.; Tropea, A.; Dugo, G. Phenolic profile in selected Sicilian wines produced by different techniques of breeding and cropping methods. Ital. J. Food Sci. 2014, 26, 41–55. [Google Scholar]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Pobiega, K. The aspects of microbial biomass use in the utilization of selected waste from the agro-food industry. Open Life Sci. 2020, 15, 787–796. [Google Scholar] [CrossRef]
- O’Shea, N.; Ktenioudaki, A.; Smyth, T.; Mcloughlin, P.; Doran, L.; Auty, M.; Arendt, E.; Gallagher, E. Physicochemical assessment of two fruit by-products as functional ingredients: Apple and orange pomace. J. Food Eng. 2015, 153, 89–95. [Google Scholar] [CrossRef]
- Dufossé, L. Microbial Production of Food Grade Pigments. Food Technol. Biotech. 2006, 44, 313–321. [Google Scholar]
- Benavente-Valdésa, J.R.; Aguilara, C.; Contreras-Esquivela, J.C.; Méndez-Zavalab, A.; Montañez, J. Strategies to enhance the production of photosynthetic pigments and lipids in Chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Ferracane, A.; Tropea, A.; Salafia, F. Production and Maturation of Soaps with Non-Edible Fermented Olive Oil and Comparison with Classic Olive Oil Soaps. Fermentation 2021, 7, 245. [Google Scholar] [CrossRef]
- Tropea, A.; Potortì, A.G.; Lo Turco, V.; Russo, E.; Vadalà, R.; Rand, R.; Di Bella, G. Aquafeed Production from Fermented Fish Waste and Lemon Peel. Fermentation 2021, 7, 272. [Google Scholar] [CrossRef]
- Jarunglumlert, T.; Bampenrat, A.; Sukkathanyawat, H.; Prommuak, C. Enhanced Energy Recovery from Food Waste by Co-Production of Bioethanol and Biomethane Process. Fermentation 2021, 7, 265. [Google Scholar] [CrossRef]
- Pandit, S.; Savla, N.; Sonawane, J.M.; Sani, A.M.; Gupta, P.K.; Mathuriya, A.S.; Rai, A.K.; Jadhav, D.A.; Jung, S.P.; Prasad, R. Agricultural Waste and Wastewater as Feedstock for Bioelectricity Generation Using Microbial Fuel Cells: Recent Advances. Fermentation 2021, 7, 169. [Google Scholar] [CrossRef]
- Beigbeder, J.-B.; de Medeiros Dantas, J.M.; Lavoie, J.-M. Optimization of Yeast, Sugar and Nutrient Concentrations for High Ethanol Production Rate Using Industrial Sugar Beet Molasses and Response Surface methodology. Fermentation 2021, 7, 86. [Google Scholar] [CrossRef]
- Asimakopoulou, G.; Karnaouri, A.; Staikos, S.; Stefanidis, S.D.; Kalogiannis, K.G.; Lappas, A.A.; Topakas, E. Production of Omega-3 Fatty Acids from the Microalga Crypthecodiniumcohnii by Utilizing Both Pentose and Hexose Sugars from Agricultural Residues. Fermentation 2021, 7, 219. [Google Scholar] [CrossRef]
- Tropea, A.; Wilson, D.; Lo Curto, R.B.; Dugo, G.; Saugman, P.; Troy-Davies, P.; Waldron, K.W. Simultaneous saccharification and fermentation of lignocellulosic waste material for second generation ethanol production. J. Biol. Res. 2015, 88, 142–143. [Google Scholar]
- Nasseri, A.T.; Rasoul-Amini, S.; Morowvat, M.H.; Ghasemi, Y. Single cell protein: Production and process. Am. J. Food Technol. 2011, 6, 103–116. [Google Scholar] [CrossRef]
- Mensah, J.K.M.; Twumasi, P. Use of pineapple waste for single cell protein (SCP) production and the effect of substrate concentration on the yield. J. Food Process Eng. 2017, 40, 1–9. [Google Scholar] [CrossRef]
- Anichebe, C.O.; Uba, B.O.; Okoye, E.L.; Onochie, C.C. Comparative Study on Single Cell Protein (SCP) Production by Trichoderma viride From Pineapple Wastes and Banana Peels. Int. J. Res. Publ. 2019, 23, 8. [Google Scholar] [CrossRef]
- Khan, M.; Khan, S.S.; Ahmed, Z.; Tanveer, A. Production of Single Cell Protein from Saccharomyces cerevisiae by utilizing Fruit Wastes. Nanobiotech. Univers. 2010, 1, 127–132. [Google Scholar]
- Ukaegbu-Obi, K.M. Single Cell Protein: A Resort to Global Protein Challenge and Waste Management. J. Microbiol. Microb. Technol. 2016, 1, 5. [Google Scholar]
- Hülsen, T.; Hsieh, K.; Lu, Y.; Tait, S.; Batstone, D.J. Simultaneous treatment and single cell protein production from agri-industrial wastewaters using purple phototrophic bacteria or microalgae–a comparison. Bioresour. Technol. 2018, 254, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Yunus, F.; Nadeem, M.; Rashid, F. Single-cell protein production through microbial conversion of lignocellulosic residue (wheat bran) for animal feed. J. Inst. Brew. 2015, 121, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Milala, M.A.; Yakubu, M.; Burah, B.; Laminu, H.H.; Bashir, H. Production and optimization of single cell protein from orange peels by Saccharomyces cerevisiae. J. Biosci. Biotechnol. Discov. 2018, 3, 99–104. [Google Scholar] [CrossRef]
- Mahan, K.M.; Le, R.K.; Wells, T.; Anderson, S.; Yuan, J.S.; Stoklosa, R.J.; Bhalla, A.; Hodge, D.B.; Ragauskas, A.J. Production of single cell protein from agro-waste using Rhodococcusopacus. J. Ind. Microbiol. Biotechnol. 2018, 45, 795–801. [Google Scholar] [CrossRef]
- Jiru, T.M.; Melku, B. Single Cell Protein Production from Torula Yeast (Cyberlindnera sp.) Using Banana Peel Hydrolysate. J. Adv. Microbiol. 2018, 13, 1–7. [Google Scholar] [CrossRef]
- Oshoma, C.E.; Eguakun-Owie, S.O.; Obuekwe, I.S. Utilization of banana peel as a substrate for Single cell protein and Amylase production by Aspergillus niger. Afr. Sci. 2017, 18, 143–149. [Google Scholar]
- Schultz, N.; Chang, L.F.; Hauck, A.; Reuss, M.; Syldatk, C. Microbial production of single-cell protein from deproteinized whey concentrates. Appl. Microbiol. Biotechnol. 2006, 69, 515–520. [Google Scholar] [CrossRef]
- Anupama, P.; Ravindra, P. Value-added food: Single cell protein. Biotech. Adv. 2000, 18, 459–479. [Google Scholar] [CrossRef]
- Aruna, T.E.; Aworh, O.C.; Raji, A.O.; Olagunju, A.I. Protein enrichment of yam peels by fermentation with Saccharomycescerevisiae (BY4743). Ann. Agric. Sci. 2017, 62, 33–37. [Google Scholar] [CrossRef]
- Soofiani, N.M.; Lundh, T.; Mahboubi, A.; Kiessling, A.; Taherzadeh, M.J. Evaluation of filamentous fungal biomass cultivated on vinasse as an alternative nutrient source of fish feed: Protein, lipid, and mineral composition. Fermentation 2019, 5, 99. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; MahboobiSoofiani, N.; Mahboubi, A.; Ferreira, J.A.; Lundh, T.; Kiessling, A.; Taherzadeh, M.J. Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed. Fermentation 2021, 7, 152. [Google Scholar] [CrossRef]
- Tropea, A.; Wilson, D.; Cicero, N.; Potortì, A.G.; La Torre, G.L.; Dugo, G.; Richardson, D.; Waldron, K.W. Development of minimal fermentation media supplementation for ethanol production using two Saccharomyces cerevisiae strains. Nat. Prod. Res. 2016, 30, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, P. Single Cell Protein Production from Lignocellulosic Biomass; Springer: Berlin/Heidelberg, Germany, 2017; pp. 31–36. [Google Scholar]
- White, L.A.; Newman, M.C.; Cromwell, G.L.; Lindemann, M.D. Brewers dried yeast as a source of mannan oligosaccharides for weanling pigs. J. Anim. Sci. 2002, 80, 2619–2628. [Google Scholar] [CrossRef]
- Lara-Flores, M.; Olvera-Novoa, M.A.; Guzma’n-Me’ndez, B.E.; Lo’-pez-Madrid, W. Use of the bacteria Streptococcus faecium and Lactobacillus acidophilus, and the yeast Saccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromis niloticus). Aquaculture 2002, 216, 193–201. [Google Scholar] [CrossRef]
- Deliana, D.; Sri, O.T.; Eka, T.; Muhammad, N.; Haznan, A. Comparison of SHF and SSF processes using enzyme and dry yeast for optimization of bioethanol production from empty fruit bunch. Energy Procedia 2015, 68, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Jiaqiang, W.; Jinlong, H.; Shumiao, Z.; Mingxiong, H.; Guoquan, H.; Xiangyang, G.; Nan, P. Single-cell Protein and Xylitol Production by a Novel Yeast Strain Candida intermedia FL023 from lignocellulosic Hydrolysates and Xylose. Appl. Biochem. Biotechnol. 2017, 185, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Jach, M.E.; Serefko, A.; Ziaja, M.; Kieliszek, M. Yeast Protein as an Easily Accessible Food Source. Metabolites 2022, 12, 63. [Google Scholar] [CrossRef]
- Putra, M.D.; Abasaeed, A.E.; Al-Zahrani, S.M. Prospective production of fructose and single cell protein from date palm waste. Electron. J. Biotechnol. 2020, 48, 46–52. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists Arlington: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Carrier, M.; Loppinet, S.A.; Denux, D.; Lasnier, J.; Ham Pichavant, F.; Cansell, F.; Aymonier, C. Thermogravimetric analysis as a new method to determine the lignocellulosic composition of biomass. Biomass Bioenergy 2011, 35, 298–307. [Google Scholar] [CrossRef]
- Waldron, K.W.; Selvendran, R.R. Composition of the cell walls of different asparagus (Asparagus officinalis) tissues. Physiol. Plant. 1990, 80, 568–575. [Google Scholar] [CrossRef]
- Mandalari, G.; Faulds, C.B.; Sancho, A.I.; Saija, A.; Bisignano, G.; Lo Curto, R.; Waldron, K.W. Fractionation and characterisation of arabinoxylans from brewers spent grain and wheat bran. J. Cereal Sci. 2005, 42, 205–212. [Google Scholar] [CrossRef]
- Blakeney, A.B.; Harris, P.J.; Henry, R.J.; Stone, B.A. A simple and rapid preparation of alditol acetates for monosaccharide analysis. Carbohydr. Res. 1983, 113, 291–299. [Google Scholar] [CrossRef]
- Bastos, R.; Coelho, E.; Coimbra, M.A. Modifications of Saccharomyces pastorianus cell wall polysaccharides with brewing process. Carbohydr. Polym. 2015, 124, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Rae, A.L.; Harris, P.J.; Bacic, A.; Clarke, A.E. Composition of the cell walls of Nicotiana alata Link et Otto pollen tubes. Planta 1985, 166, 128–133. [Google Scholar] [CrossRef]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Schiavone, M.; Sieczkowski, N.; Castex, M.; Dague, E.; François, J.M. Effects of the strain background and autolysis process on the composition and biophysical properties of the cell wall from two different industrial yeasts. FEMS Yeast Res. 2015, 15, fou012. [Google Scholar] [CrossRef] [Green Version]
- Chun, N.C.W.; Ismail, A.F.; Makhatar, M.M.Z.; Jamaluddin, F.M.N.; Tajarudin, H.A. Conversion of food waste via two-stage fermentation to controllable chicken feed nutrients by local isolated microorganism. Int. J. Recycl. Org. Waste Agric. 2020, 9, 33–47. [Google Scholar] [CrossRef]
- Aruna, T.E. Production of value-added product from pineapple peels using solid state fermentation. Innov. Food Sci. Emerg. Technol. 2019, 57, 102193. [Google Scholar] [CrossRef]
- Kosa, M.; Ragauskas, A.J. Lignin to lipid bioconversion by oleaginous Rhodococci. Green Chem. 2013, 15, 2070–2074. [Google Scholar] [CrossRef]
- Mehrnoosh, H.; Fatemeh, A.; Hamid, R.G. Single Cell Protein Production by Saccharomyces cerevisiae Using an Optimized Culture Medium Composition in a Batch Submerged Bioprocess. Appl. Biochem. 2016, 179, 1336–1345. [Google Scholar] [CrossRef]
- Zaldivar, J.; Nielsen, J.; Olsson, L. Fuel ethanol production from lignocellulose: A challenge for metabolic engineering and process integration. Appl. Microbiol. Biotechnol. 2001, 56, 17–34. [Google Scholar] [CrossRef]
- Forssell, P.; Kontkanen, H.; Schols, H.A.; Hinz, S.; Eijsink, V.G.H.; Treimo, J.; Robertson, J.A.; Waldron, K.W.; Faulds, C.B.; Buchert, J. Hydrolysis of Brewers’ Spent Grain by Carbohydrate Degrading Enzymes. J. Inst. Brew. 2008, 4, 114–120. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; Rojas-Garbanzo, C.; Velàzquez-Carrillo, C. Effetct of initial sugar concentration on the production of L(+) lactic acid by simultaneous enzymatic hydrolysis and fermentation of an agro-industrial waste product of pineapple (Ananas comosus) using Lactobacillus caseisubspeiesrhamnosus. Int. J. Biotechnol. Wellness Ind. 2012, 1, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Nadège, R.; Mourente, G.; Sadasivam, K.; Corraze, G. Replacement of a large portion of fish oil by vegetable oils does not affect lipogenesis, lipid transport and tissue lipid uptake in European seabass (Dicentrarchuslabrax L.). Aquaculture 2006, 261, 1077–1087. [Google Scholar] [CrossRef]
- Babalola, T.O.O.; Apata, D.F. Chemical and quality evaluation of some alternative lipid sources for aqua feed production. Agric. Biol. J. N. Am. 2011, 2, 935–943. [Google Scholar] [CrossRef]
- Fickers, P.; Benetti, P.H.; Wache, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilization by the yeast Yarrowialipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Yano, Y.; Oikawa, H.; Satomi, M. Reduction of lipids in fishmeal prepared from fish waste by a yeast Yarrowialipolytica. Int. J. Food Microbiol. 2007, 121, 302–307. [Google Scholar] [CrossRef]
- Oseni, O.A.; Akindahunsi, A.A. Some phytochemical properties and effect of fermentation on the seed of Jatropha curcas L. J. Food Technol. 2011, 6, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Lateef, A.; Oloke, J.K.; Gueguim Kana, E.B.; Oyeniyi, S.O.; Onifade, O.R.; Oyeleye, A.O.; Oladosu, O.C.; Oyelami, A.O. Improving the quality of agro-wastes by solid-state fermentation: Enhanced antioxidant activities and nutritional qualities. World J. Microbiol. Biotechnol. 2008, 24, 2369–2374. [Google Scholar] [CrossRef]
- Eriksen, D.T.; Hsieh, P.C.H.; Lynn, P.; Zhao, H. Directed evolution of a cellobiose utilization pathway in Saccharomyces cerevisiae by simultaneously engineering multiple proteins. Microb. Cell Fact. 2013, 12, 61. [Google Scholar] [CrossRef] [Green Version]
- van Maris, A.J.A.; Abbott, D.A.; Bellissimi, E.; van den Brink, J.; Kuyper, M.; Luttik, M.A.H.; Wisselink, H.W.; Scheffers, W.A.; van Dijken, J.P.; Pronk, J.T. Alcoholic fermentation of carbon sources in biomass hydrolysates by Saccharomyces cerevisiae: Current status. Antonie Van Leeuwenhoek 2006, 90, 391–418. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Crude Protein % | True Protein % | Crude Lipid % | Ash % | Lignin % | |
|---|---|---|---|---|---|
| 0 h | 8.52 ± 0.81 (A) | 6.06 ± 0.18 (A) | 11.47 ± 0.82 (A) | 3.93 ± 0.11 (A) | 1.65 ± 0.03 (A) |
| 24 h | 16.56 ± 0.95 (A) | 14.66 ± 0.73 (A) | 12.57 ± 0.90 (A) | 3.90 ± 0.14 (A) | 2.72 ± 0.59 (A) |
| 48 h | 24.24 ± 0.93 (A) | 22.68 ± 1.09 (A) | 12.84 ± 0.99 (A) | 2.98 ± 0.16 (AB) | 4.46 ± 0.42 (AB) |
| 72 h | 35.77 ± 1.57(B) | 33.48 ± 1.32 (B) | 12.53 ± 0.76 (A) | 2.84 ± 0.22 (AB) | 4.99 ± 0.09 (AB) |
| 96 h | 39.60 ± 1.34 (B) | 37.69 ± 1.36 (B) | 14.59 ± 0.83 (A) | 1.36 ± 0.26 (B) | 6.28 ± 0.55 (B) |
| 120 h | 40.19 ± 2.13 (B) | 38.43 ± 1.37 (B) | 14.46 ± 0.83 (A) | 1.08 ± 0.08 (B) | 6.29 ± 0.61 (B) |
| 144 h | 38.38 ± 1.07 (B) | 36.55 ± 1.34 (B) | 14.36 ± 0.76 (A) | 1.09 ± 0.07 (B) | 6.32 ± 0.62 (B) |
| SFAs (%) | MUFAs (%) | PUFAs (%) | |
|---|---|---|---|
| 0 h | 24.91 ± 1.51 (A) | 35.48 ± 2.04 (A) | 38.29 ± 1.66 (A) |
| 24 h | 23.99 ± 1.49 (A) | 36.44 ± 2.08 (A) | 36.18 ± 1.54 (A) |
| 48 h | 24.01 ± 1.45 (A) | 46.43 ± 2.21 (A) | 26.55 ± 1.47 (A) |
| 72 h | 23.34 ± 1.44 (A) | 47.99 ± 2.35 (A) | 26.56 ± 1.38 (B) |
| 96 h | 24.55 ± 1.41 (A) | 47.32 ± 2.49 (B) | 26.2 ± 1.29 (B) |
| 120 h | 24.68 ± 1.39 (A) | 47.02 ± 2.52 (B) | 26.39 ± 1.29 (B) |
| 144 h | 24.46 ± 1.29 (A) | 47.28 ± 2.85 (B) | 26.34 ± 1.14 (B) |
| Hours | Residue | Totals | Rhamnose | Fucose | Arabinose | Xylose | Mannose | Galactose | Glucose | Uronic Acid |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3.9 | 736.67 ± 36.8 | 1.34 ± 0.34 | 0.80 ± 0.12 | 75.69 ± 3.87 | 198.51 ± 20.85 | 29.67 ± 3.38 | 55.82 ± 5.61 | 268.25 ± 20.58 | 106.59 ± 9.65 |
| 24 | 2.8 | 645.01 ± 32.6 | 1.26 ± 0.26 | 0.84 ± 0.07 | 64.66 ± 3.68 | 174.95 ± 14.85 | 28.68 ± 1.86 | 44.09 ± 4.58 | 209.34 ± 16.55 | 97.62 ± 5.24 |
| 48 | 2.1 | 519.06 ± 25.8 | 1.34 ± 0.19 | 0.76 ± 0.06 | 59.78 ± 2.25 | 132.04 ± 33.62 | 15.24 ± 1.08 | 38.38 ± 1.61 | 148.55 ± 15.51 | 80.07 ± 4.25 |
| 72 | 1.9 | 387.53 ± 22.6 | 1.21 ± 0.23 | 0.63 ± 0.08 | 43.34 ± 3.17 | 97.28 ± 21.03 | 13.46 ± 1.64 | 22.65 ± 1.91 | 116.94 ± 21.62 | 57.25 ± 5.26 |
| 96 | 1.4 | 289.55 ± 26.6 | 1.01 ± 0.25 | 0.51 ± 0.04 | 35.03 ± 4.01 | 97.26 ± 6.3 | 13.24 ± 1.18 | 20.59 ± 2.48 | 85.23 ± 6.52 | 36.65 ± 3.14 |
| 120 | 1.2 | 273.73 ± 24.0 | 1.16 ± 0.20 | 0.50 ± 0.04 | 29.56 ± 1.68 | 97.06 ± 5.14 | 12.71 ± 0.95 | 17.08 ± 1.26 | 83.24 ± 6.24 | 32.21 ± 2.54 |
| 144 | 1.2 | 266.99 ± 27.3 | 1.15 ± 0.20 | 0.51 ± 0.04 | 29.71 ± 1.86 | 92.02 ± 6.01 | 12.52 ± 1.02 | 16.25 ± 1.58 | 80.78 ± 4.75 | 29.01 ± 3.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tropea, A.; Ferracane, A.; Albergamo, A.; Potortì, A.G.; Lo Turco, V.; Di Bella, G. Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation 2022, 8, 91. https://doi.org/10.3390/fermentation8030091
Tropea A, Ferracane A, Albergamo A, Potortì AG, Lo Turco V, Di Bella G. Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation. 2022; 8(3):91. https://doi.org/10.3390/fermentation8030091
Chicago/Turabian StyleTropea, Alessia, Antonio Ferracane, Ambrogina Albergamo, Angela Giorgia Potortì, Vincenzo Lo Turco, and Giuseppa Di Bella. 2022. "Single Cell Protein Production through Multi Food-Waste Substrate Fermentation" Fermentation 8, no. 3: 91. https://doi.org/10.3390/fermentation8030091
APA StyleTropea, A., Ferracane, A., Albergamo, A., Potortì, A. G., Lo Turco, V., & Di Bella, G. (2022). Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation, 8(3), 91. https://doi.org/10.3390/fermentation8030091