
Cultivation of Microalgae in Unsterile Malting Effluent for Biomass Production and Lipid Productivity Improvement

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
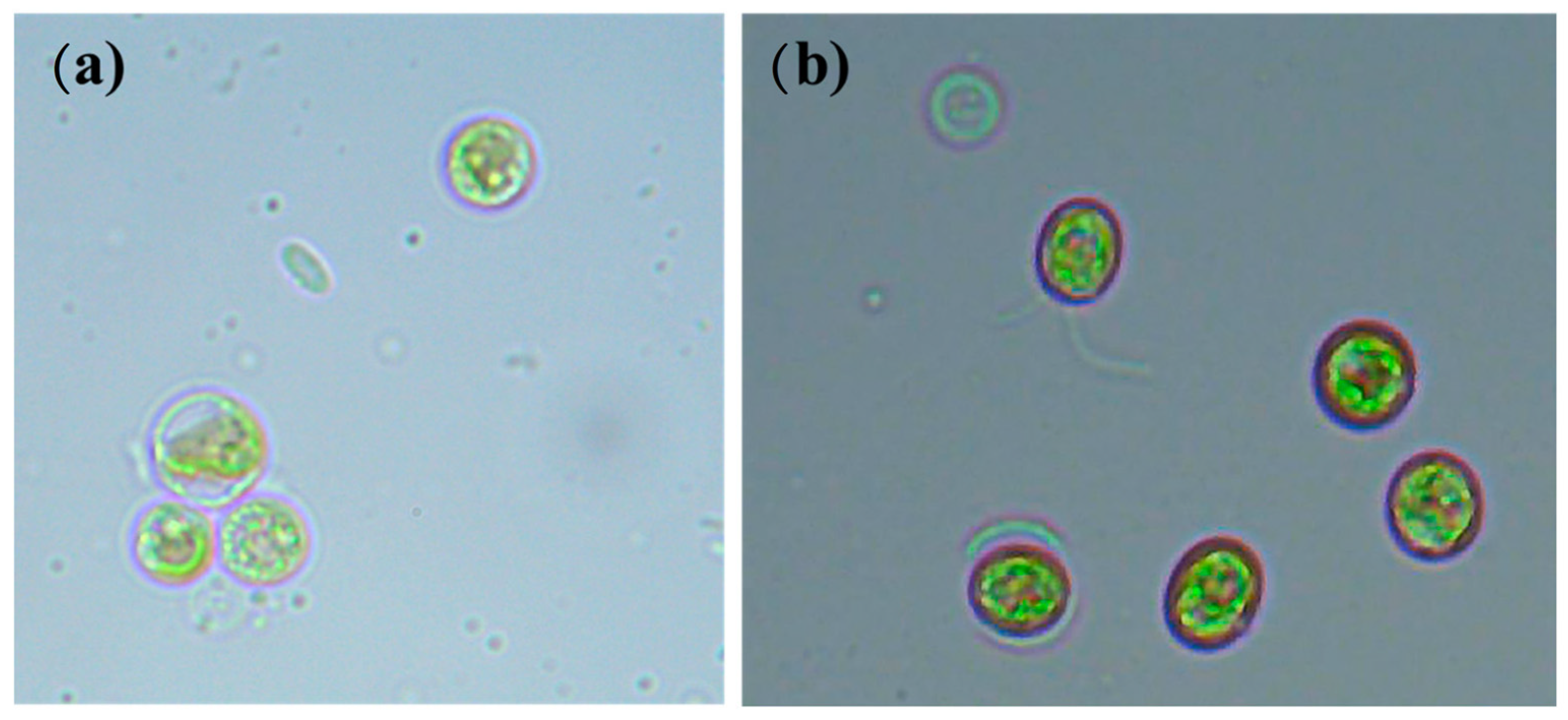
2.1. Sample Collection and Microalgae Isolation
2.2. Microalgae Strain and Cultivation Conditions
2.3. Characteristics of Malting Effluents
2.4. Measurement of Microalgae Growth Characteristics
2.5. Estimation of Total Chlorophyll Content
2.6. Lipid Extraction
2.7. Statistical Analysis
3. Results and Discussion
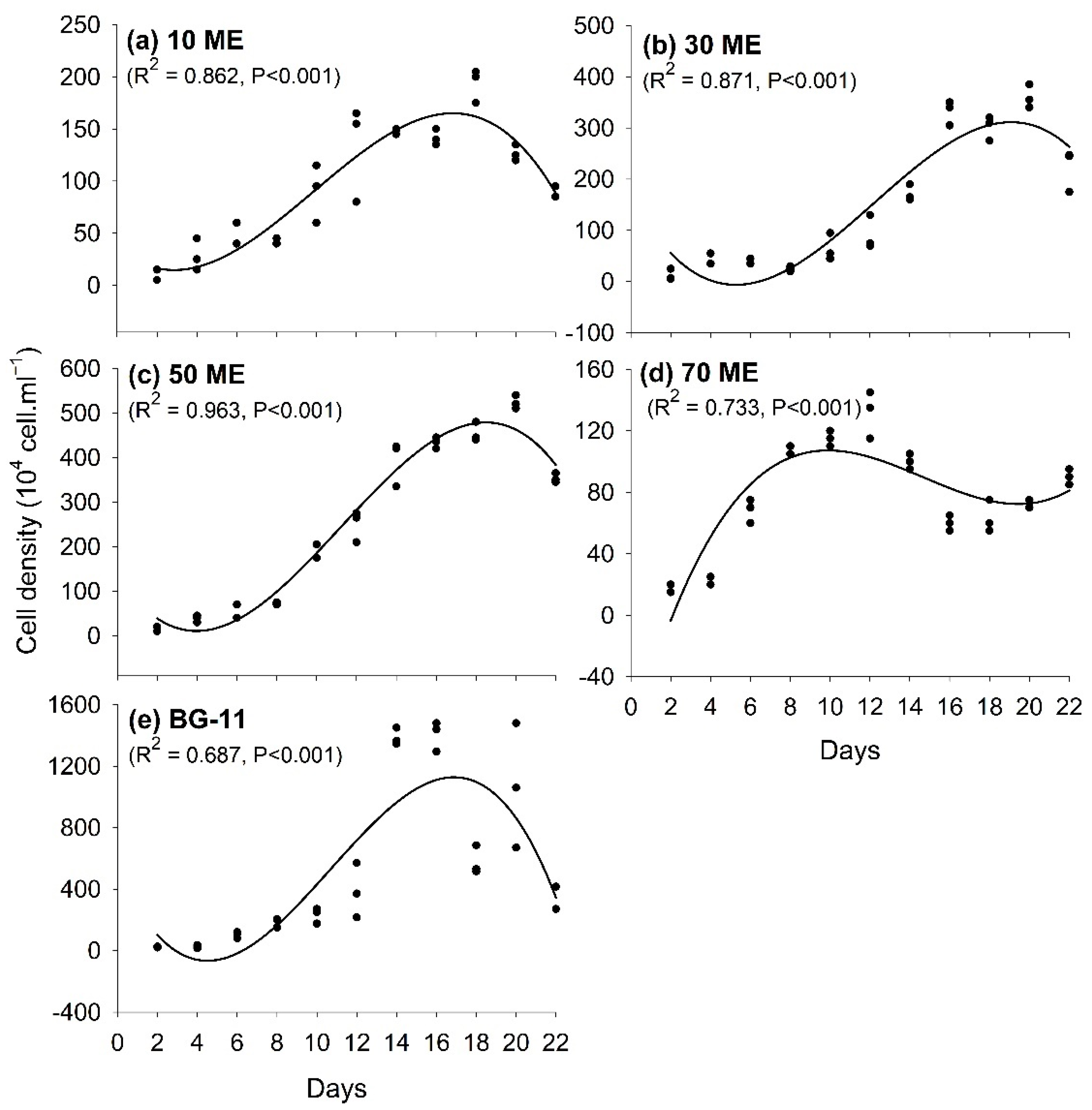
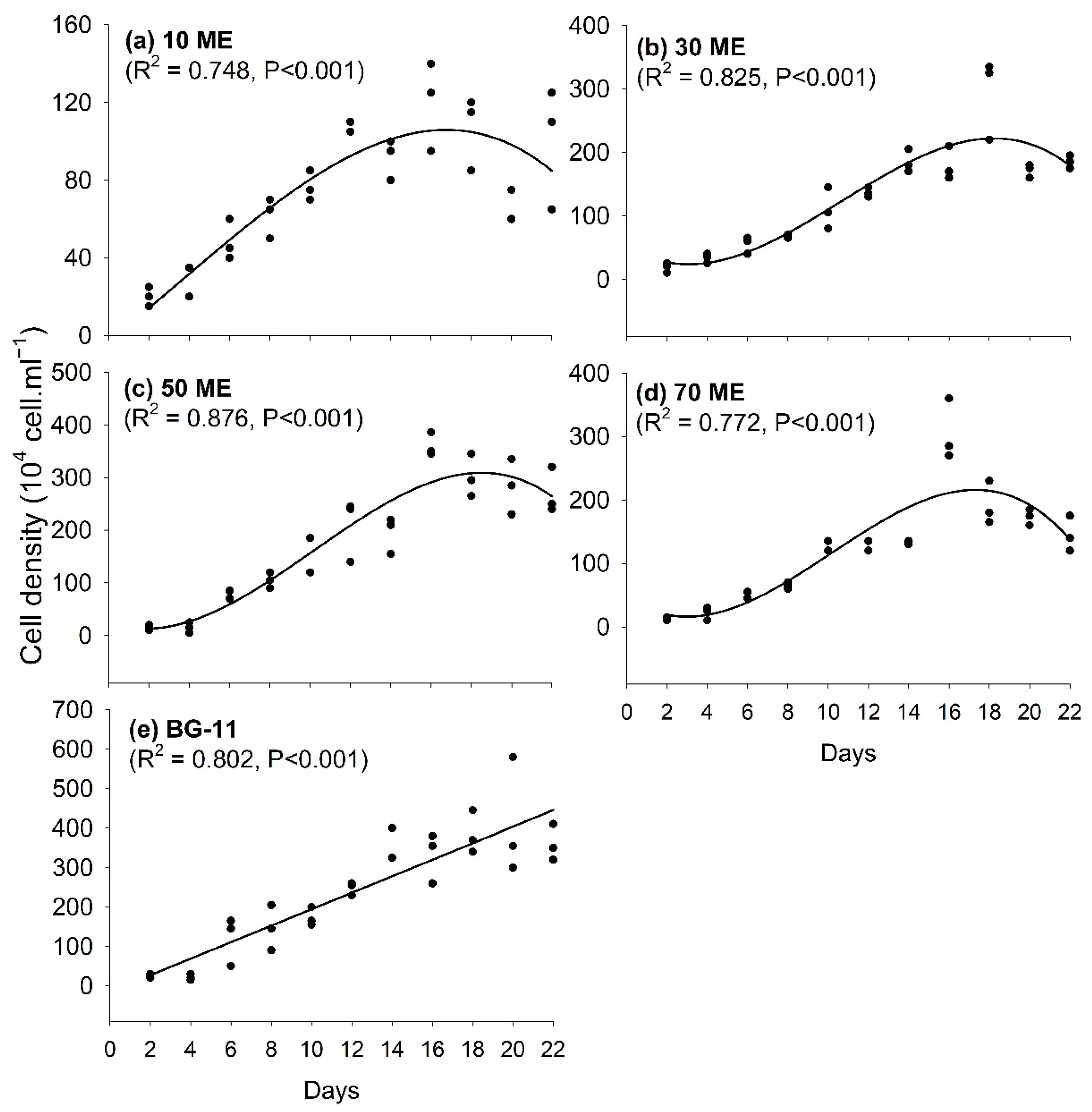
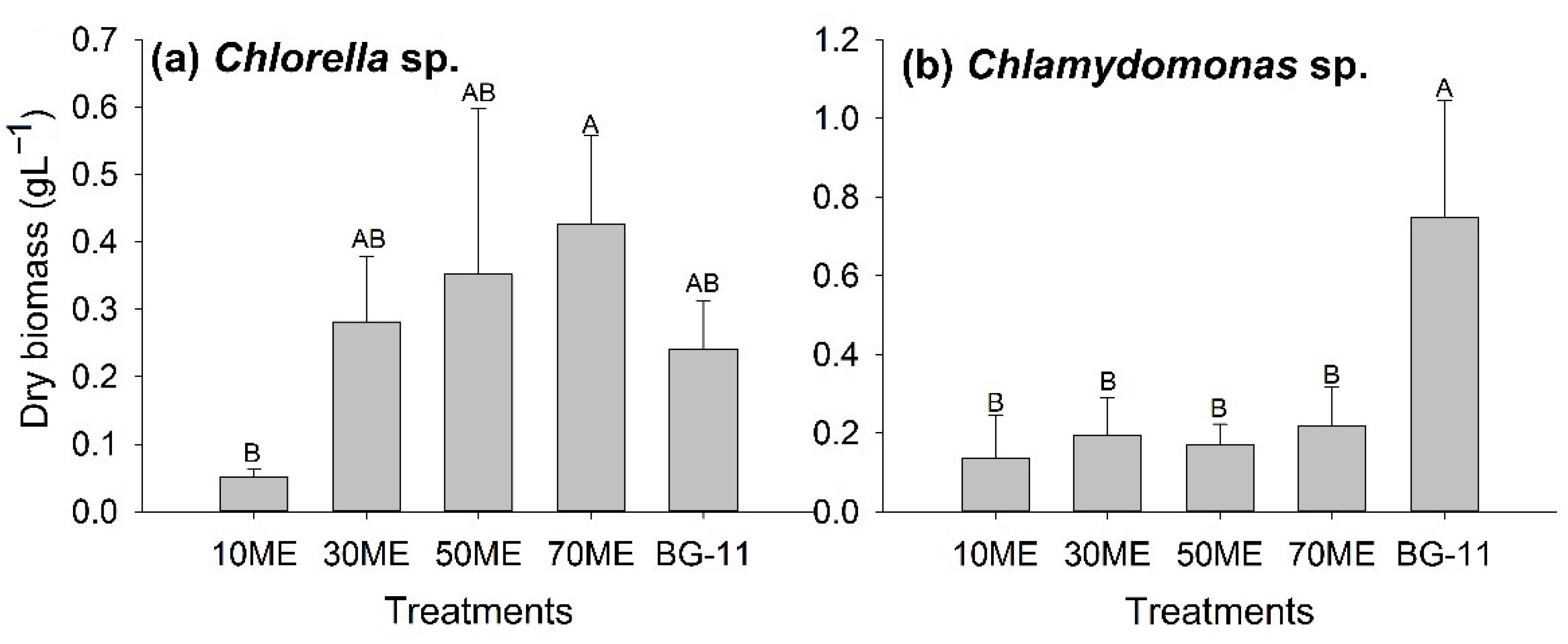
3.1. Effects of Malting Effluent on Algal Growth and Chlorophyll Content
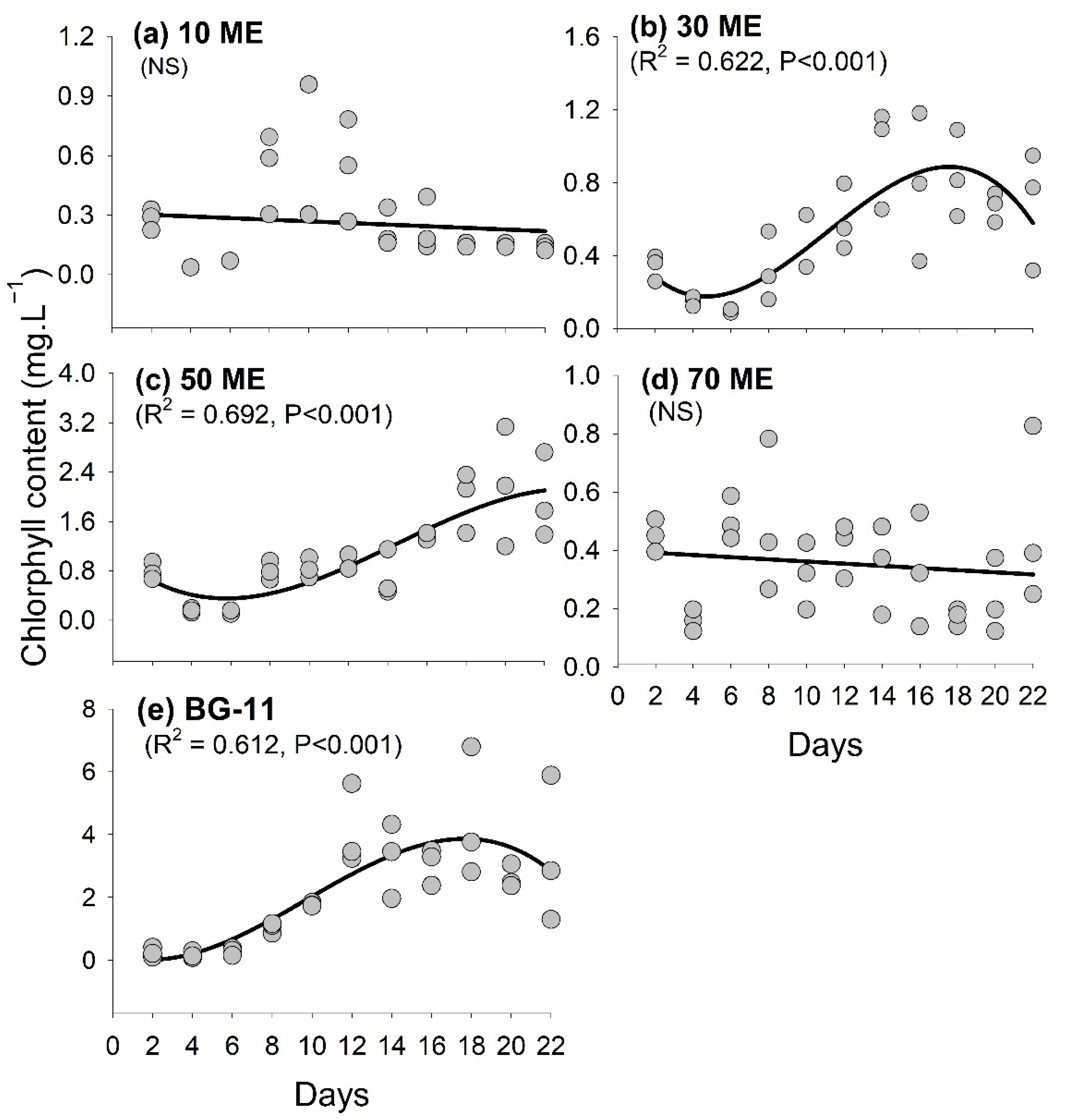
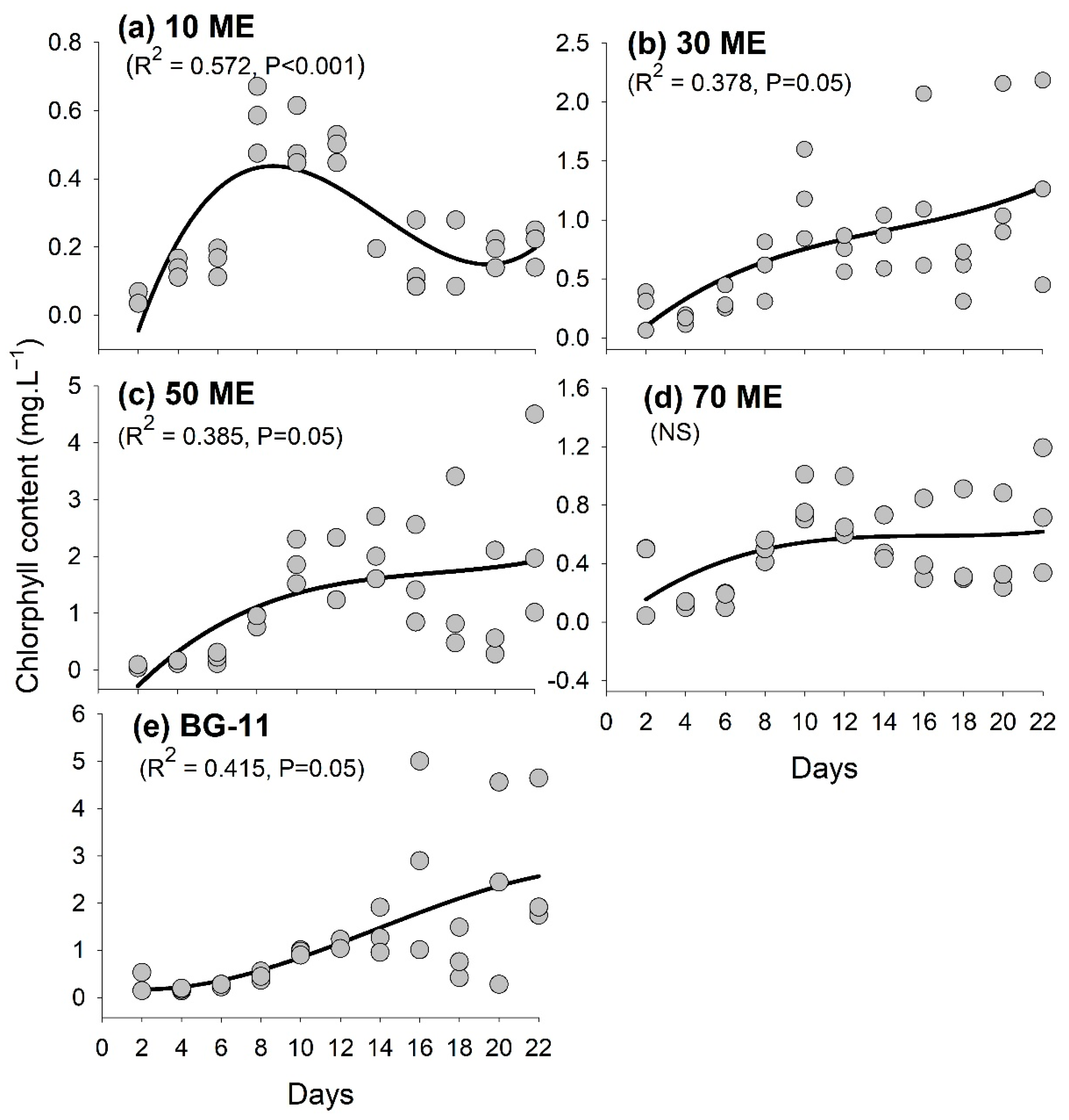
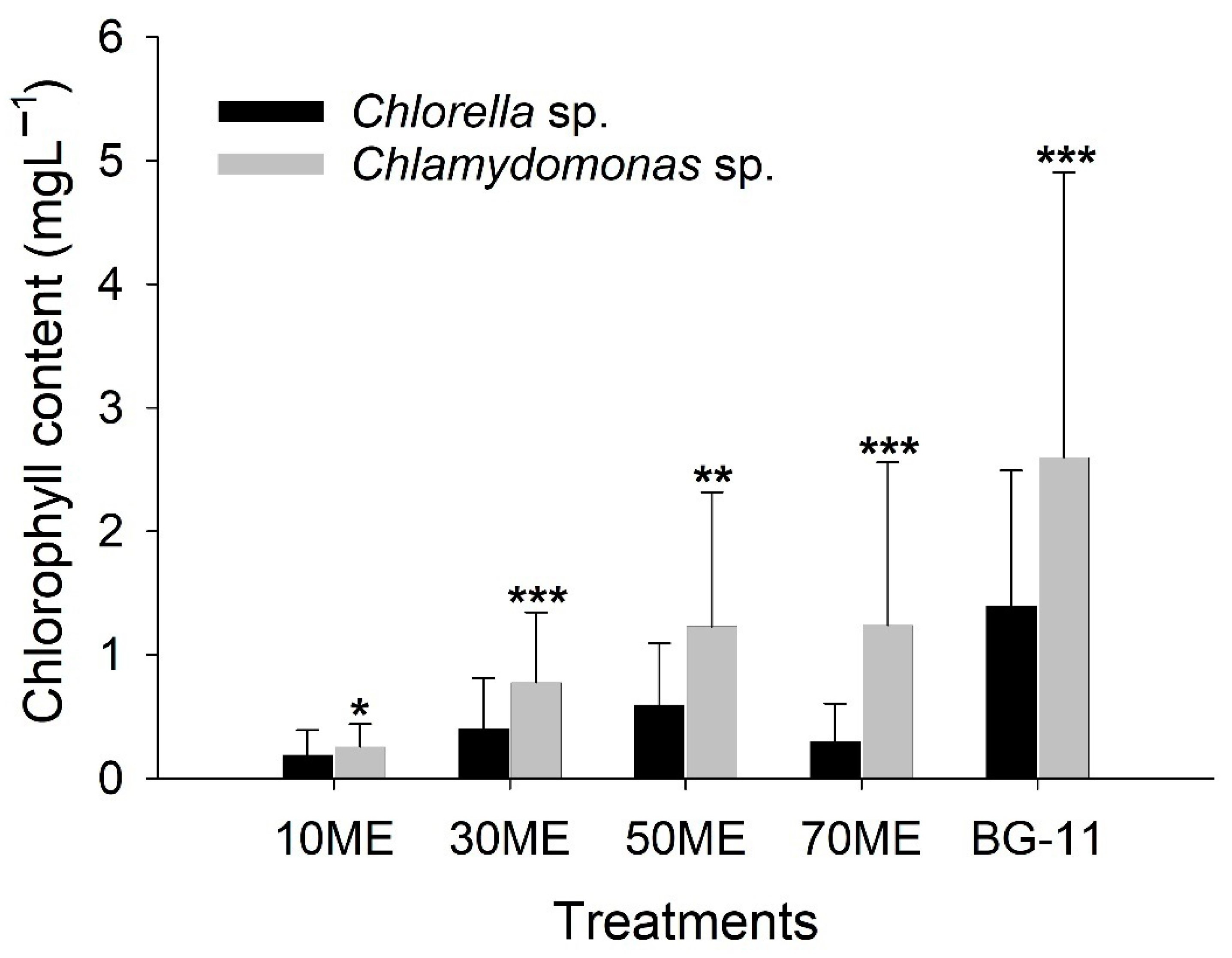
3.2. Chlorophyll Content Variation
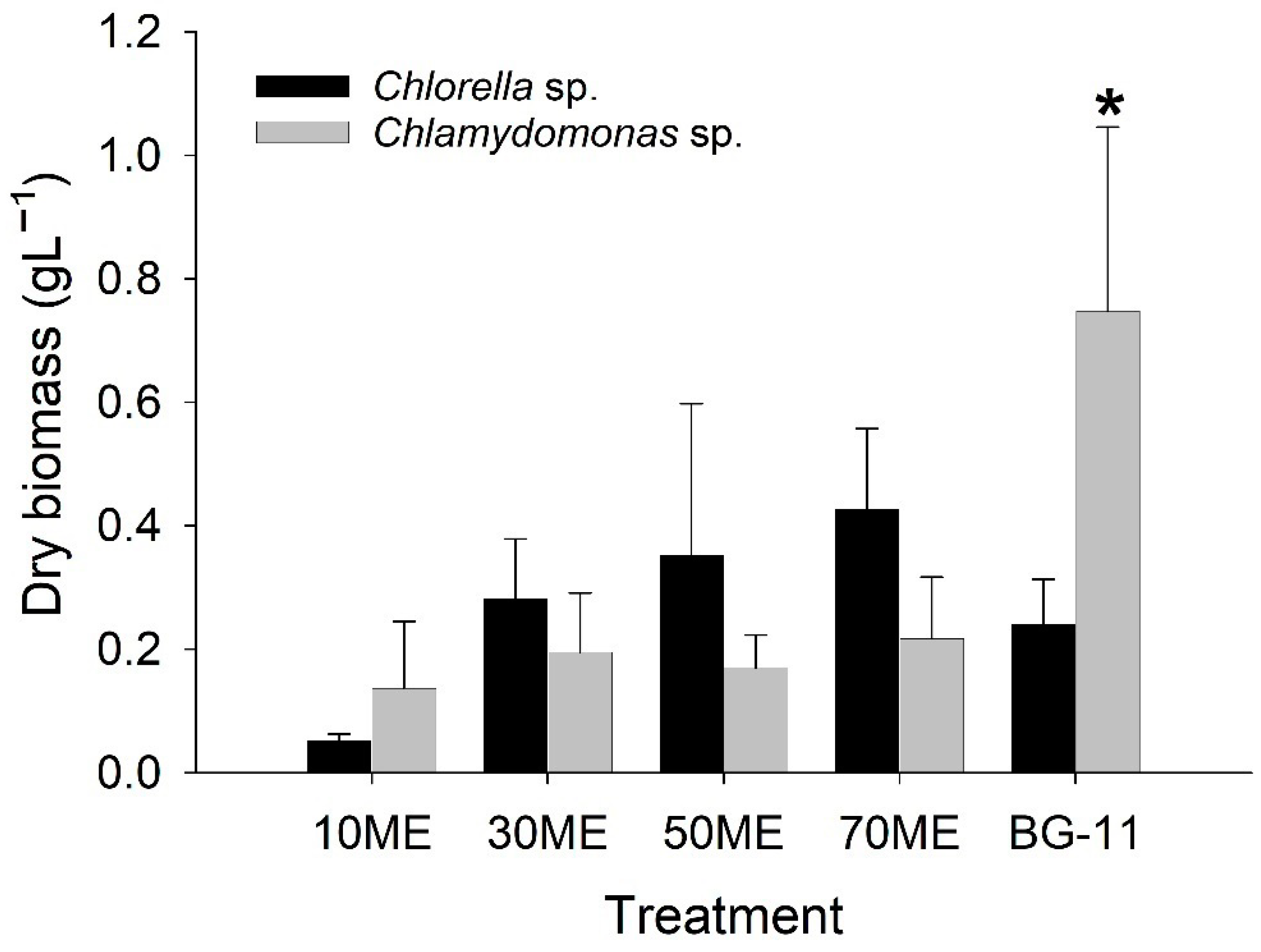
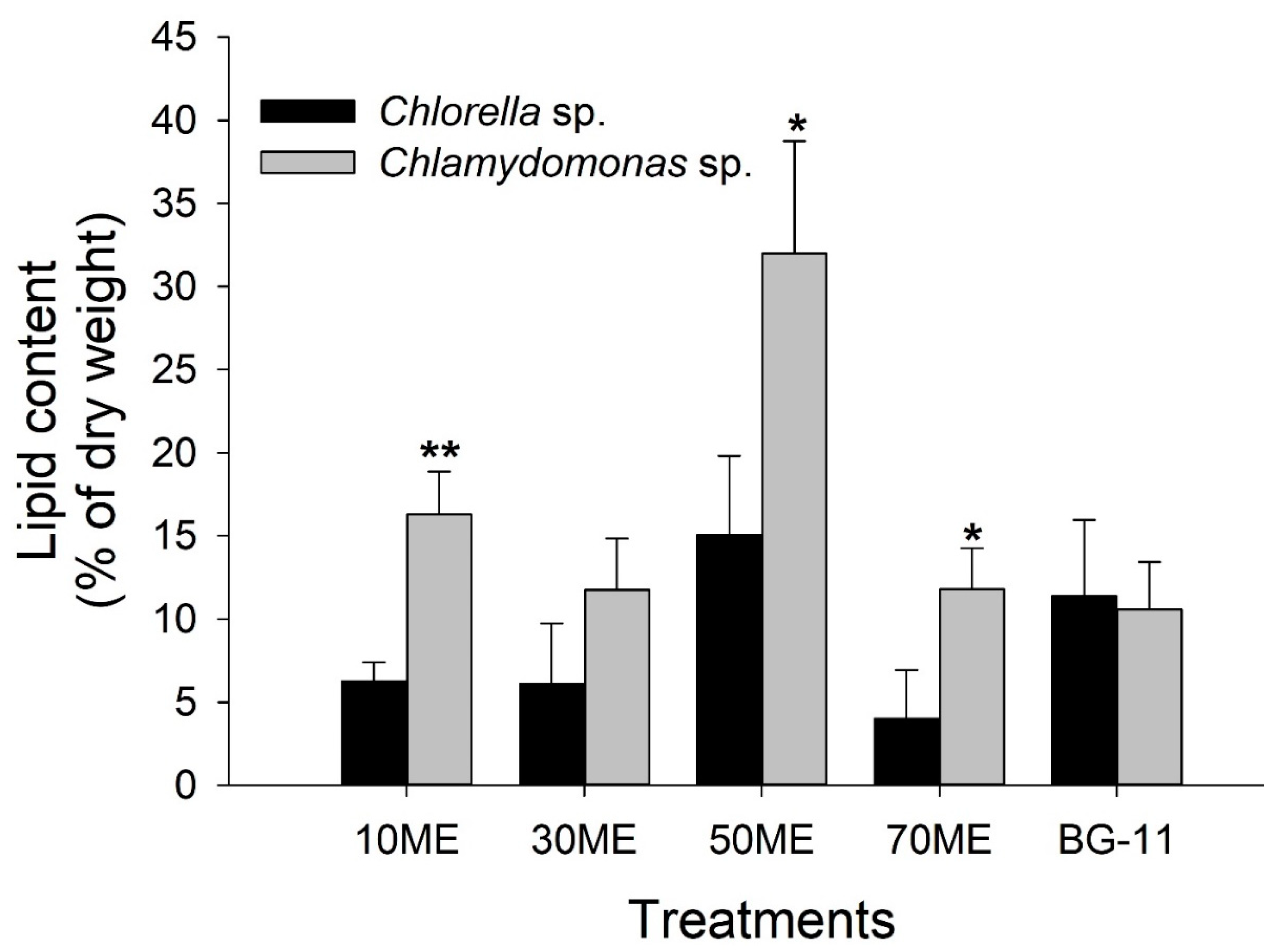
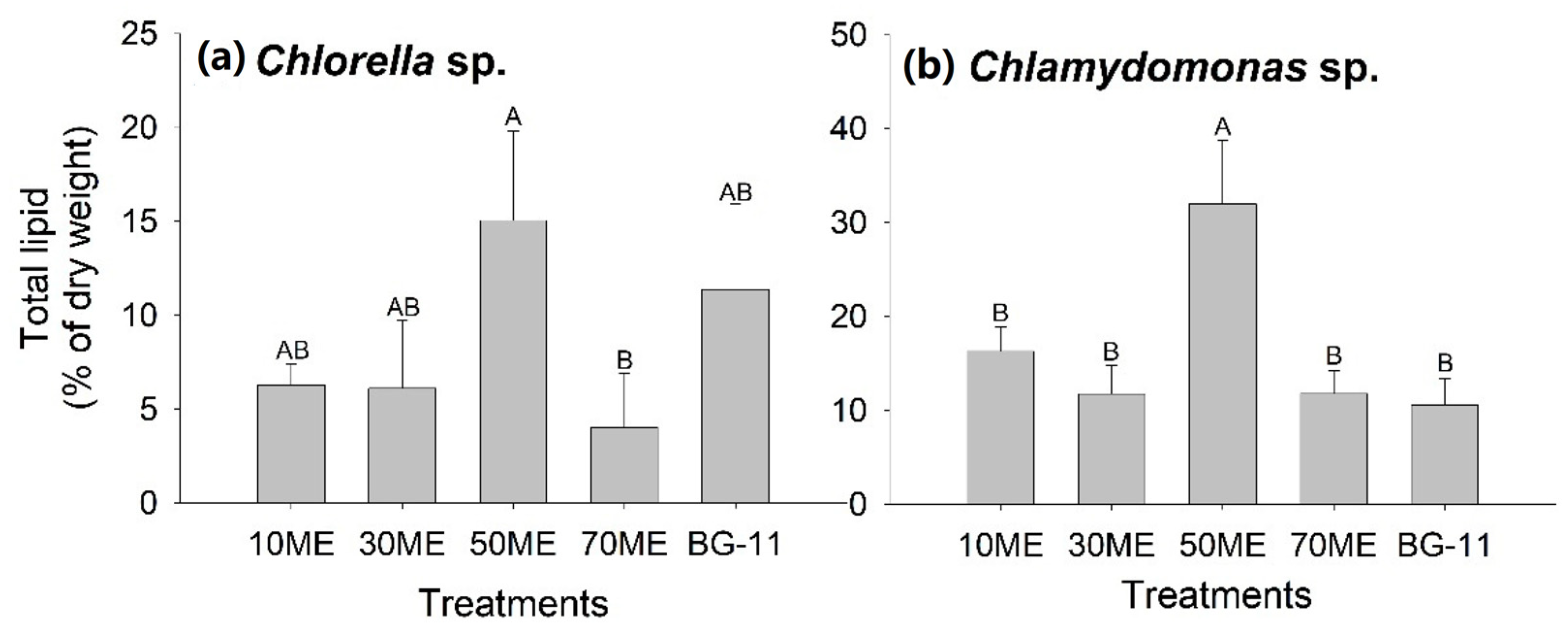
3.3. Effects of Malting Effluent on Lipid Accumulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siwal, S.S.; Zhang, Q.; Devi, N.; Saini, A.K.; Saini, V.; Pareek, B.; Gaidukovs, S.; Thakur, V.K. Recovery processes of sustainable energy using different biomass and wastes. Renew. Sust. Energ. Rev. 2021, 150, 111483. [Google Scholar] [CrossRef]
- Macqueen, D.; Korhaliller, S. Bundles of Energy: The Case for Renewable Biomass Energy; IIED: London, UK, 2011. [Google Scholar]
- Xia, A.; Jacob, A.; Tabassum, M.R.; Herrmann, C.; Murphy, J.D. Production of hydrogen, ethanol and volatile fatty acids through co-fermentation of macro-and micro-algae. Bioresour. Technol. 2016, 205, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Fan, J.; Wang, Q. Omics Application of Bio-Hydrogen Production Through Green Alga Chlamydomonas reinhardtii. Front. Bioeng. Biotechnol. 2019, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.J.l.B.; Laurens, L.M. Microalgae as biodiesel & biomass feedstocks: Review & analysis of the biochemistry, energetics & economics. Energy Environ. Sci. 2010, 3, 554–590. [Google Scholar]
- Li, Y.; Horsman, M.; Wu, N.; Lan, C.Q.; Dubois-Calero, N. Biofuels from microalgae. Biotechnol. Prog. 2008, 24, 815–820. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol. Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef]
- Khoo, K.S.; Chew, K.W.; Yew, G.Y.; Leong, W.H.; Chai, Y.H.; Show, P.L.; Chen, W.-H. Recent advances in downstream processing of microalgae lipid recovery for biofuel production. Bioresour. Technol. 2020, 304, 122996. [Google Scholar] [CrossRef]
- Kumar, B.R.; Mathimani, T.; Sudhakar, M.P.; Rajendran, K.; Nizami, A.-S.; Brindhadevi, K.; Pugazhendhi, A. A state of the art review on the cultivation of algae for energy and other valuable products: Application, challenges, and opportunities. Renew. Sust. Energ. Rev. 2021, 138, 110649. [Google Scholar] [CrossRef]
- Du, H.; Ahmed, F.; Lin, B.; Li, Z.; Huang, Y.; Sun, G.; Ding, H.; Wang, C.; Meng, C.; Gao, Z. The Effects of Plant Growth Regulators on Cell Growth, Protein, Carotenoid, PUFAs and Lipid Production of Chlorella pyrenoidosa ZF Strain. Energies 2017, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Almarashi, J.Q.M.; El-Zohary, S.E.; Ellabban, M.A.; Abomohra, A.E.-F. Enhancement of lipid production and energy recovery from the green microalga Chlorella vulgaris by inoculum pretreatment with low-dose cold atmospheric pressure plasma (CAPP). Energy Convers. Manag. 2020, 204, 112314. [Google Scholar] [CrossRef]
- Han, S.-F.; Jin, W.; Tu, R.; Abomohra, A.E.-F.; Wang, Z.-H. Optimization of aeration for biodiesel production by Scenedesmus obliquus grown in municipal wastewater. Bioprocess Biosyst. Eng. 2016, 39, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Abomohra, A.E.-F.; Eladel, H.; El-Esawi, M.; Wang, S.; Wang, Q.; He, Z.; Feng, Y.; Shang, H.; Hanelt, D. Effect of lipid-free microalgal biomass and waste glycerol on growth and lipid production of Scenedesmus obliquus: Innovative waste recycling for extraordinary lipid production. Bioresour. Technol. 2018, 249, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Qiu, S.; Amadu, A.A.; Shen, Y.; Wang, L.; Wu, Z.; Ge, S. Simultaneous improvements on nutrient and Mg recoveries of microalgal bioremediation for municipal wastewater and nickel laterite ore wastewater. Bioresour. Technol. 2020, 297, 122517. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.; Gonçalves, A.L.; Esteves, A.F.; Souza, S.M.A.G.U.d.; Souza, A.A.U.d.; Vilar, V.J.P.; Pires, J.C.M. Microalgal Growth in Paper Industry Effluent: Coupling Biomass Production with Nutrients Removal. Appl. Sci. 2020, 10, 3009. [Google Scholar] [CrossRef]
- Li, G.; Zhang, J.; Li, H.; Hu, R.; Yao, X.; Liu, Y.; Zhou, Y.; Lyu, T. Towards high-quality biodiesel production from microalgae using original and anaerobically-digested livestock wastewater. Chemosphere 2020, 273, 128578. [Google Scholar] [CrossRef]
- Ashokkumar, V.; Chen, W.-H.; Ngamcharussrivichai, C.; Agila, E.; Ani, F.N. Potential of sustainable bioenergy production from Synechocystis sp. cultivated in wastewater at large scale—A low cost biorefinery approach. Energy Convers. Manag. 2019, 186, 188–199. [Google Scholar] [CrossRef]
- Javed, F.; Aslam, M.; Rashid, N.; Shamair, Z.; Khan, A.L.; Yasin, M.; Fazal, T.; Hafeez, A.; Rehman, F.; Rehman, M.S.U.; et al. Microalgae-based biofuels, resource recovery and wastewater treatment: A pathway towards sustainable biorefinery. Fuel 2019, 255, 115826. [Google Scholar] [CrossRef]
- Hernando, M.D.; Fernández-Alba, A.R.; Tauler, R.; Barceló, D. Toxicity assays applied to wastewater treatment. Talanta 2005, 65, 358–366. [Google Scholar] [CrossRef]
- Giraldo, N.D.; Buchelly, R.J.R.; Hincapié, D.E.; Atehortua, L. Transformation of Brewery Subproducts into Valuable Biomass Using Mixotrophic Culture of Chlorella pyrenoidosa and Associated Bacteria. Braz. Arch. Biol. Technol. 2020, 63, e20190229. [Google Scholar] [CrossRef]
- Campaña, D.H.; Echevarría, M.E.U.; Airasca, A.O.; Couce, M.L.A. Physicochemical and phytotoxic characterisation of residual sludge from the malting of barley. J. Ind. Pollut. Control. 2014, 2, 115. [Google Scholar]
- Vriens, L.; Soest, H.v.; Verachtert, H. Biological treatment of malting and brewing effluents. Crit. Rev. Biotechnol. 1990, 10, 1–46. [Google Scholar] [CrossRef]
- EoC. Technical Pollution Prevention Guide for Brewery and Wine Operations in the Lower Fraser Basin; Environment Canada, Environmental Protection and Fraser Pollution Abatement: Vancouver, BC, Canada, 1997; p. 101.
- MoJ. Wastewater Systems Effluent Regulations (SOR/2012-139); Minister of Justice, Government of Canada: Ottawa, ON, Canada, 2012; p. 80.
- Stanier, R.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification, Enumeration and Use as Bioindicators; John Wiley & Sons: Oxford, UK, 2015. [Google Scholar]
- Wahidin, S.; Idris, A.; Shaleh, S.R.M. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Liu, Z.; Wang, C.; Li, S.; Kitamura, Y. Different interaction performance between microplastics and microalgae: The bio-elimination potential of Chlorella sp. L38 and Phaeodactylum tricornutum MASCC-0025. Sci. Total Environ. 2020, 723, 138146. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.-Q.; Kurade, M.B.; Jeon, B.-H. Ecotoxicological effects of enrofloxacin and its removal by monoculture of microalgal species and their consortium. Environ. Pollut. 2017, 226, 486–493. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Guo, H.; Chen, H.; Fan, L.; Linklater, A.; Zheng, B.; Jiang, D.; Qin, W. Enzymes produced by biomass-degrading bacteria can efficiently hydrolyze algal cell walls and facilitate lipid extraction. Renew. Energ. 2017, 109, 195–201. [Google Scholar] [CrossRef]
- Cobos, M.; Paredes, J.D.; Maddox, J.D.; Vargas-Arana, G.; Flores, L.; Aguilar, C.P.; Marapara, J.L.; Castro, J.C. Isolation and Characterization of Native Microalgae from the Peruvian Amazon with Potential for Biodiesel Production. Energies 2017, 10, 224. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Ding, J.; Zhao, F.; Cao, Y.; Xing, L.; Liu, W.; Mei, S.; Li, S. Cultivation of microalgae in dairy farm wastewater without sterilization. Int. J. Phytoremediation 2015, 17, 222–227. [Google Scholar] [CrossRef]
- Marchão, L.; da Silva, T.L.; Gouveia, L.; Reis, A. Microalgae-mediated brewery wastewater treatment: Effect of dilution rate on nutrient removal rates, biomass biochemical composition, and cell physiology. J. Appl. Phycol. 2018, 30, 1583–1595. [Google Scholar] [CrossRef] [Green Version]
- Vidotti, A.D.S.; Riaño-Pachón, D.M.; Mattiello, L.; Giraldi, L.A.; Winck, F.V.; Franco, T.T. Analysis of autotrophic, mixotrophic and heterotrophic phenotypes in the microalgae Chlorella vulgaris using time-resolved proteomics and transcriptomics approaches. Algal Res. 2020, 51, 102060. [Google Scholar] [CrossRef]
- Patel, A.K.; Joun, J.M.; Hong, M.E.; Sim, S.J. Effect of light conditions on mixotrophic cultivation of green microalgae. Bioresour. Technol. 2019, 282, 245–253. [Google Scholar] [CrossRef]
- Iwasaki, K.; Evenhuis, C.; Tamburic, B.; Kuzhiumparambil, U.; O’Connor, W.; Ralph, P.; Szabó, M. Improving light and CO2 availability to enhance the growth rate of the diatom, Chaetoceros muelleri. Algal Res. 2021, 55, 102234. [Google Scholar] [CrossRef]
- Marcilhac, C.; Sialve, B.; Pourcher, A.-M.; Ziebal, C.; Bernet, N.; Béline, F. Digestate color and light intensity affect nutrient removal and competition phenomena in a microalgal-bacterial ecosystem. Water Res. 2014, 64, 278–287. [Google Scholar] [CrossRef]
- O’Rourke, R.; Gaffney, M.; Murphy, R. The effects of Parachlorella kessleri cultivation on brewery wastewater. Water Sci. Technol. 2016, 73, 1401–1408. [Google Scholar] [CrossRef]
- George, B.; Pancha, I.; Desai, C.; Chokshi, K.; Paliwal, C.; Ghosh, T.; Mishra, S. Effects of different media composition, light intensity and photoperiod on morphology and physiology of freshwater microalgae Ankistrodesmus falcatus–A potential strain for bio-fuel production. Bioresour. Technol. 2014, 171, 367–374. [Google Scholar] [CrossRef]
- Mohan, S.V.; Rohit, M.; Chiranjeevi, P.; Chandra, R.; Navaneeth, B. Heterotrophic microalgae cultivation to synergize biodiesel production with waste remediation: Progress and perspectives. Bioresour. Technol. 2015, 184, 169–178. [Google Scholar] [CrossRef]
- Jena, U.; Vaidyanathan, N.; Chinnasamy, S.; Das, K. Evaluation of microalgae cultivation using recovered aqueous co-product from thermochemical liquefaction of algal biomass. Bioresour. Technol. 2011, 102, 3380–3387. [Google Scholar] [CrossRef]
- Ren, H.-Y.; Liu, B.-F.; Kong, F.; Zhao, L.; Xing, D.; Ren, N.-Q. Enhanced energy conversion efficiency from high strength synthetic organic wastewater by sequential dark fermentative hydrogen production and algal lipid accumulation. Bioresour. Technol. 2014, 157, 355–359. [Google Scholar] [CrossRef]
- Huang, Z.; Li, L.; Huang, G.; Yan, Q.; Shi, B.; Xu, X. Growth-inhibitory and metal-binding proteins in Chlorella vulgaris exposed to cadmium or zinc. Aquat. Toxicol. 2009, 91, 54–61. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Nitrogen starvation-induced cellular crosstalk of ROS-scavenging antioxidants and phytohormone enhanced the biofuel potential of green microalga Acutodesmus dimorphus. Biotechnol. Biofuels 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating environmental stresses and stress tolerance of microalgae for enhanced production of lipids and value-added products–A review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.-F.; Chu, F.-F.; Lam, P.K.S.; Zeng, R.J. Biosynthesis of high yield fatty acids from Chlorella vulgaris NIES-227 under nitrogen starvation stress during heterotrophic cultivation. Water Res. 2015, 81, 294–300. [Google Scholar] [CrossRef]
- Gojkovic, Z.; Lu, Y.; Ferro, L.; Toffolo, A.; Funk, C. Modeling biomass production during progressive nitrogen starvation by North Swedish green microalgae. Algal Res. 2020, 47, 101835. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef]
- Janssen, J.H.; Wijffels, R.H.; Barbosa, M.J. Lipid Production in Nannochloropsis gaditana during Nitrogen Starvation. Biology 2019, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, E.C.; Wilkie, A.C.; Kirst, M.; Rathinasabapathi, B. Metabolic regulation of triacylglycerol accumulation in the green algae: Identification of potential targets for engineering to improve oil yield. Plant Biotechnol. J. 2016, 14, 1649–1660. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements and Compounds | Concentrations in Undiluted Malting Effluent (mg·L−1) | Concentrations in BG-11 Medium (mg·L−1) | Elements and Compounds | Concentration in Undiluted Malting Effluent (mg·L−1) | Concentrations in BG-11 Medium (mg·L−1) |
|---|---|---|---|---|---|
| Macro Elements | Trace Elements | ||||
| Na | 310.2 | 410.3 | Fe | 7.300 | 0.688 |
| K | 1010 | 13.7 | Zn | 1.259 | 0.051 |
| Ca | 43.8 | 9.8 | Mn | 0.603 | 0.503 |
| Mg | 31.5 | 7.4 | Mo | 0.012 | 0.1546 |
| Nitrate (as N) | 820 | 247.1 | Cu | 0.080 | 0.020 |
| Ammonia nitrogen (as N) | 22.3 | 0.6 | B | 0.187 | 0.150 |
| Orthophosphate-Dissolved (as P) | 60.5 | NA | Co | 0.005 | 0.01 |
| Phosphorus (P)- Total | 65.7 | 5.4 | V | <0.01 | NA |
| Sulfate (SO42−) | 179 | 29.2 | W | <0.1 | NA |
| Heavy metals | Ti | <0.02 | NA | ||
| Cr | <0.002 | NA | Ba | 0.107 | NA |
| Ni | 0.041 | NA | Se | 0.115 | NA |
| Pb | 0.01 | NA | Si | 12.7 | NA |
| As | <0.002 | NA | Final pH | 4.62 | 7.5 |
| Ag | 0.001 | NA | |||
| Cd | <0.002 | NA | |||
| Hg | <0.002 | NA | |||
| Other elements | |||||
| Na2EDTA | NA | 1 | |||
| Citric acid Amino acids | NA 0.210 | 10.7 NA | |||
| Total Organic Carbon (TOC) | 1870 | NA | |||
| Biochemical Oxygen Demand (BOD) | 3220 | NA | |||
| Species | Cultivation Medium | Specific Growth Rate (104 Cell·mL−1) | Maximum Lipid Productivity (mg·L−1·day−1) | Percentage of Lipid Productivity (mg·L−1·day−1) |
|---|---|---|---|---|
| Chlorella sp. | 10 ME | 10.25 ± 12.69 b | 7.33 ± 2.54 b | 6.27 ± 1.13 ab |
| 30 ME | 20.3 ± 25.93 b | 35.2 ± 23.28 ab | 6.11 ± 3.63 ab | |
| 50 ME | 25.42 ± 26.91 b | 99.73 ± 54.31 a | 15.05 ± 4.76 a | |
| 70 ME | 7.25 ± 8.31 b | 32.27 ± 13.44 ab | 4.01 ± 2.92 b | |
| BG-11 | 95.17 ± 167.52 a | 55.73 ± 11.07 ab | 11.38 ± 4.57 ab | |
| Chlamydomonas sp. | 10 ME | 6.81 ± 7.5 b | 45.47 ± 29.3 c | 16.29 ± 2.59 b |
| 30 ME | 15.6 ± 18.8 ab | 49.87 ± 29.3 bc | 11.75 ± 3.08 b | |
| 50 ME | 20.28 ± 27.16 ab | 114.4 ± 15.24 ab | 31.99 ± 6.76 a | |
| 70 ME | 15.52 ± 27.12 ab | 52.8 ± 15.86 bc | 11.79 ± 2.46 b | |
| BG-11 | 26.29 ± 29.99 a | 161.33 ± 29.95 a | 10.56 ± 2.86 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatiwada, J.R.; Guo, H.; Shrestha, S.; Chio, C.; Chen, X.; Mokale Kognou, A.L.; Qin, W. Cultivation of Microalgae in Unsterile Malting Effluent for Biomass Production and Lipid Productivity Improvement. Fermentation 2022, 8, 186. https://doi.org/10.3390/fermentation8040186
Khatiwada JR, Guo H, Shrestha S, Chio C, Chen X, Mokale Kognou AL, Qin W. Cultivation of Microalgae in Unsterile Malting Effluent for Biomass Production and Lipid Productivity Improvement. Fermentation. 2022; 8(4):186. https://doi.org/10.3390/fermentation8040186
Chicago/Turabian StyleKhatiwada, Janak Raj, Haipeng Guo, Sarita Shrestha, Chonlong Chio, Xuatong Chen, Aristide Laurel Mokale Kognou, and Wensheng Qin. 2022. "Cultivation of Microalgae in Unsterile Malting Effluent for Biomass Production and Lipid Productivity Improvement" Fermentation 8, no. 4: 186. https://doi.org/10.3390/fermentation8040186
APA StyleKhatiwada, J. R., Guo, H., Shrestha, S., Chio, C., Chen, X., Mokale Kognou, A. L., & Qin, W. (2022). Cultivation of Microalgae in Unsterile Malting Effluent for Biomass Production and Lipid Productivity Improvement. Fermentation, 8(4), 186. https://doi.org/10.3390/fermentation8040186