
Evaluation of Lignocellulosic Wastewater Valorization with the Oleaginous Yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
- High lipid production rate, yield, and titer, as these are the key process indicators [23];
- Selective production of lipids without any chemical side products [24];
- A broad substrate range to metabolize all compounds present in the wastewater, i.e., organic acids, aromatics, xylan, glucan, and lignin;
- A high tolerance to the inhibiting compounds, i.e., aromatics found in the wastewater. This ability limits contamination by unwanted microorganisms and avoids the use of energy-intensive sterilization methods, e.g., autoclaving [24].
2. Materials and Methods
2.1. Steam Explosion
2.2. Strains
2.3. Substrate Measurement
2.3.1. Aromatics
2.3.2. Sugars and Organic Acids
2.3.3. Total Dissolved Organic Carbon
2.3.4. Lignin, Xylan, and Glucan
2.4. Characterization of the Phenolic Waste Stream
2.5. Repeated Batch Fermentation
2.5.1. Inoculum
2.5.2. Fermentation
2.5.3. Growth
2.5.4. Lipids
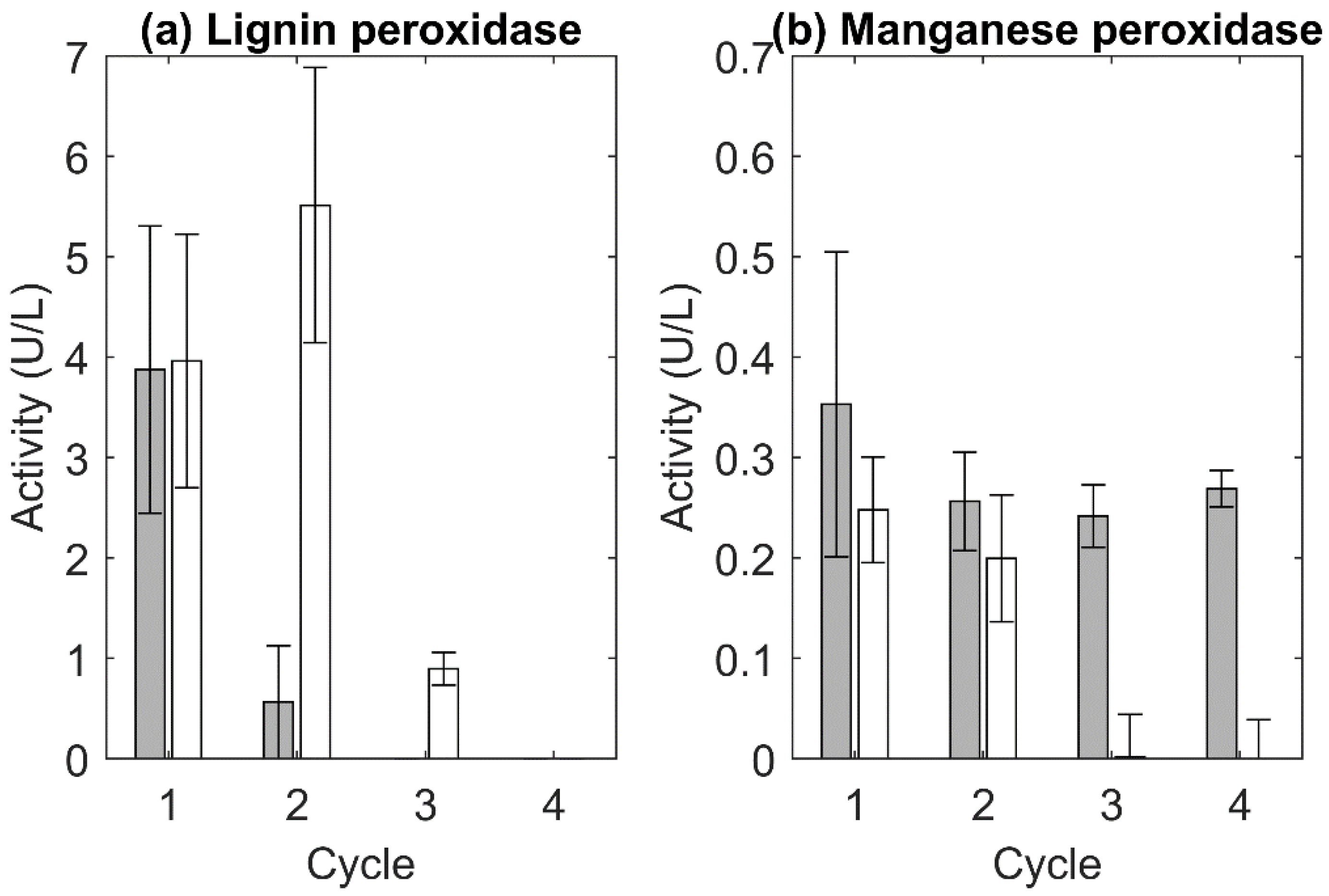
2.5.5. Laccase, Lignin Peroxidase, and Manganese Peroxidase Activity Measurement
3. Results and Discussion
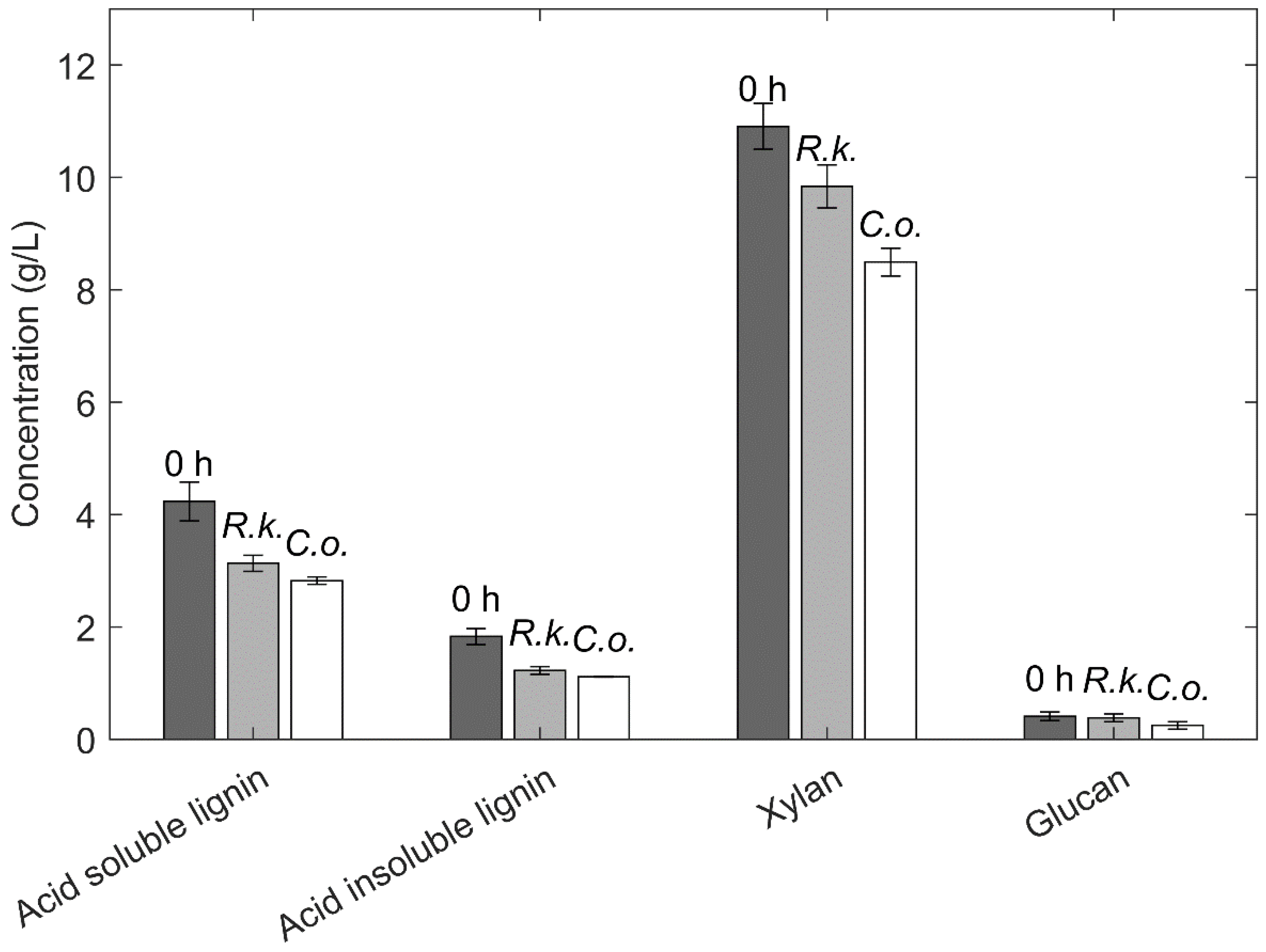
3.1. Composition of the Phenolic Waste Stream
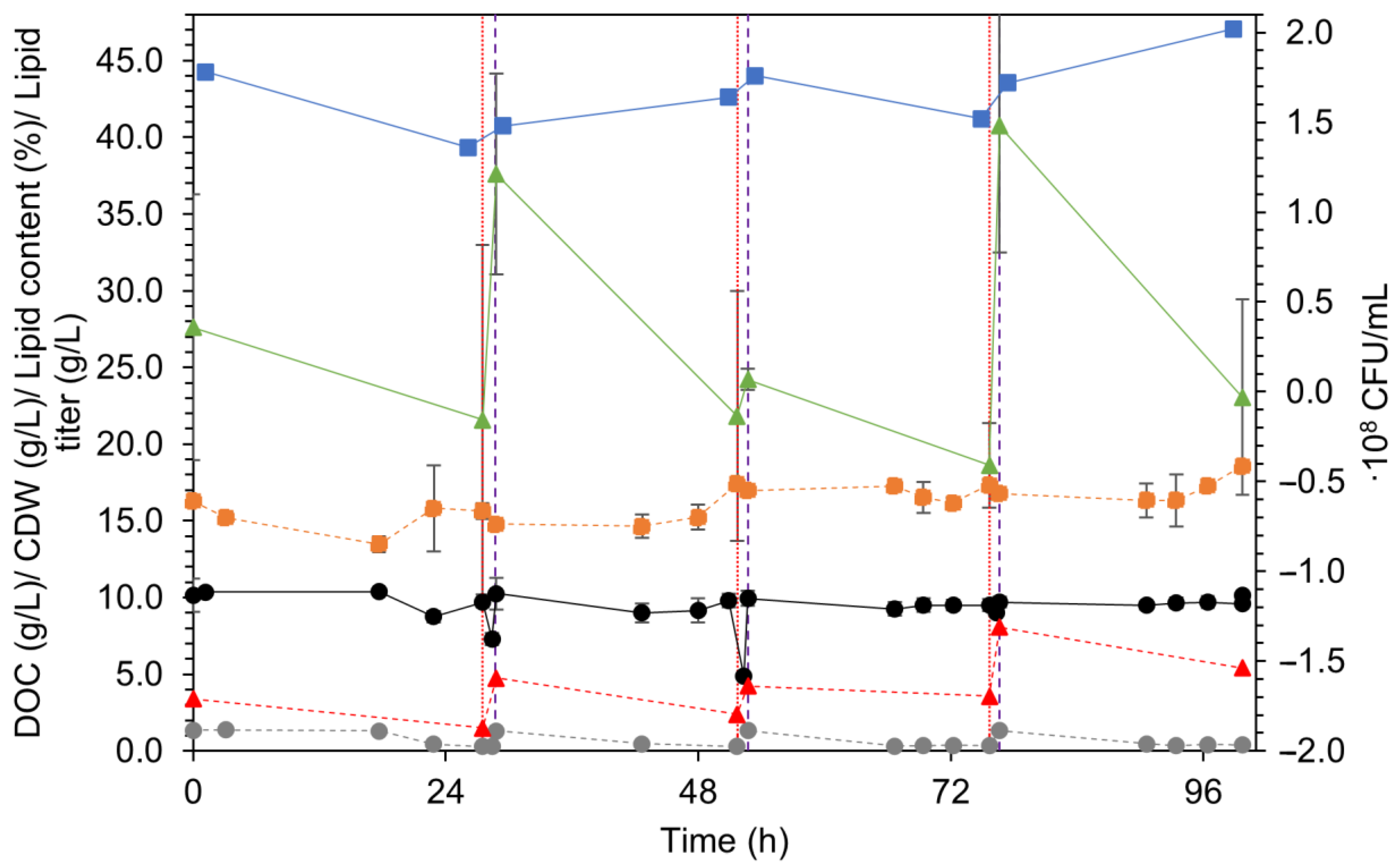
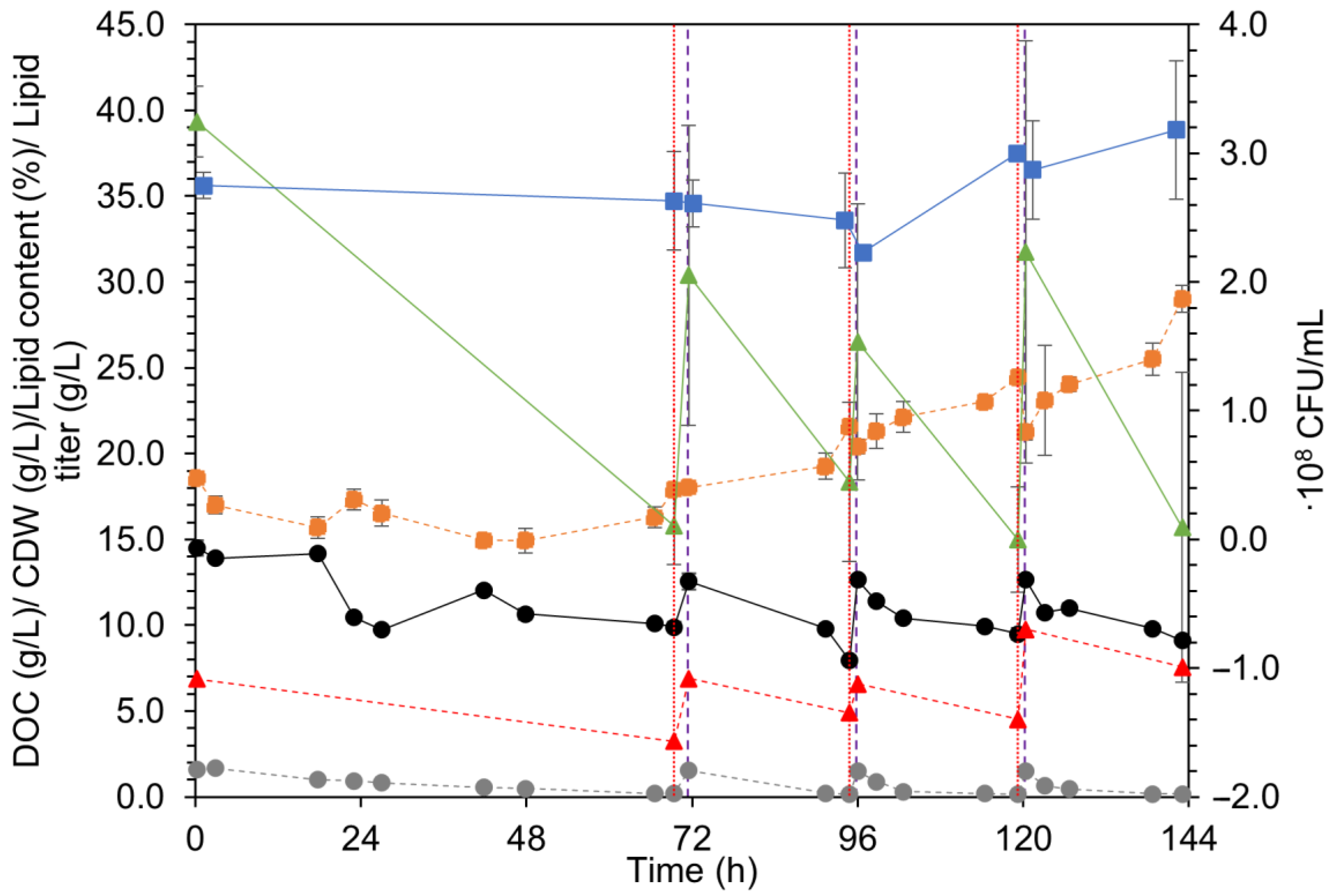
3.2. Repeated Batch: Overview
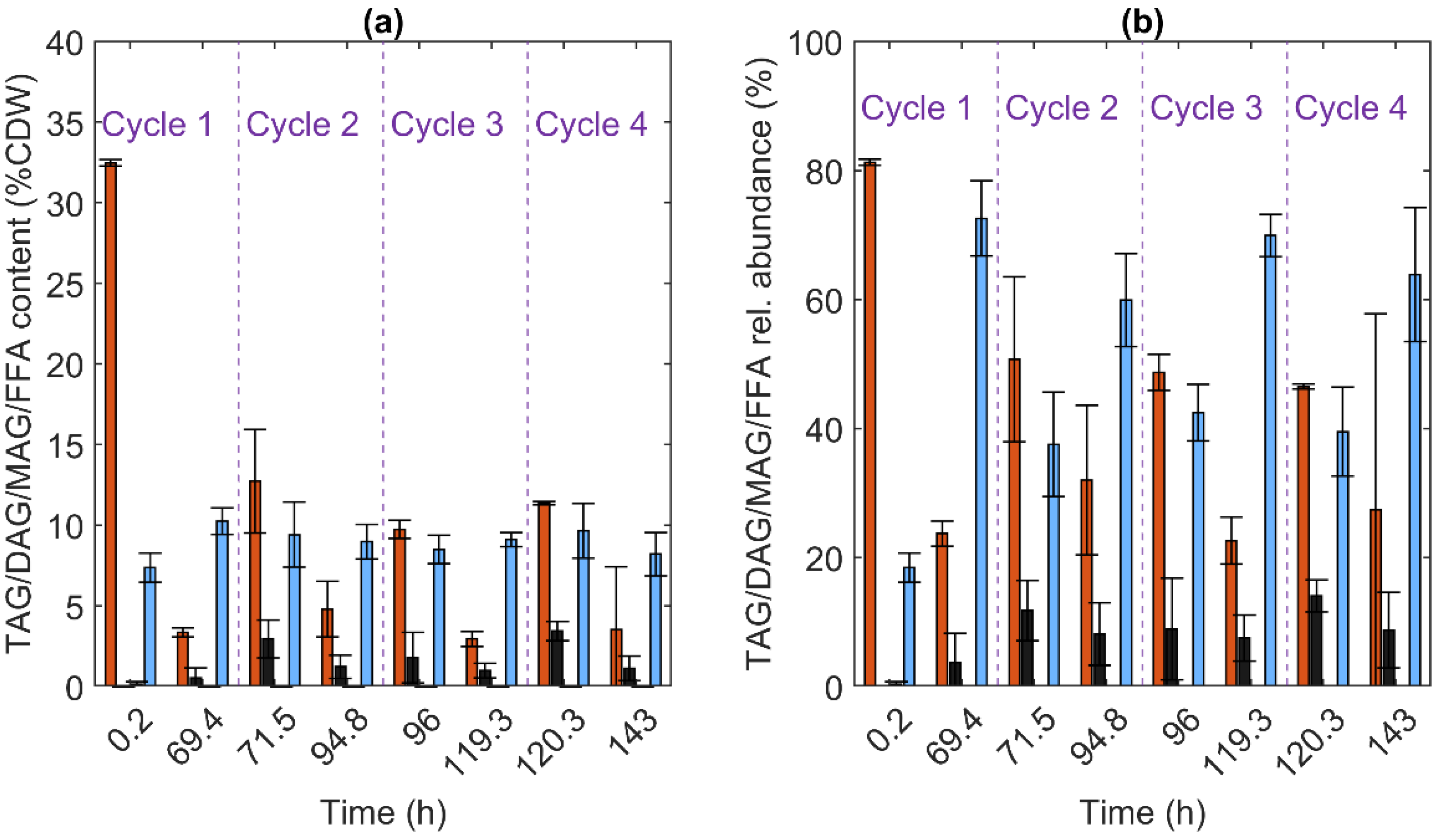
3.2.1. R. kratochvilovae
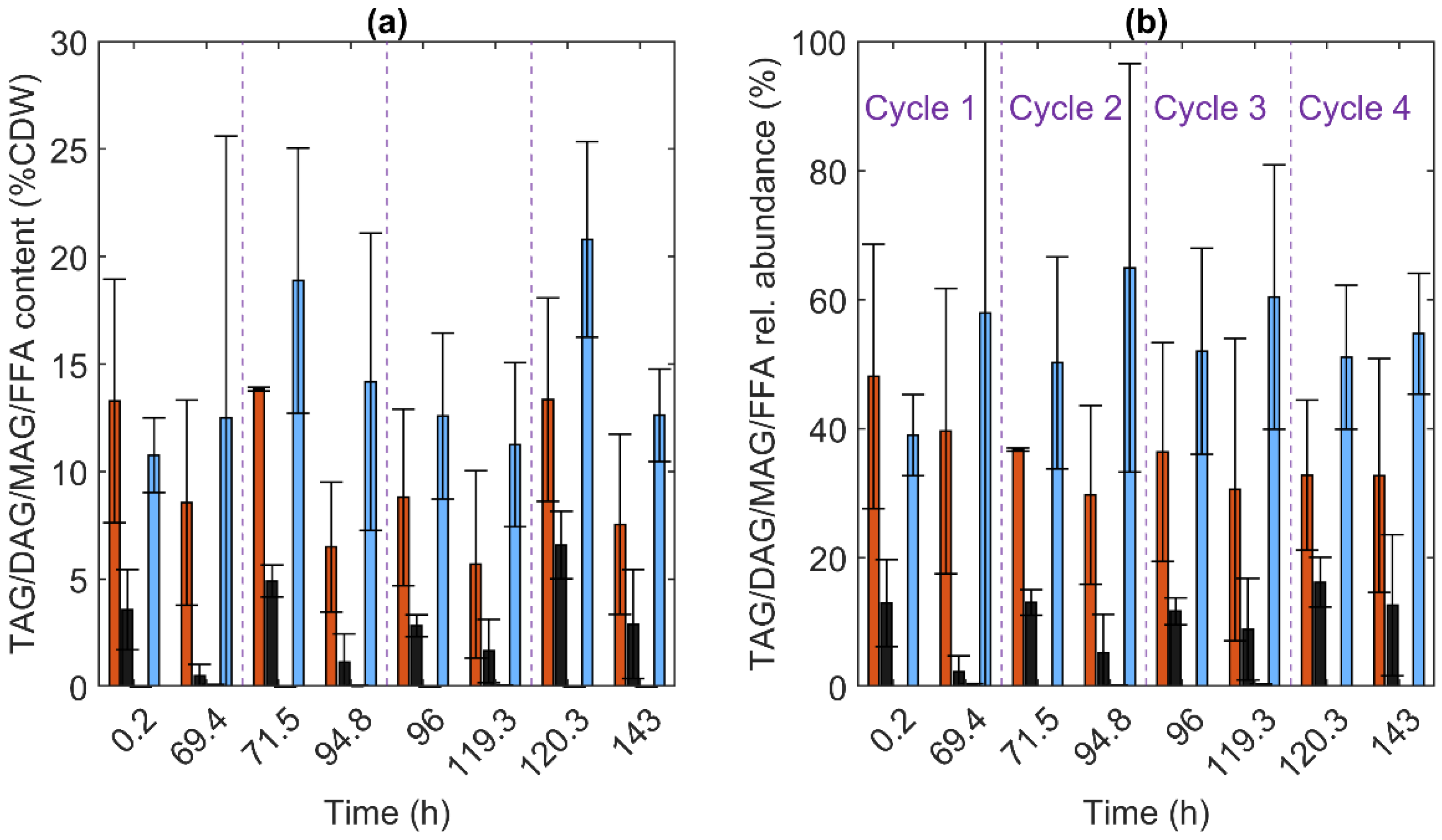
3.2.2. C. oleaginosum
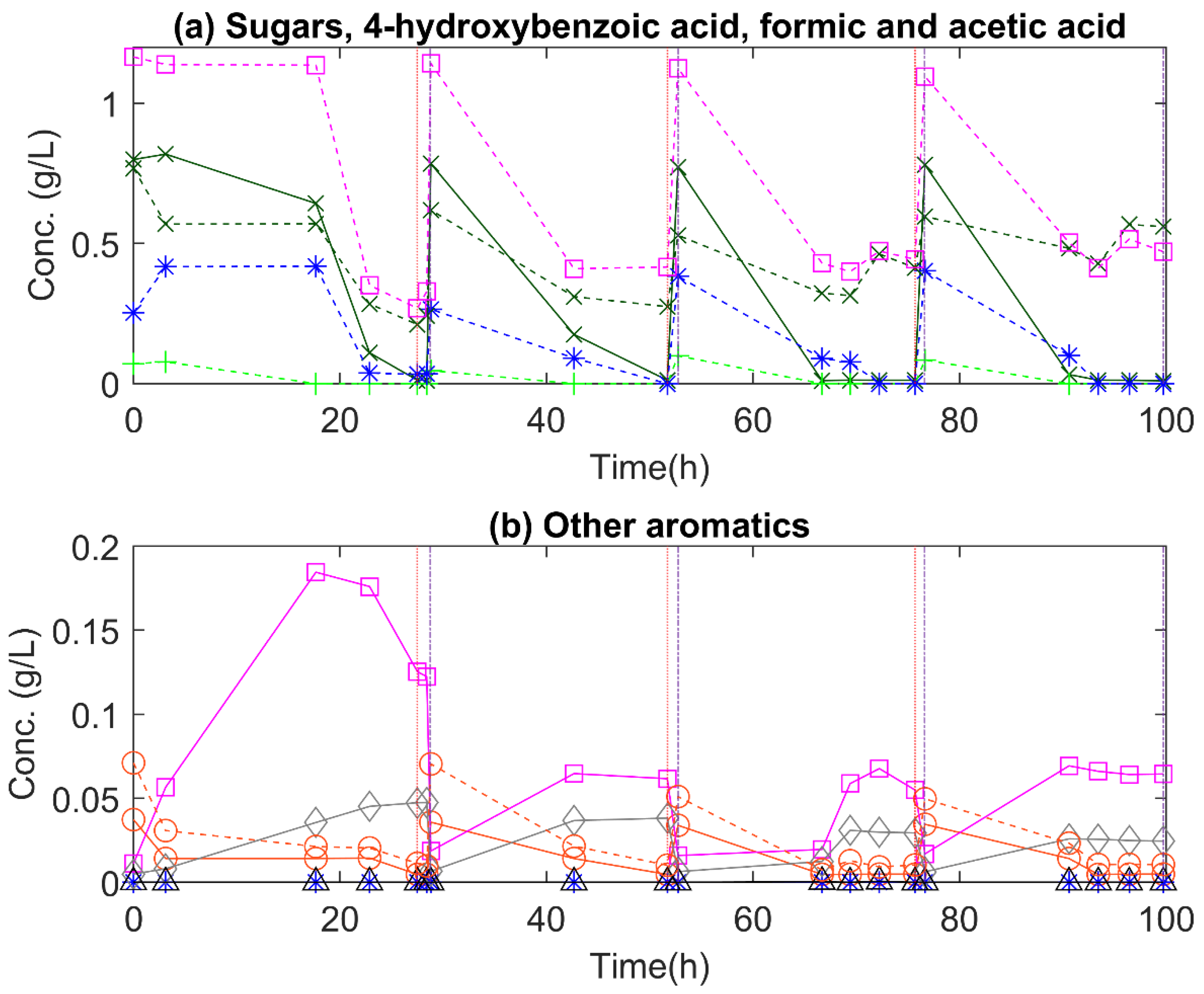
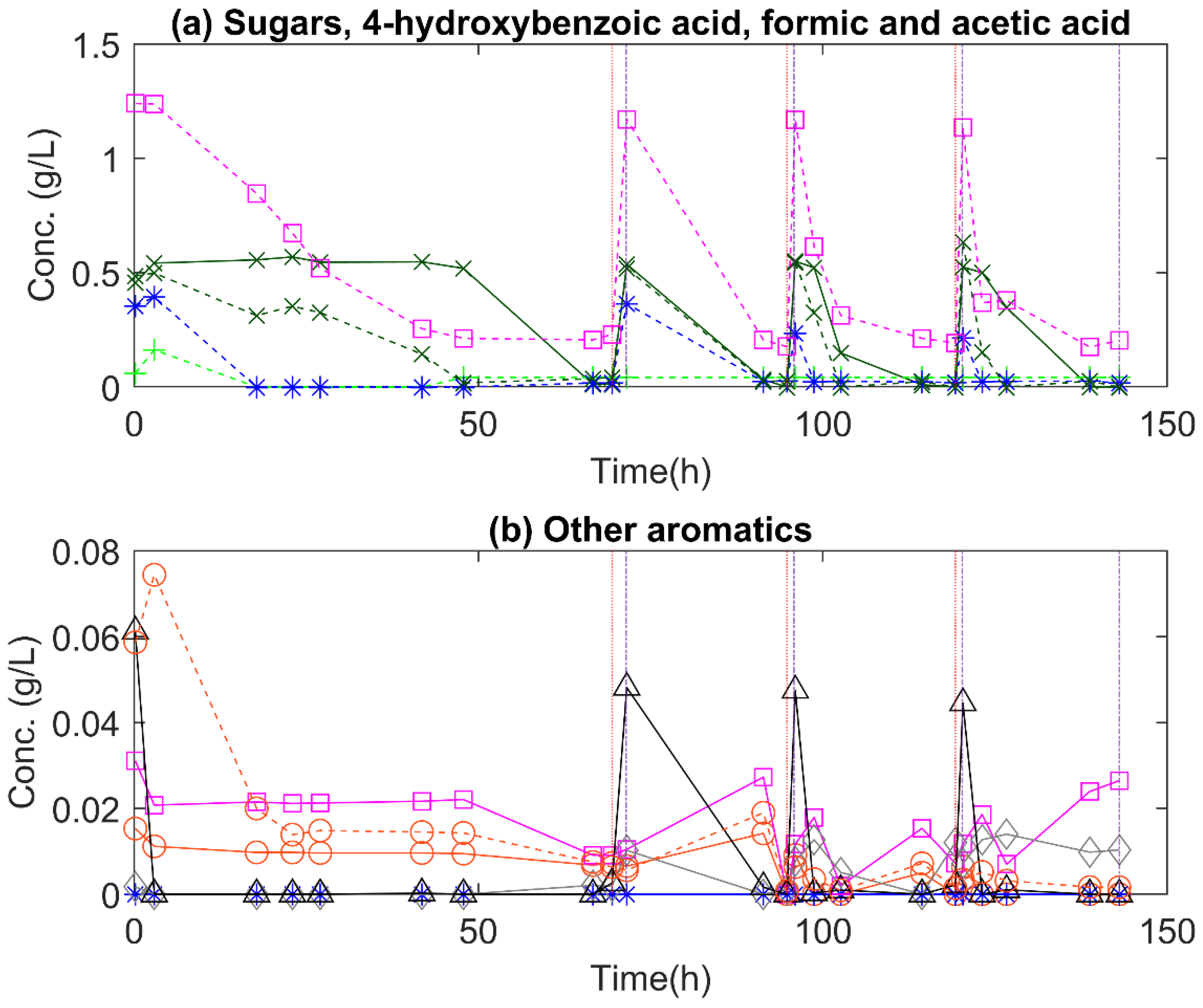
3.3. Monomeric Substrates
3.3.1. R. kratochvilovae
3.3.2. C. oleaginosum
3.4. Non-Monomeric Substrates
3.5. Lipids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista. Oil Consumption Worldwide from 1998 to 2019. Available online: https://www.statista.com/statistics/265239/global-oil-consumption-in-barrels-per-day/ (accessed on 15 April 2021).
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Sannigrahi, P.; Ragauskas, A.J.; Tuskan, G.A. Poplar as a feedstock for biofuels: A review of compositional characteristics. Biofuels Bioprod. Biorefin. 2010, 4, 209–226. [Google Scholar] [CrossRef]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Wittner, N.; Broos, W.; Bauwelinck, J.; Slezsák, J.; Vlaeminck, S.E.; Cornet, I. Enhanced fungal delignification and enzymatic digestibility of poplar wood by combined CuSO4 and MnSO4 supplementation. Process Biochem. 2021, 108, 129–137. [Google Scholar] [CrossRef]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Wells, T.; Wei, Z.; Ragauskas, A. Bioconversion of lignocellulosic pretreatment effluent via oleaginous Rhodococcus opacus DSM 1069. Biomass Bioenergy 2015, 72, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Tobin, T.; Gustafson, R.; Bura, R.; Gough, H.L. Integration of wastewater treatment into process design of lignocellulosic biorefineries for improved economic viability. Biotechnol. Biofuels 2020, 13, 24. [Google Scholar] [CrossRef]
- Scott, F.; Quintero, J.; Morales, M.; Conejeros, R.; Cardona, C.; Aroca, G. Process design and sustainability in the production of bioethanol from lignocellulosic materials. Electron. J. Biotechnol. 2013, 16, 13. [Google Scholar] [CrossRef]
- Amirsadeghi, M.; Shields-Menard, S.; French, T.; Hernandez, R. Lipid Production by Rhodotorula glutinis from Pulp and Paper Wastewater for Biodiesel Production. J. Sustain. Bioenergy Syst. 2015, 5, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Jarboui, R.; Baati, H.; Fetoui, F.; Gargouri, A.; Gharsallah, N.; Ammar, E. Yeast performance in wastewater treatment: Case study ofRhodotorula mucilaginosa. Environ. Technol. 2012, 33, 951–960. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Pruthi, V.; Pruthi, P.A. Biological treatment of pulp and paper industry effluent by oleaginous yeast integrated with production of biodiesel as sustainable transportation fuel. J. Clean. Prod. 2017, 142, 2858–2864. [Google Scholar] [CrossRef]
- Yousuf, A.; Sannino, F.; Addorisio, V.; Pirozzi, D. Microbial Conversion of Olive Oil Mill Wastewaters into Lipids Suitable for Biodiesel Production. J. Agric. Food Chem. 2010, 58, 8630–8635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, M.; Zhang, X.; Tan, T. Microbial lipid production and organic matters removal from cellulosic ethanol wastewater through coupling oleaginous yeasts and activated sludge biological method. Bioresour. Technol. 2018, 267, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Neste. Palm and Oil Seed Prices|Neste. Available online: https://www.neste.com/investors/market-data/palm-and-rapeseed-oil-prices (accessed on 27 March 2022).
- IndexMundi. Soybean Oil Monthly Price—US Dollars per Metric Ton. Available online: https://www.indexmundi.com/commodities/?commodity=soybean-oil (accessed on 27 March 2022).
- Cargill. Arachidonic Acid. Available online: https://www.cargill.com/food-bev/emea/arachidonic-acid (accessed on 24 October 2021).
- Ratledge, C. Microbial oils: An introductory overview of current status and future prospects. OCL 2013, 20, D602. [Google Scholar] [CrossRef]
- DSM. Infant Nutrition. Available online: https://www.dsm.com/human-nutrition/en/events/eln-campaign.html (accessed on 24 October 2021).
- Ochsenreither, K.; Glück, C.; Stressler, T.; Fischer, L.; Syldatk, C. Production Strategies and Applications of Microbial Single Cell Oils. Front. Microbiol. 2016, 7, 1539. [Google Scholar] [CrossRef] [Green Version]
- Dusser, P. The European Energy Policy for 2020–2030 RED II: What future for vegetable oil as a source of bioenergy? OCL 2019, 26, 51. [Google Scholar] [CrossRef] [Green Version]
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Van Winden, W.; Noorman, H. Zero Concepts in Bioprocessing. In Proceedings of the European Forum for Industrial Biotechnology, Online, 9 October 2020. [Google Scholar]
- Straathof, A.J.J.; Wahl, S.A.; Benjamin, K.R.; Takors, R.; Wierckx, N.; Noorman, H.J. Grand Research Challenges for Sustainable Industrial Biotechnology. Trends Biotechnol. 2019, 37, 1042–1050. [Google Scholar] [CrossRef]
- Yaguchi, A.; Robinson, A.; Mihealsick, E.; Blenner, M. Metabolism of aromatics by Trichosporon oleaginosus while remaining oleaginous. Microb. Cell Factories 2017, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Sartaj, K.; Arora, N.; Pruthi, V.; Pruthi, P.A. Biodegradation of phenol via meta cleavage pathway triggers de novo TAG biosynthesis pathway in oleaginous yeast. J. Hazard. Mater. 2017, 340, 47–56. [Google Scholar] [CrossRef]
- Yu, X.; Zheng, Y.; Xiong, X.; Chen, S. Co-utilization of glucose, xylose and cellobiose by the oleaginous yeast Cryptococcus curvatus. Biomass Bioenergy 2014, 71, 340–349. [Google Scholar] [CrossRef]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 2017, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Shen, H.; Zhou, W.; Wang, Y.; Yang, X.; Zhao, Z.K. Efficient conversion of acetate into lipids by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2015, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Zhang, X.; Tan, T. Simultaneously enhanced intracellular lipogenesis and β-carotene biosynthesis of Rhodotorula glutinis by light exposure with sodium acetate as the substrate. Bioresour. Technol. 2020, 295, 122274. [Google Scholar] [CrossRef]
- Ali, S.S.; Al-Tohamy, R.; Koutra, E.; El-Naggar, A.H.; Kornaros, M.; Sun, J.Z. Valorizing lignin-like dyes and textile dyeing wastewater by a newly constructed lipid-producing and lignin modifying oleaginous yeast consortium valued for biodiesel and bioremediation. J. Hazard. Mater. 2021, 403, 123575. [Google Scholar] [CrossRef]
- Yaguchi, A.; Franaszek, N.; O’Neill, K.; Lee, S.; Sitepu, I.; Boundy-Mills, K.; Blenner, M. Identification of oleaginous yeasts that metabolize aromatic compounds. J. Ind. Microbiol. Biotechnol. 2020, 47, 801–813. [Google Scholar] [CrossRef]
- Kosa, M.; Ragauskas, A.J. Lignin to lipid bioconversion by oleaginous Rhodococci. Green Chem. 2013, 15, 2070–2074. [Google Scholar] [CrossRef]
- Xenopoulos, E.; Giannikakis, I.; Chatzifragkou, A.; Koutinas, A.; Papanikolaou, S. Lipid Production by Yeasts Growing on Commercial Xylose in Submerged Cultures with Process Water Being Partially Replaced by Olive Mill Wastewaters. Processes 2020, 8, 819. [Google Scholar] [CrossRef]
- Mathew, L.; Salustra, U.; Cheryl, S.; Bonnee, R. The Oleaginous Red Yeast Rhodotorula/Rhodosporidium: A Factory for Industrial Bioproducts. In Yeasts in Biotechnology; InTech Open: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.E.S.; Sant’Ana, A.M.D.S.; Martini, M.; Sorce, C.; Andreucci, A.; de Melo, D.J.N.; da Silva, F.L.H. Rhodotorula glutinis cultivation on cassava wastewater for carotenoids and fatty acids generation. Biocatal. Agric. Biotechnol. 2019, 22, 101419. [Google Scholar] [CrossRef]
- IndexMundi. Sugar Monthly Price—US Dollars per Kilogram. Available online: https://www.indexmundi.com/commodities/?commodity=sugar (accessed on 27 March 2022).
- Yang, Q.; Zhang, H.; Li, X.; Wang, Z.; Xu, Y.; Ren, S.; Chen, X.; Xu, Y.; Hao, H.; Wang, H. Extracellular enzyme production and phylogenetic distribution of yeasts in wastewater treatment systems. Bioresour. Technol. 2012, 129, 264–273. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dayo-Owoyemi, I.; Nobre, F.S.; Pagnocca, F.C.; Chaud, L.C.S.; Pessoa, A.; Felipe, M.D.G.A.; Sette, L.D. Taxonomic assessment and enzymes production by yeasts isolated from marine and terrestrial Antarctic samples. Extremophiles 2013, 17, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Tsukagawa, Y.; Chino, M.; Soda, K. Increased Transglycosylation Activity of Rhodotorula glutinis Endo-β-Glucanase in Media Containing Organic Solvent. Biosci. Biotechnol. Biochem. 2001, 65, 1889–1892. [Google Scholar] [CrossRef] [PubMed]
- Hof, H. Rhodotorula spp. in the gut—Foe or friend? GMS Infect. Dis. 2019, 7, Doc02. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, N.; Vanderghem, C.; Blecker, C.; Paquot, M. La steam explosion: Application en tant que prétraitement de la matière cellulosique. Biotechnol. Agron. Soc. Environ. 2010, 14, 561–566. [Google Scholar]
- Transgenomic. Chromatography Products. p. 25. Available online: https://www.obrnutafaza.hr/transgenomic/Transgenomic-Overview.pdf (accessed on 23 April 2022).
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. In Laboratory Analytical Procedure (LAP); National Renewable Energy Laboratory: Golden, CO, USA, 2008. Available online: https://nrel-primo.hosted.exlibrisgroup.com/primo-explore/fulldisplay?docid=NREL_ALMA5148083920003216&context=L&vid=Pubs&lang=en_US&search_scope=PUBS&adaptor=Local%20Search%20Engine&tab=default_tab&query=any,contains,Determination%20of%20structural%20carbohydrates%20and%20lignin%20in%20biomass&offset=0 (accessed on 23 April 2022).
- Bauwelinck, J.; Caluwé, M.; Wijnants, M.; Wittner, N.; Broos, W.; Dries, J.; Akkermans, V.; Tavernier, S.; Cornet, I. Chocolate industry side streams as a valuable feedstock for microbial long-chain dicarboxylic acid production. Biochem. Eng. J. 2020, 167, 107888. [Google Scholar] [CrossRef]
- Glenn, J.K.; Gold, M.H. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1985, 242, 329–341. [Google Scholar] [CrossRef]
- Vares, T.; Kalsi, M.; Hatakka, A. Lignin Peroxidases, Manganese Peroxidases, and Other Ligninolytic Enzymes Produced by Phlebia radiata during Solid-State Fermentation of Wheat Straw. Appl. Environ. Microbiol. 1995, 61, 3515–3520. [Google Scholar] [CrossRef] [Green Version]
- Elisashvili, V.; Kachlishvili, E.; Tsiklauri, N.; Metreveli, E.; Khardziani, T.; Agathos, S.N. Lignocellulose-degrading enzyme production by white-rot Basidiomycetes isolated from the forests of Georgia. World J. Microbiol. Biotechnol. 2009, 25, 331–339. [Google Scholar] [CrossRef]
- Singh, A.K.; Bilal, M.; Iqbal, H.M.N.; Meyer, A.S.; Raj, A. Bioremediation of lignin derivatives and phenolics in wastewater with lignin modifying enzymes: Status, opportunities and challenges. Sci. Total Environ. 2021, 777, 145988. [Google Scholar] [CrossRef]
- Reed, N.R.; Kwok, E.S.C. Furfural. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 685–688. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Ravi, K.; García-Hidalgo, J.; Gorwa-Grauslund, M.F.; Liden, G. Conversion of lignin model compounds by Pseudomonas putida KT2440 and isolates from compost. Appl. Microbiol. Biotechnol. 2017, 101, 5059–5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watcharawipas, A.; Watanabe, D.; Takagi, H. Sodium Acetate Responses in Saccharomyces cerevisiae and the Ubiquitin Ligase Rsp5. Front. Microbiol. 2018, 9, 2495. [Google Scholar] [CrossRef] [PubMed]
- Mollapour, M.; Piper, P.W. Hog1 Mitogen-Activated Protein Kinase Phosphorylation Targets the Yeast Fps1 Aquaglyceroporin for Endocytosis, Thereby Rendering Cells Resistant to Acetic Acid. Mol. Cell. Biol. 2007, 27, 6446–6456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Pol, E.C.; Bakker, R.R.; Baets, P.; Eggink, G. By-products resulting from lignocellulose pretreatment and their inhibitory effect on fermentations for (bio)chemicals and fuels. Appl. Microbiol. Biotechnol. 2014, 98, 9579–9593. [Google Scholar] [CrossRef]
- Falade, A.O.; Nwodo, U.U.; Iweriebor, B.C.; Green, E.; Mabinya, L.V.; Okoh, A. Lignin peroxidase functionalities and prospective applications. MicrobiologyOpen 2016, 6, e00394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Zhang, S.; Liu, H.; Shen, H.; Lin, X.; Yang, F.; Zhou, Y.; Jin, G.; Ye, M.; Zou, H.; et al. A multi-omic map of the lipid-producing yeast Rhodosporidium toruloides. Nat. Commun. 2012, 3, 1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourist, R.; Bracharz, F.; Lorenzen, J.; Kracht, O.N.; Chovatia, M.; Daum, C.; Deshpande, S.; Lipzen, A.; Nolan, M.; Ohm, R.A.; et al. Genomics and Transcriptomics Analyses of the Oil-Accumulating Basidiomycete Yeast Trichosporon oleaginosus: Insights into Substrate Utilization and Alternative Evolutionary Trajectories of Fungal Mating Systems. mBio 2015, 6, e00918-15. [Google Scholar] [CrossRef] [Green Version]
- Magdouli, S.; Brar, S.K.; Blais, J.F. Morphology and rheological behaviour of Yarrowia lipolytica: Impact of dissolved oxygen level on cell growth and lipid composition. Process Biochem. 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Yen, H.-W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation in the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71–74. [Google Scholar] [CrossRef]
- Talley, J.; Mohiuddin, S. Fatty Acid Oxidation. Available online: https://www.ncbi.nlm.nih.gov/books/NBK556002/ (accessed on 27 March 2021).
- Kavšček, M.; Bhutada, G.; Madl, T.; Natter, K. Optimization of lipid production with a genome-scale model of Yarrowia lipolytica. BMC Syst. Biol. 2015, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; St. Leger, R.J. The Metarhizium anisopliae Perilipin Homolog MPL1 Regulates Lipid Metabolism, Appressorial Turgor Pressure, and Virulence. J. Biol. Chem. 2007, 282, 21110–21115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspeta, L.; Castillo, T.; Nielsen, J. Modifying Yeast Tolerance to Inhibitory Conditions of Ethanol Production Processes. Front. Bioeng. Biotechnol. 2015, 3, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capusoni, C.; Rodighiero, V.; Cucchetti, D.; Galafassi, S.; Bianchi, D.; Franzosi, G.; Compagno, C. Characterization of lipid accumulation and lipidome analysis in the oleaginous yeasts Rhodosporidium azoricum and Trichosporon oleaginosus. Bioresour. Technol. 2017, 238, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, C.; Xie, L.; Cao, Z. Intracellular pH and metabolic activity of long-chain dicar☐ylic acid-producing yeastCandida tropicalis. J. Biosci. Bioeng. 2003, 96, 349–353. [Google Scholar] [CrossRef]
- Liu, S.; Li, C.; Fang, X.; Cao, Z. Optimal pH control strategy for high-level production of long-chain α,ω-dicarboxylic acid by Candida tropicalis. Enzym. Microb. Technol. 2004, 34, 73–77. [Google Scholar] [CrossRef]
- Iftikhar, T.; Niaz, M.; Zia, M.A.; Haq, I.U. Production of extracellular lipases by Rhizopus oligosporus in a stirred fermentor. Braz. J. Microbiol. 2010, 41, 1124–1132. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration 1 (mg/L) | OC 2 Concentration 1 (mg C/L) |
|---|---|---|
| MONOMERIC | ||
| Furans | ||
| 5-HMF | 19.8 ± 0.1 | 11.131 ± 0.06 |
| Furfural | 29.7 ± 0.1 | 18.6 ± 0.06 |
| Phenolics | ||
| 3,4-Dihydroxybenzaldehyde | 7 ± 0 | 4.13 ± 0 |
| 4-Hydroxybenzoic acid | 365 ± 2 | 222 ± 1 |
| 4-Hydroxybenzaldehyde | 6.1 ± 0.2 | 4.2 ± 0.1 |
| Vanillic acid | 10.8 ± 0.1 | 62.0 ± 0.05 |
| Syringic acid | 2.78 ± 0.03 | 1.52 ± 0.02 |
| Vanillin | 6.6 ± 0.8 | 4.2 ± 0.5 |
| Syringaldehyde | 36.9 ± 0. 4 | 21.9 ± 0.2 |
| Sugars | ||
| Glucose | 77.97 ± 0.05 | 31.19 ± 0.02 |
| Xylose | 419 ± 2 | 168 ± 1 |
| Organic acids | ||
| Formic acid | 546 ± 40 | 142 ± 10 |
| Acetic acid | 1134 ± 6 | 454 ± 2 |
| NON-MONOMERIC | ||
| Lignin 3 | 6064 ± 372 | 1058 ± 762 |
| Acid soluble lignin 4 | 4234 ± 343 | NA |
| Acid Insoluble lignin | 1830 ± 144 | NA |
| Xylan | 10,853 ± 862 | 4934 ± 392 |
| Glucan | 390 ± 4 | 175 ± 17 |
| TOTAL OC (MEASURED) | 7255 ± 653 | |
| Monomeric OC | 1089 ± 11 | |
| Non-monomeric OC 5 | 6166 ± 653 | |
| TOTAL NITROGEN IN mg N/L | 25 ± 3 | |
| C/N RATIO IN g/g | 290 ± 44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broos, W.; Wittner, N.; Geerts, J.; Dries, J.; Vlaeminck, S.E.; Gunde-Cimerman, N.; Richel, A.; Cornet, I. Evaluation of Lignocellulosic Wastewater Valorization with the Oleaginous Yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509. Fermentation 2022, 8, 204. https://doi.org/10.3390/fermentation8050204
Broos W, Wittner N, Geerts J, Dries J, Vlaeminck SE, Gunde-Cimerman N, Richel A, Cornet I. Evaluation of Lignocellulosic Wastewater Valorization with the Oleaginous Yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509. Fermentation. 2022; 8(5):204. https://doi.org/10.3390/fermentation8050204
Chicago/Turabian StyleBroos, Waut, Nikolett Wittner, Jordi Geerts, Jan Dries, Siegfried E. Vlaeminck, Nina Gunde-Cimerman, Aurore Richel, and Iris Cornet. 2022. "Evaluation of Lignocellulosic Wastewater Valorization with the Oleaginous Yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509" Fermentation 8, no. 5: 204. https://doi.org/10.3390/fermentation8050204
APA StyleBroos, W., Wittner, N., Geerts, J., Dries, J., Vlaeminck, S. E., Gunde-Cimerman, N., Richel, A., & Cornet, I. (2022). Evaluation of Lignocellulosic Wastewater Valorization with the Oleaginous Yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509. Fermentation, 8(5), 204. https://doi.org/10.3390/fermentation8050204