
Construction of L-Asparaginase Stable Mutation for the Application in Food Acrylamide Mitigation

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Chemicals
2.2. Construction of Recombinant Strains
2.3. Expression, Purification, and Activity Assay
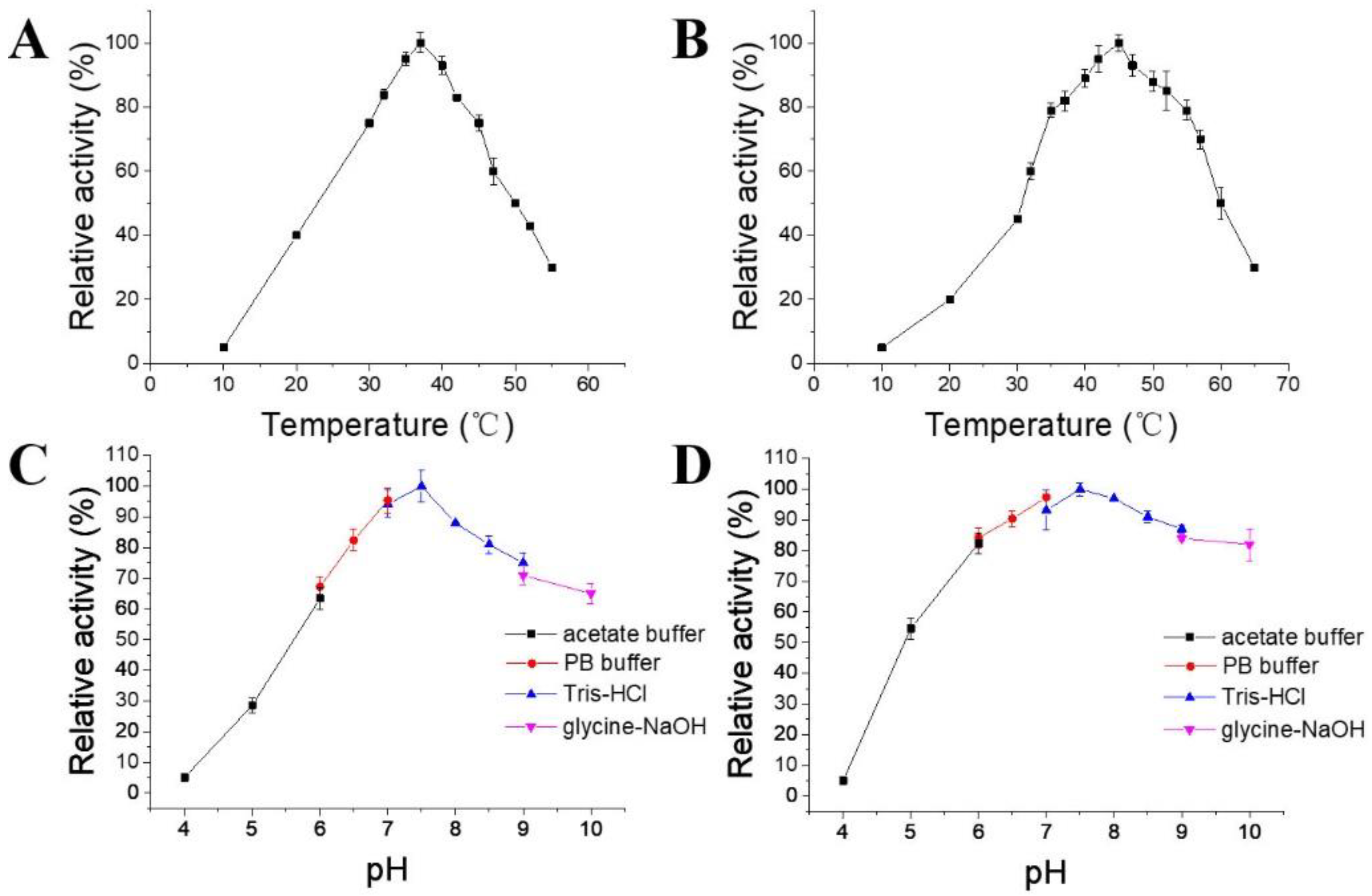
2.4. Determination of Optimum Temperature, Optimum pH, and Thermal Stability
2.5. Structure Modeling and MD Simulation
2.6. Application of ECA in French Fries and Bread
2.7. Assay of Concentration Assay
3. Results and Discussion
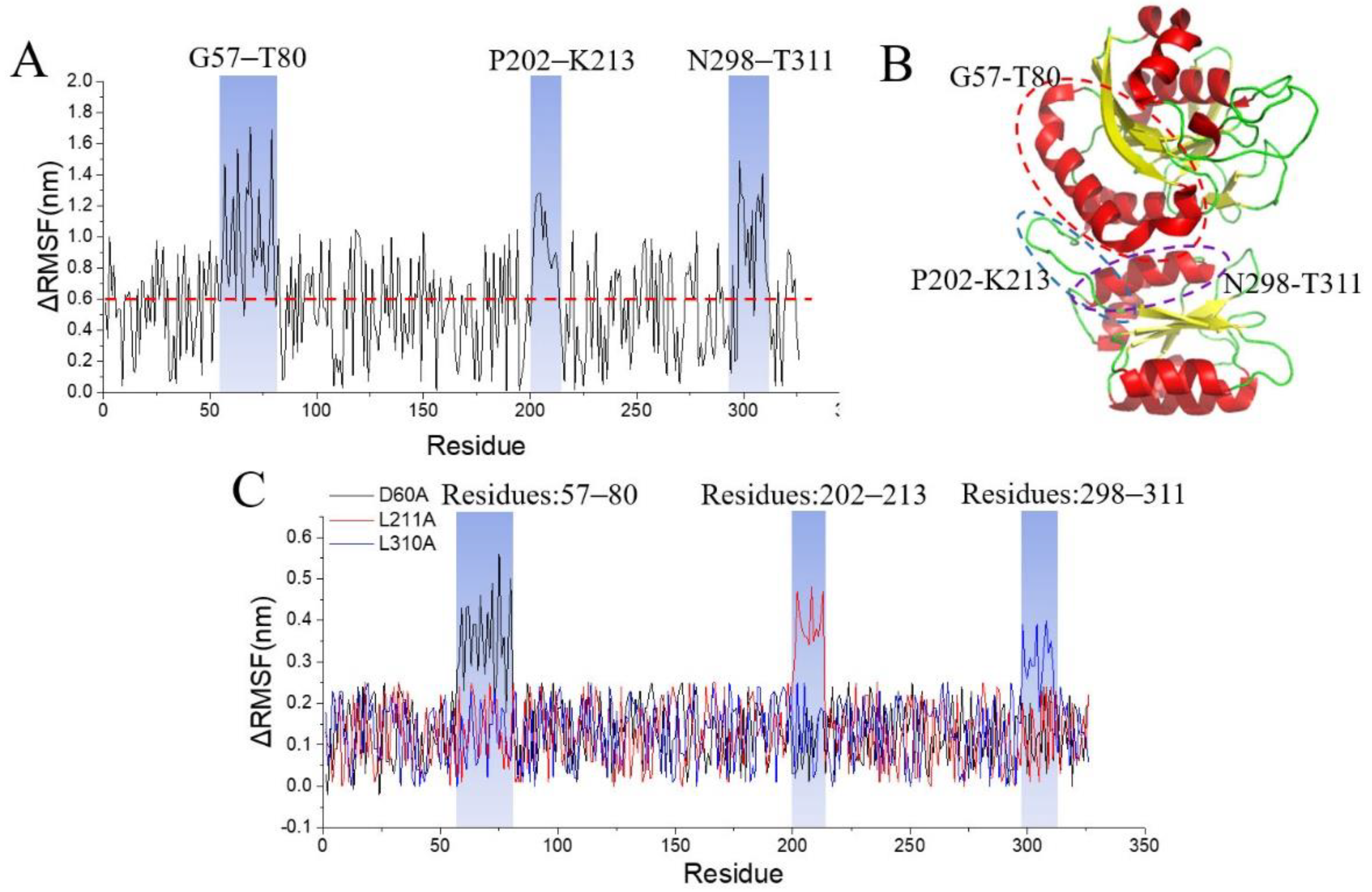
3.1. Identification of ECA Stability Key Domains
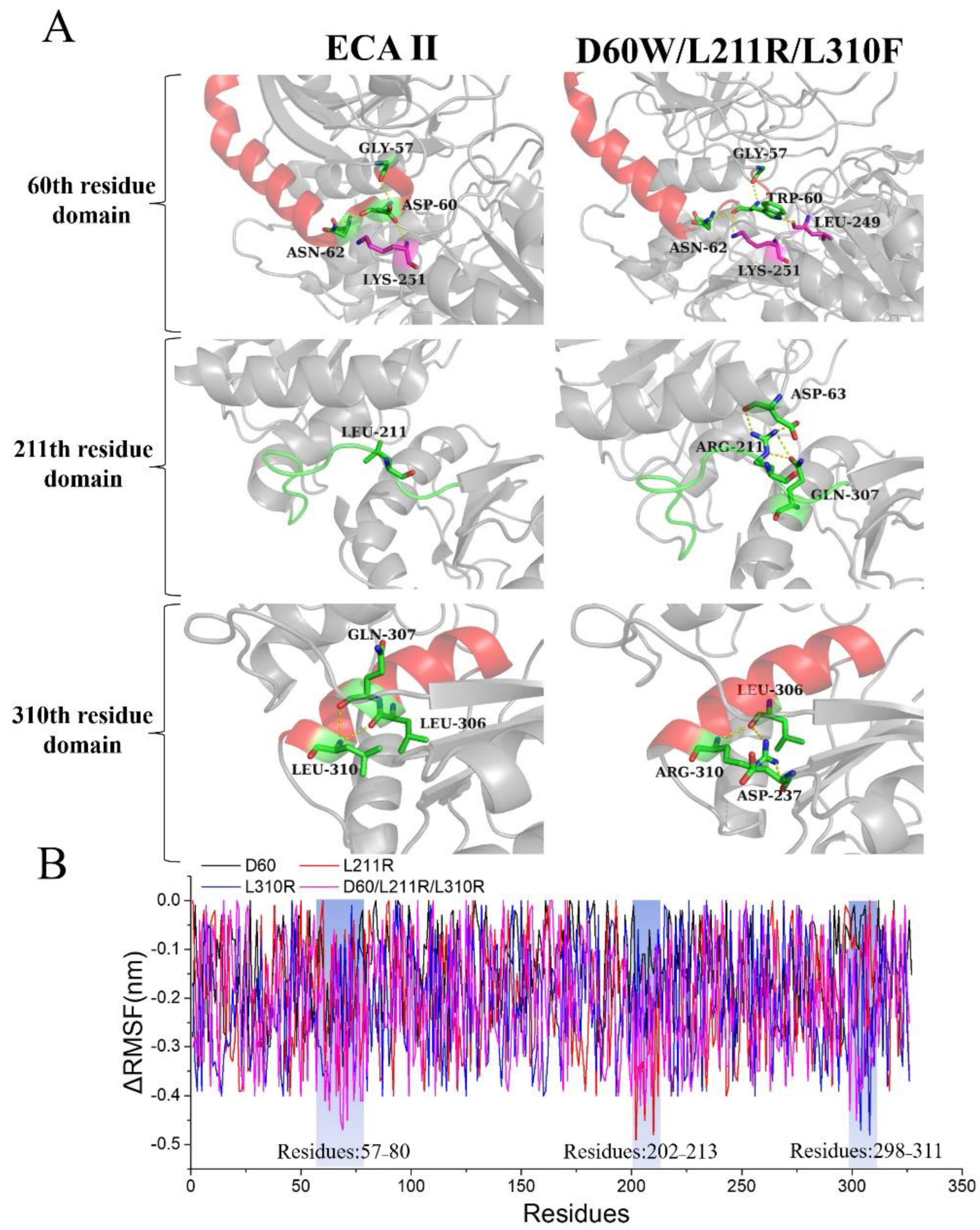
3.2. Identification of Stability Key Residues and Construction of Stability Mutation
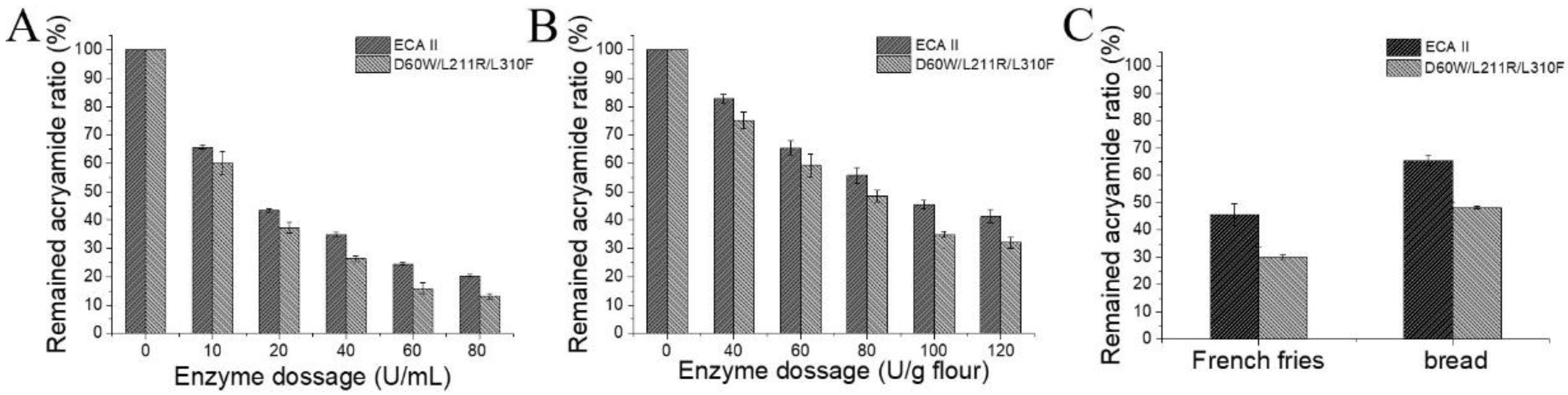
3.3. Effect of the Enzyme on Acrylamide Formation in French Fries and Bread under Controlled Conditions
3.4. Effect of the Enzyme on Acrylamide Formation in French Fries and Bread under Uncontrolled Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunha, M.C.; Dos Santos Aguilar, J.G.; de Melo, R.R.; Nagamatsu, S.T.; Ali, F.; de Castro, R.J.S.; Sato, H.H. Fungal L-asparaginase: Strategies for production and food applications. Food Res. Int. 2019, 126, 108658. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, H.; Zhang, W.; Xu, W.; Mu, W. Efficient control of acrylamide in French fries by an extraordinarily active and thermo-stable l-asparaginase: A lab-scale study. Food Chem. 2021, 360, 130046. [Google Scholar] [CrossRef]
- Exon, J.H. A review of the toxicology of acrylamide. J. Toxicol. Environ. Health B Crit. Rev. 2006, 9, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Daniali, G.; Jinap, S.; Hajeb, P.; Sanny, M.; Tan, C.P. Acrylamide formation in vegetable oils and animal fats during heat treatment. Food Chem. 2016, 212, 244–249. [Google Scholar] [CrossRef]
- Anese, M.; Nicoli, M.C.; Verardo, G.; Munari, M.; Mirolo, G.; Bortolomeazzi, R. Effect of vacuum roasting on acrylamide formation and reduction in coffee beans. Food Chem. 2014, 145, 168–172. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, Y.; Sun, G.; Wang, P.; Hu, X.; Chen, F. Role of glutathione on acrylamide inhibition: Transformation products and mechanism. Food Chem. 2020, 326, 126982. [Google Scholar] [CrossRef]
- Khalil, N.M.; Rodríguez-Couto, S.; El-Ghany, M.N.A. Characterization of Penicillium crustosum l-asparaginase and its acrylamide alleviation efficiency in roasted coffee beans at non-cytotoxic levels. Arch. Microbiol. 2021, 203, 2625–2637. [Google Scholar] [CrossRef]
- Kukurová, K.; Morales, F.J.; Bednáriková, A.; Ciesarová, Z. Effect of L-asparaginase on acrylamide mitigation in a fried-dough pastry model. Mol. Nutr. Food Res. 2009, 53, 1532–1539. [Google Scholar] [CrossRef]
- Jia, R.; Wan, X.; Geng, X.; Xue, D.; Xie, Z.; Chen, C. Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives. Microorganisms 2021, 9, 1659. [Google Scholar] [CrossRef]
- Bhagat, J.; Kaur, A.; Chadha, B.S. Single step purification of asparaginase from endophytic bacteria Pseudomonas oryzihabitans exhibiting high potential to reduce acrylamide in processed potato chips. Food Bioprod. Process. 2016, 99, 222–230. [Google Scholar] [CrossRef]
- Meghavarnam, A.K.; Janakiraman, S. Evaluation of acrylamide reduction potential of l-asparaginase from Fusarium culmorum (ASP-87) in starchy products. LWT 2018, 89, 32–37. [Google Scholar] [CrossRef]
- Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Reduction of acrylamide level through blanching with treatment by an extremely thermostable l-asparaginase during French fries processing. Extremophiles 2015, 19, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Liu, Y.; Mu, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a novel L-asparaginase from Paenibacillus barengoltzii being suitable for acrylamide reduction in potato chips and mooncakes. Int. J. Biol. Macromol. 2017, 96, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.M.; Xu, M.J.; He, B.B.; Rao, Z.M. Cloning, Expression, and Characterization of L-Asparaginase from a Newly Isolated Bacillus subtilis B11-06. J. Agric. Food Chem. 2013, 61, 9428–9434. [Google Scholar] [CrossRef]
- Farahat, M.G.; Amr, D.; Galal, A. Molecular cloning, structural modeling and characterization of a novel glutaminase-free L-asparaginase from Cobetia amphilecti AMI6. Int. J. Biol. Macromol. 2020, 143, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, H.V.; Kornbrust, B.A.; Østergaard, P.R.; Stringer, M.A. Evaluating the Potential for Enzymatic Acrylamide Mitigation in a Range of Food Products Using an Asparaginase from Aspergillus oryzae. J. Agric. Food Chem. 2009, 57, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, X.; Xu, S.; Xu, M.; Yang, T.; Wang, L.; Zhang, H.; Fang, H.; Osire, T.; Rao, Z. Insight into the thermostability of thermophilic L-asparaginase and non-thermophilic L-asparaginase II through bioinformatics and structural analysis. Appl. Microbiol. Biotechnol. 2019, 103, 7055–7070. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, M.; Ge, X.; Zhang, X.; Yang, T.; Xu, Z.; Rao, Z. Reengineering of the feedback-inhibition enzyme N-acetyl-l-glutamate kinase to enhance l-arginine production in Corynebacterium crenatum. J. Ind. Microbiol. Biotechnol. 2017, 44, 271–283. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Xu, S.; Zhang, H.; Xu, M.; Yang, T.; Wang, L.; Qian, H.; Zhang, H.; Fang, H.; et al. Simultaneous cell disruption and semi-quantitative activity assays for high-throughput screening of thermostable L-asparaginases. Sci. Rep. 2018, 8, 7915. [Google Scholar] [CrossRef]
- Roberts, J.; Burson, G.; Hill, J.M. New Procedures for Purification of l -Asparaginase with High Yield from Escherichia coli. J. Bacteriol. 1968, 95, 2117–2123. [Google Scholar] [CrossRef] [Green Version]
- Lubkowski, J.; Wlodawer, A. Geometric considerations support the double-displacement catalytic mechanism of l -asparaginase. Protein Sci. 2019, 28, 1850–1864. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.; Woetzel, N.; Meiler, J. Bcl::Cluster: A method for clustering biological molecules coupled with visualization in the Pymol Molecular Graphics System. In Proceedings of the 2011 IEEE 1st International Conference on Computational Advances in Bio and Medical Sciences (ICCABS), Orlando, FL, USA, 3–5 February 2011; Volume 2011, pp. 13–18. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Osés, G.; Osuna, S.; Gao, X.; Sawaya, M.; Gilson, L.; Collier, S.J.; Huisman, G.W.; Yeates, T.O.; Tang, Y.; Houk, K.N. The role of distant mutations and allosteric regulation on LovD active site dynamics. Nat. Chem. Biol. 2014, 10, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehr, D.D.; Nussinov, R.; Wright, P.E. The role of dynamic conformational ensembles in biomolecular recognition. Nat. Chem. Biol. 2009, 5, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehr, D.D.; Dyson, H.J.; Wright, P.E. An NMR Perspective on Enzyme Dynamics. Chem. Rev. 2006, 106, 3055–3079. [Google Scholar] [CrossRef]
- Alley, E.C.; Khimulya, G.; Biswas, S.; AlQuraishi, M.; Church, G.M. Unified rational protein engineering with sequence-based deep representation learning. Nat. Methods 2019, 16, 1315–1322. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins Struct. Funct. Bioinform. 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Kutzner, C.; van der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for Highly Efficient, Load-Balanced, and Scalable Molecular Simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Cui, W.; Cheng, Z.; Peplowski, L.; Liu, Z.; Kobayashi, M.; Zhou, Z. Improving the Thermostability and Catalytic Efficiency of the Subunit-Fused Nitrile Hydratase by Semi-Rational Engineering. ChemCatChem 2018, 10, 1370–1375. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Swain, A.L.; Jaskolski, M.; Housset, D.; Rao, J.K.M.; Wlodawer, A. Crystal-Structure of Escherichia coli L-Asparaginase, an Enzyme Used in Cancer-Therapy. Proc. Natl. Acad. Sci. USA 1993, 90, 1474–1478. [Google Scholar] [CrossRef] [Green Version]
- Tomar, R.; Sharma, P.; Srivastava, A.; Bansal, S.; Ashish; Kundu, B. Structural and functional insights into an archaeal L-asparaginase obtained through the linker-less assembly of constituent domains. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 3187–3197. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Rao, J.K.M.; Wlodawer, A.; Gribskov, M.R. A Left-Handed Crossover Involved in Amidohydrolase Catalysis—Crys-tal-Structure of Erwinia-Chrysanthemi L-Asparaginase with Bound L-Aspartate. FEBS Lett. 1993, 328, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, R.; Yang, T.; Liu, Q.; Zheng, J.; Wang, F.; Liu, F.; Xu, M.; Zhang, X.; Rao, Z. Relieving Allosteric Inhibition by Designing Active Inclusion Bodies and Coating of the Inclusion Bodies with Fe3O4 Nanomaterials for Sustainable 2-Oxobutyric Acid Production. ACS Catal. 2018, 8, 8889–8901. [Google Scholar] [CrossRef]
- Yang, B.; Wang, H.; Song, W.; Chen, X.; Liu, J.; Luo, Q.; Liu, L. Engineering of the Conformational Dynamics of Lipase to Increase Enantioselectivity. ACS Catal. 2017, 7, 7593–7599. [Google Scholar] [CrossRef]
- Brosnan, M.P.; Kelly, C.T.; Fogarty, W.M. Investigation of the Mechanisms of Irreversible Thermoinactivation of Bacil-lus-Stearothermophilus Alpha-Amylase. Eur. J. Biochem. 1992, 203, 225–231. [Google Scholar] [CrossRef]
- Vidya, J.; Ushasree, M.V.; Pandey, A. Effect of surface charge alteration on stability of l-asparaginase II from Escherichia sp. Enzym. Microb. Technol. 2014, 56, 15–19. [Google Scholar] [CrossRef]
- Niu, C.; Zhu, L.; Xu, X.; Li, Q. Rational Design of Disulfide Bonds Increases Thermostability of a Mesophilic 1,3-1,4-beta-Glucanase from Bacillus terquilensis. PLoS ONE 2016, 11, e0154036. [Google Scholar] [CrossRef] [Green Version]
- Long, S.; Zhang, X.; Rao, Z.; Chen, K.; Xu, M.; Yang, T.; Yang, S. Amino acid residues adjacent to the catalytic cavity of tetramer l -asparaginase II contribute significantly to its catalytic efficiency and thermostability. Enzym. Microb. Technol. 2016, 82, 15–22. [Google Scholar] [CrossRef]
- Keramat, J.; LeBail, A.; Prost, C.; Jafari, M. Acrylamide in Baking Products: A Review Article. Food Bioprocess Technol. 2011, 4, 530–543. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Van der Fels-Klerx, H.J.; Peters, R.J.B.; Van Boekel, M.A.J.S. Acrylamide and 5-hydroxymethylfurfural formation during baking of biscuits: Part I: Effects of sugar type. Food Chem. 2016, 192, 575–585. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Optimum Temperature (°C) | T(1/2, 50 °C) (Min) | Optimum pH | Km (μM) | Specific Activity (U/mg) |
|---|---|---|---|---|---|
| ECA II | 37 | 60 ± 5 | 7.5 | 18 ± 5 | 235 ± 21 |
| D60W | 40 | 85 ± 5 | 7.0 | 15 ± 6 | 245 ± 31 |
| L211R | 42 | 95 ± 5 | 7.5 | 24 ± 8 | 290 ± 33 |
| L310R | 40 | 80 ± 5 | 8.0 | 14 ± 6 | 217 ± 20 |
| D60W/L211R/L310R | 45 | 110 ± 10 | 7.5 | 26 ± 6 | 281 ± 29 |
| D60A | 35 | 40 ± 5 | 7.0 | 28 | 271 ± 29 |
| D60I | 37 | 50 ± 5 | 7.0 | 41 | 301 ± 41 |
| D60V | 37 | 60 ± 5 | 7.0 | 56 | 223 ± 21 |
| D60F | 40 | 75 ± 5 | 7.0 | 32 | 189 ± 31 |
| D60M | 37 | 60 ± 5 | 7.5 | 15 | 233 ± 33 |
| D60Q | 40 | 75 ± 5 | 7.5 | 66 | 199 ± 24 |
| D60T | 37 | 55 ± 5 | 7.0 | 90 | 211 ± 17 |
| D60N | 37 | 45 ± 5 | 7.0 | 55 | 273 ± 19 |
| D60Y | 40 | 70 ± 5 | 7.5 | 24 | 248 ± 24 |
| D60E | 37 | 60 ± 5 | 7.0 | 33 | 221 ± 44 |
| L211G | 35 | 40 ± 5 | 7.5 | 77 | 281 ± 37 |
| L211A | 35 | 45 ± 5 | 7.5 | 63 | 277 ± 21 |
| L211I | 37 | 55 ± 5 | 7.5 | 69 | 249 ± 11 |
| L211V | 37 | 45 ± 5 | 7.5 | 93 | 211 ± 18 |
| L211P | 40 | 75 ± 5 | 7.0 | 45 | 198 ± 55 |
| L211F | 40 | 65 ± 5 | 7.5 | 51 | 255 ± 23 |
| L211W | 37 | 65 ± 5 | 7.0 | 101 | 294 ± 36 |
| L211S | 40 | 60 ± 5 | 7.0 | 67 | 211 ± 19 |
| L211T | 37 | 45 ± 5 | 7.5 | 32 | 234 ± 12 |
| L211N | 37 | 55 ± 5 | 8.0 | 55 | 189 ± 11 |
| L211D | 40 | 65 ± 5 | 8.0 | 41 | 243 ± 28 |
| L211E | 40 | 70 ± 5 | 7.5 | 27 | 257 ± 31 |
| L211K | 40 | 70 ± 5 | 7.5 | 91 | 231 ± 37 |
| L310A | 37 | 45 | 7.5 | 33 | 221 ± 41 |
| L310I | 37 | 55 | 7.5 | 48 | 232 ± 21 |
| L310P | 40 | 70 | 7.5 | 19 | 203 ± 19 |
| L310F | 40 | 75 | 8.0 | 36 | 199 ± 23 |
| L310M | 37 | 60 | 8.0 | 52 | 198 ± 41 |
| L310W | 40 | 65 | 8.0 | 20 | 188 ± 28 |
| L310Q | 37 | 50 | 7.5 | 13 | 236 ± 23 |
| L310T | 37 | 55 | 7.5 | 54 | 210 ± 31 |
| L310C | 37 | 50 | 7.5 | 33 | 179 ± 51 |
| L310N | 37 | 55 | 7.5 | 64 | 198 ± 31 |
| L310Y | 40 | 65 | 7.5 | 88 | 204 ± 42 |
| L310K | 37 | 60 | 7.5 | 91 | 218 ± 16 |
| L310H | 37 | 65 | 7.5 | 31 | 221 ± 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, B.; Ma, P.; Fan, Y.; Guan, B.; Hu, Y.; Zhang, Y.; Yan, W.; Li, X.; Ni, Y. Construction of L-Asparaginase Stable Mutation for the Application in Food Acrylamide Mitigation. Fermentation 2022, 8, 218. https://doi.org/10.3390/fermentation8050218
Yuan B, Ma P, Fan Y, Guan B, Hu Y, Zhang Y, Yan W, Li X, Ni Y. Construction of L-Asparaginase Stable Mutation for the Application in Food Acrylamide Mitigation. Fermentation. 2022; 8(5):218. https://doi.org/10.3390/fermentation8050218
Chicago/Turabian StyleYuan, Bing, Pengfei Ma, Yuxuan Fan, Bo Guan, Youzhen Hu, Yan Zhang, Wenli Yan, Xu Li, and Yongqing Ni. 2022. "Construction of L-Asparaginase Stable Mutation for the Application in Food Acrylamide Mitigation" Fermentation 8, no. 5: 218. https://doi.org/10.3390/fermentation8050218
APA StyleYuan, B., Ma, P., Fan, Y., Guan, B., Hu, Y., Zhang, Y., Yan, W., Li, X., & Ni, Y. (2022). Construction of L-Asparaginase Stable Mutation for the Application in Food Acrylamide Mitigation. Fermentation, 8(5), 218. https://doi.org/10.3390/fermentation8050218