
Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ingredients, Chemicals, and Strain
2.2. Preparation of Fermented Oat Extract
2.3. Estimation of Viable L. plantarum B-59151
2.4. Determination of Total Phenolic Content (TPC)
2.5. Determination of Total Antioxidant Capacity (TAC) by the DPPH and ABTS Methods
2.6. Determination of the β-Glucan Content
2.7. Determination of the GABA Content
2.8. Animals and Experimental Design
2.8.1. Determination of Fasting Blood Glucose Level (FBG), Lipid Profile, Liver and Kidneys’ Functions
2.8.2. Determination of Oxidative Stress Biomarkers
2.9. Statistical Analysis
3. Results
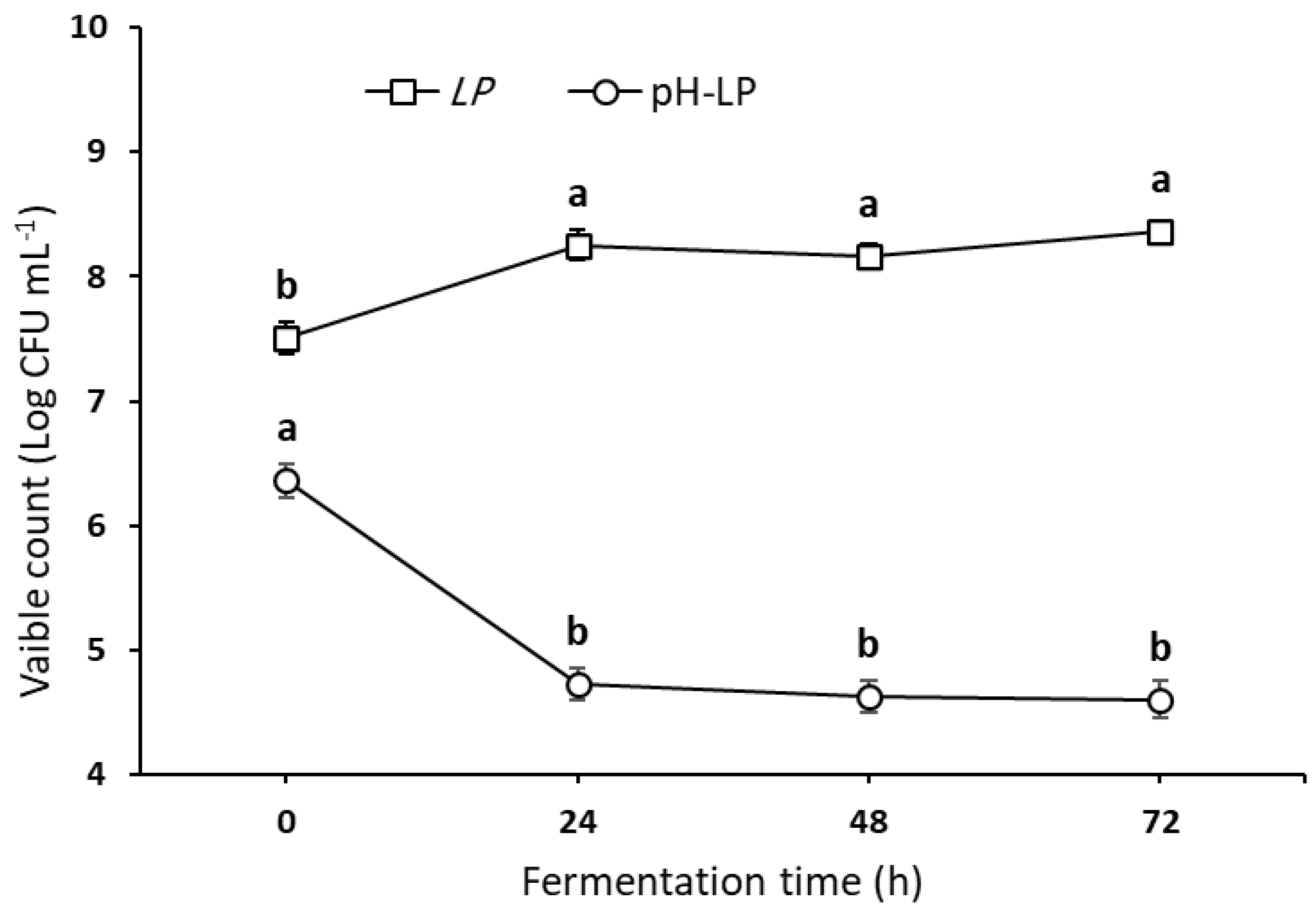
3.1. Survival of L. plantarum and Related pH Value
3.2. Phytochemicals and Antioxidant Capacity
3.3. Gamma-Aminobutyric Acid and β-Glucan Contents during Fermentation
3.4. The Hypoglycemic Efficiency
3.5. The Hypolipidemic Efficiency
3.6. The Liver’s Functions
3.7. The Kidneys’ Functions
3.8. Antioxidant Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- International Diabetes Federation. IDF Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2021; Volume 102, pp. 147–148. [Google Scholar]
- Cole, J.B.; Florez, J.C. Genetics of Diabetes Mellitus and Diabetes Complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Cai, X.; Xu, M.; Li, Y. Effect of Oat Intake on Glycaemic Control and Insulin Sensitivity: A Meta-Analysis of Randomised Controlled Trials. Br. J. Nutr. 2014, 112, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.F.; Lobato, R.V.; de Araújo, T.V.; Zangerônimo, M.G.; de Sousa, R.V.; Pereira, L.J. Effect of beta-Glucans in the Control of Blood Glucose Levels of Diabetic Patients: A Systematic Review. Nutr. Hosp. 2015, 31, 170–177. [Google Scholar]
- Shen, X.L.; Zhao, T.; Zhou, Y.; Shi, X.; Zou, Y.; Zhao, G. Effect of Oat β-glucan Intake on Glycaemic Control and Insulin Sensitivity of Diabetic patients: A Meta-Analysis of Randomized Controlled Trials. Nutrients 2016, 8, 39. [Google Scholar] [PubMed] [Green Version]
- Ahmad, M.; Gul-Zaffar; Dar, Z.A.; Habib, M. A review on Oat (Avena sativa L.) as A Dual-Purpose Crop. Sci. Res. Essay 2014, 9, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, S.; Singh, R.; Tomar, S.K. Nutritional and Functional Properties of Oats: An Update. J. Innov. Biol. 2014, 1, 3–14. [Google Scholar]
- Angelov, A.; Yaneva-Marinova, T.; Gotcheva, V. Oats as A Matrix of Choice for Developing Fermented Functiona Beverages. J. Food Sci. Technol. 2018, 55, 2351–2360. [Google Scholar] [CrossRef]
- Boz, H. Phenolic Amides (avenanthramides) in Oats—A Review. Czech J. Food Sci. 2015, 33, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Meydani, M. Potential Health Benefits of Avenanthramides of Oats. Nutr. Rev. 2009, 67, 731–735. [Google Scholar] [CrossRef]
- Cai, S.; Gao, F.; Zhang, X.; Wang, O.; Wu, W.; Zhu, S.; Zhang, D.; Zhou, F.; Ji, B. Evaluation of γ-aminobutyric Acid, Phytate and Antioxidant Activity of Tempeh-like Fermented Oats (Avena sativa L.) Prepared with Different Filamentous Fungi. J. Food Sci. Technol. 2014, 51, 2544–2551. [Google Scholar] [CrossRef] [Green Version]
- Sunderam, V.; Mohammed, S.S.S.; Madhavan, Y.; Dhinakaran, M.; Sampath, S.; Patteswaran, N.; Thangavelu, L.; Lawrence, A.V. Free Radical Scavenging Activity and Cytotoxicity Study of Fermented Oats (Avena sativa). Int. J. Res. Pharm. Sci. 2020, 11, 1259–1262. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes Through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, X.; Dong, Y.; Shi, L.; Xu, T.; Wu, F. The Anti-Obesity Effect of Fermented Barley Extracts with Lactobacillus plantarum dy-1 and Saccharomyces cerevisiae in Diet-Induced Obese Rats. Food Funct. 2017, 8, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Saraphanchotiwitthaya, A.; Sripalakit, P. Production of γ-aminobutyric Acid from Red Kidney Bean and Barley Grain Fermentation by Lactobacillus brevis TISTR 860. Biocatal. Agric. Biotechnol. 2018, 16, 49–53. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of GABA (γ-aminobutyric acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Adeghate, E.; Ponery, A.S. GABA in the Endocrine Pancreas: Cellular Localization and Function in Normal and Diabetic Rats. Tissue Cell 2002, 34, 1–6. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Prasanth, M.I.; Chaiyasut, C. A mini Review on Antidiabetic Properties of Fermented Foods. Nutrients 2018, 10, 1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindner, J.D.D.; Valentina, B. New Insights into Food Fermentation. Foods 2022, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, D.H.; Kang, H.J.; Shin, M.; Yang, S.-Y.; Yang, J.; Jung, Y.H. Enhanced Production of γ-aminobutyric Acid (GABA) using Lactobacillus plantarum EJ2014 with Simple Medium Composition. LWT Food Sci. Technol. 2021, 137, 110443. [Google Scholar] [CrossRef]
- De Vries, M.C.; Vaughan, E.E.; Kleerebezem, M.; de Vos, W.M. Lactobacillus plantarum—Survival, Functional and Potential Probiotic Properties in the Human Intestinal Tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Won, G.; Choi, S.-I.; Park, N.; Kim, J.-E.; Kang, C.-H.; Kim, G.-H. In Vitro Antidiabetic, Antioxidant Activity, and Probiotic Activities of Lactiplantibacillus plantarum and Lacticaseibacillus paracasei Strains. Curr. Microbiol. 2021, 78, 3181–3191. [Google Scholar] [CrossRef]
- Li, X.; Cai, X.; Ma, X.; Jing, L.; Gu, J.; Bao, L.; Li, J.; Xu, M.; Zhang, Z.; Li, Y. Short-and Long-Term Effects of Wholegrain OAT Intake on Weight Management and Glucolipid Metabolism in Overweight Type-2 Diabetics: A Randomized Control Trial. Nutrients 2016, 8, 549. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, C.G.; Reinheimer, J.A. Culture Media for the Enumeration of Bifidobacterium bifidum and Lactobacillus acidophilus in the Presence of Yoghurt Bacteria. Int. Dairy J. 1999, 9, 497–505. [Google Scholar] [CrossRef]
- Nsimba, R.Y.; Kikuzaki, H.; Konishi, Y. Antioxidant Activity of Various Extracts and Fractions of Chenopodium quinoa and Amaranthus spp. seeds. Food Chem. 2008, 106, 760–766. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, H.; Chen, J.; Fan, W.; Dong, J.; Kong, W.; Sun, J.; Cao, Y.; Cai, G. Evolution of Phenolic Compounds and Antioxidant Activity During Malting. J. Agr. Food Chem. 2007, 55, 10994–11001. [Google Scholar] [CrossRef]
- Yuwa-amornpitak, T.; Butkhup, L.; Yeunyaw, P.-N. Amino Acids and Antioxidant Activities of Extracts From Wild Edible Mushrooms From A Community Forest in the Nasrinual District, Maha Sarakham, Thailand. Food Sci. Technol. 2020, 40, 712–720. [Google Scholar] [CrossRef]
- Shiju, T.M.; Rajesh, N.G.; Viswanathan, P. Renoprotective Effect of Aged Garlic Extract in Streptozotocin-Induced Diabetic Rats. Indian J. Pharmacol. 2013, 45, 18. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose Translation from Animal to Human Studies Revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-density Lipoprotein CHOLESTEROL in Plasma, without use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Nwagha, U.; Ikekpeazu, E.; Ejezie, F.; Neboh, E.; Maduka, I. Atherogenic Index of Plasma as Useful Predictor of Cardiovascular Risk Among Postmenopausal Women in Enugu, Nigeria. Afr. Health Sci. 2010, 10, 248–252. [Google Scholar]
- Beutler, E. Improved Method for the Determination of Blood Glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Steel, R.G. Principles and Procedures of Statistics A Biometrical Approach, 3rd ed.; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Schofield, J.D.; Liu, Y.; Rao-Balakrishna, P.; Malik, R.A.; Soran, H. Diabetes dyslipidemia. Diabetes Ther. 2016, 7, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Gohari, S.T.; El-Batawy, O.I.; Mahdy, S.M. Biological Attributes of Unfermented and Fermented Oat Milk Compared to Cow Milk in Alloxan-Induced Diabetic Rats. J. Food Dairy Sci. 2019, 10, 61–69. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, Q.P.; Duan, J.L.; Tian, C.R. Dynamic Changes in γ-aminobutyric Acid and Glutamate Decarboxylase Activity in Oats (Avena nuda L.) During Steeping and Germination. J. Agric. Food Chem. 2010, 58, 9759–9763. [Google Scholar] [CrossRef]
- Bozbulut, R.; Sanlier, N. Promising Effects of β-glucans on Glyceamic Control in Diabetes. Trends Food Sci. Technol. 2019, 83, 159–166. [Google Scholar] [CrossRef]
- Chen, L.; Wu, D.; Schlundt, J.; Conway, P.L. Development of a Dairy-Free Fermented Oat-Based Beverage With Enhanced Probiotic and Bioactive Properties. Front. Microbiol. 2020, 11, 609734. [Google Scholar] [CrossRef]
- Angelov, A.; Gotcheva, V.; Kuncheva, R.; Hristozova, T. Development of a New Oat-based Probiotic Drink. Int. J. Food Microbiol. 2006, 112, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Cai, S.; Nout, R.M.; Wang, Y.; Xia, Y.; Li, Y.; Ji, B. Production of Oat-based Synbiotic Beverage by Two-Stage Fermentation with Rhizopus oryzae and Lactobacillus acidophilus. J. Food Agric. Environ. 2012, 10, 175–179. [Google Scholar]
- Jaskari, J.; Kontula, P.; Siitonen, A.; Jousimies-Somer, H.; Mattila-Sandholm, T.; Poutanen, K. Oat β-glucan and Xylan Hydrolysates as Selective Substrates for Bifidobacterium and Lactobacillus strains. Appl. Microbiol. Biotechnol. 1998, 49, 175–181. [Google Scholar] [CrossRef]
- Giri, B.; Dey, S.; Das, T.; Sarkar, M.; Banerjee, J.; Dash, S.K. Chronic Hyperglycemia Mediated Physiological Alteration and Metabolic Distortion Leads to Organ Dysfunction, Infection, Cancer Progression and other Pathophysiological Consequences: An Update on Glucose Toxicity. Biomed. Pharmacother. 2018, 107, 306–328. [Google Scholar] [CrossRef] [PubMed]
- Ademiluyi, A.O.; Oboh, G. Attenuation of Oxidative Stress and Hepatic Damage by some Fermented Tropical Legume Condiment Diets in Sreptozotocin–induced Diabetes in Rats. Asian Pac. J. Trop. Med. 2012, 5, 692–697. [Google Scholar] [CrossRef] [Green Version]
- Zulkawi, N.; Ng, K.H.; Zamberi, N.R.; Yeap, S.K.; Satharasinghe, D.A.; Tan, S.W.; Ho, W.Y.; Rashid, N.Y.A.; Lazim, M.I.M.; Jamaluddin, A. Antihyperglycemic and Anti-inflammatory Effects of Fermented Food Paste in High-Fat Diet and Streptozotocin-Challenged Mice. Drug Des. Devel. Ther. 2018, 12, 1373. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, A.; Kalaivani, M. Protective Effect of Monascus fermented Rice Against STZ-induced Diabetic Oxidative Stress in Kidney of Rats. J. Food Sci. Technol. 2015, 52, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.M.; Yusof, H.M.; Long, K.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Antioxidant and Hepatoprotective Effect of Aqueous Extract of Germinated and Fermented Mung Bean on Ethanol-Mediated Liver Damage. BioMed. Res. Int. 2013, 2013, 693613. [Google Scholar]
- Al-Qabba, M.M.; El-Mowafy, M.A.; Althwab, S.A.; Alfheeaid, H.A.; Aljutaily, T.; Barakat, H. Phenolic Profile, Antioxidant Activity, and Ameliorating Efficacy of Chenopodium quinoa Sprouts against CCl4-Induced Oxidative Stress in Rats. Nutrients 2020, 12, 2904. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Bulmer, A.C.; Singh, I. A Review of the Mechanisms and Effectiveness of Dietary Polyphenols in Reducing Oxidative Stress and Thrombotic Risk. J. Hum. Nutr. Diet. 2014, 27, 1–21. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Alharbi, Y.M.; Sakr, S.S.; Albarrak, S.M.; Almundarij, T.I.; Barakat, H.; Hassan, M.F.Y. Antioxidative, Antidiabetic, and Hypolipidemic Properties of Probiotic-Enriched Fermented Camel Milk Combined with Salvia officinalis Leaves Hydroalcoholic Extract in Streptozotocin-Induced Diabetes in Rats. Antioxidants 2022, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Ha, B.J. Antihyperglycemic and Antihyperlipidemic Effects of Fermented Rhynchosia nulubilis in Alloxan-induced Diabetic Rats. Toxicol. Res. 2013, 29, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Ding, X.W.; Zhong, L.Y.; Zhu, C.; Nie, P.; Song, L.H. Beneficial Effects of Lactobacillus-Fermented Black Barley on High Fat Diet-Induced Fatty Liver in Rats. Food Funct. 2021, 12, 6526–6539. [Google Scholar] [CrossRef]


{kind=link}
| Group | Experimental Treatment |
|---|---|
| NR | Normal non-diabetic rats |
| STZ | Untreated diabetic rats |
| STZ+MET | Diabetic rats + metformin (50 mg kg−1 BW) |
| STZ+UFOE | Diabetic rats + UFOE (7 mL) |
| STZ+LFOE | Diabetic rats + LFOE (7 mL) |
| Item | Fermentation Time | |||
|---|---|---|---|---|
| 0 h | 24 h | 48 h | 72 h | |
| TPC (mg GAE g−1) | 0.70 ± 0.08 b | 0.74 ± 0.07 b | 0.94 ± 0.15 a,b | 1.04 ± 0.08 a |
| DPPH (µmol of TE g−1) | 2.14 ± 0.09 b | 2.22 ± 0.11 b | 3.19 ± 0.12 a,b | 3.56 ± 0.21 a |
| ABTS (µmol of TE g−1) | 3.47 ± 0.29 c | 3.89 ± 0.13 c | 4.58 ± 0.31 b | 5.27 ± 0.14 a |
| Item | Fermentation Time | |||
|---|---|---|---|---|
| 0 h | 24 h | 48 h | 72 h | |
| GABA * (mg 100 g−1) | 4.12 ± 0.14 b | 4.77 ± 0.17 b | 6.10 ± 0.52 a,b | 7.35 ± 0.40 a |
| β-glucan (g 100 g−1) | 2.62 ± 0.02 a | 2.60 ± 0.01 a | 2.56 ± 0.03 a | 2.45 ± 0.06 b |
| Groups * | RBG | FBG | ||
|---|---|---|---|---|
| Weak-0 | Weak-3 | Weak-6 | ||
| NR | 113.83 ± 3.82 c,A | 114.33 ± 3.06 d,A | 110.67 ± 2.55 d,A | 88.79 ± 4.28 b |
| STZ | 314.17 ± 33.65 a,A | 386.33 ± 36.6 a,A | 359.5 ± 49.11 a,A | 260.84 ± 10.35 a |
| STZ+MET | 284.33 ± 34.90 a,b,A | 238.67 ± 30.01 c,B | 201.5 ± 18.49 b,B | 89.96 ± 2.5 b |
| STZ+UFOE | 260.50 ± 27.63 b,A | 288.17 ± 44.38 b,A | 232.5 ± 25.24 b,B | 96.54 ± 6.97 b |
| STZ+LFOE | 308.50 ± 22.13 a,A | 244.83 ± 28.82 c,B | 179.33 ± 32.09 c,C | 91.00 ± 8.09 b |
| Groups * | Lipid Profile Parameters | |||||
|---|---|---|---|---|---|---|
| TG | CHO | HDL | LDL | VLDL | AI | |
| NR | 70.71 ± 3.01 c | 91.15 ± 7.88 b | 38.07 ± 5.56 a | 40.20 ± 9.79 b | 14.14 ± 0.60 c | 0.30 ± 0.08 c |
| STZ | 113.18 ± 5.44 a | 141.78 ± 11.25 a | 26.71 ± 4.07 b | 92.43 ± 14.04 a | 22.64 ± 1.09 a | 0.66 ± 0.07 a |
| STZ+MET | 97.39 ± 8.71 a,b | 103.18 ± 12.32 b | 36.37 ± 6.71 a | 47.34 ± 11.32 b | 19.48 ± 1.74 a,b | 0.48 ± 0.10 b |
| STZ+UFOE | 92.17 ± 5.85 b | 98.53 ± 5.54 b | 39.77 ± 3.72 a | 40.84 ± 4.41 b | 18.43 ± 1.17 b | 0.33 ± 0.05 b,c |
| STZ+LFOE | 81.90 ± 3.95 b,c | 104.93 ± 10.79 b | 40.34 ± 5.19 a | 48.96 ± 12.95 b | 16.38 ± 0.79 b | 0.37 ± 0.05 b,c |
| Groups * | Liver’s Functions | |||
|---|---|---|---|---|
| ALT (U L−1) | AST (U L−1) | ALP (U L−1) | T. Bili (mg dL−1) | |
| NR | 44.50 ± 2.57 b | 58.25 ± 4.43 b | 74.78 ± 27.77 b | 0.62 ± 0.15 b |
| STZ | 68.20 ± 11.59 a | 90.90 ± 10.39 a | 149.05 ± 20.83 a | 0.97 ± 0.18 a |
| STZ+MET | 47.79 ± 1.80 b | 68.84 ± 5.74 b | 101.06 ± 10.58 a,b | 0.79 ± 0.08 a,b |
| STZ+UFOE | 55.09 ± 2.74 a,b | 76.07 ± 1.57 b | 88.22 ± 6.38 b | 0.71 ± 0.04 a,b |
| STZ+LFOE | 54.02 ± 4.64 a,b | 70.16 ± 2.44 b | 76.85 ± 7.74 b | 0.65 ± 0.08 a,b |
| Group * | Kidneys’ Functions | |||||
|---|---|---|---|---|---|---|
| T. Protein (g dL−1) | Albumin (g dL−1) | Globulin (g dL−1) | Creatinine (mg dL−1) | Urea (mg dL−1) | BUN (mg dL−1) | |
| NR | 8.81 ± 0.17 b | 4.11 ± 0.24 a | 4.70 ± 0.18 a | 0.77 ± 0.03 c | 34.18 ± 6.74 b | 16.07 ± 3.17 b |
| STZ | 7.20 ± 0.28 d | 3.53 ± 0.08 a | 3.67 ± 0.30 a | 1.28 ± 0.05 a | 68.66 ± 10.51 a | 32.27 ± 4.94 a |
| STZ+MET | 7.79 ± 0.35 c,d | 4.01 ± 0.31 a | 3.78 ± 0.57 a | 0.96 ± 0.07 b | 47.31 ± 6.04 b | 22.24 ± 2.84 b |
| STZ+UFOE | 9.94 ± 0.32 a | 4.01 ± 0.30 a | 4.98 ± 0.38 a | 0.88 ± 0.06 b,c | 41.04 ± 5.91 b | 19.29 ± 2.78 b |
| STZ+LFOE | 8.23 ± 0.40 b,c | 3.73 ± 0.29 a | 4.50 ± 0.51 a | 0.81 ± 0.07 b,c | 39.70 ± 6.86 b | 18.66 ± 3.22 b |
| Group * | Antioxidant Biomarkers | |||
|---|---|---|---|---|
| GSH (µg dL−1) | MDA (nmol mL−1) | CAT (U L−1) | SOD (U L−1) | |
| NR | 75.11 ± 9.02 a | 14.63 ± 0.5 c | 74.99 ± 7.14 a | 59.88 ± 3.85 a |
| STZ | 36.17 ± 4.63 c | 20.63 ± 0.37 a | 48.43 ± 5.69 c,d | 35.63 ± 2.52 c |
| STZ+MET | 50.08 ± 5.05 b,c | 17.48 ± 0.63 b | 58.18 ± 4.84 c | 44.45 ± 3.2 b,c |
| STZ+UFOE | 45.56 ± 2.22 b,c | 16.67 ± 0.35 b | 62.81 ± 6.84 b | 53.56 ± 2.14 a,b |
| STZ+LFOE | 55.97 ± 2.71 b | 16.67 ± 0.29 b | 69.79 ± 5.05 a | 51.20 ± 4.10 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algonaiman, R.; Alharbi, H.F.; Barakat, H. Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats. Fermentation 2022, 8, 267. https://doi.org/10.3390/fermentation8060267
Algonaiman R, Alharbi HF, Barakat H. Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats. Fermentation. 2022; 8(6):267. https://doi.org/10.3390/fermentation8060267
Chicago/Turabian StyleAlgonaiman, Raya, Hend F. Alharbi, and Hassan Barakat. 2022. "Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats" Fermentation 8, no. 6: 267. https://doi.org/10.3390/fermentation8060267
APA StyleAlgonaiman, R., Alharbi, H. F., & Barakat, H. (2022). Antidiabetic and Hypolipidemic Efficiency of Lactobacillus plantarum Fermented Oat (Avena sativa) Extract in Streptozotocin-Induced Diabetes in Rats. Fermentation, 8(6), 267. https://doi.org/10.3390/fermentation8060267