
Probiotic and Antioxidant Potential of the Lactobacillus Spp. Isolated from Artisanal Fermented Pickles


 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Lactic Acid Bacteria
2.2. Molecular Identification of Bacterial Isolates
2.3. Probiotic Characterization
2.3.1. Acid Tolerance
2.3.2. Bile Tolerance
2.3.3. Cell Auto-Aggregation
2.4. Cell Surface Hydrophobicity
2.5. Determination of Pepsin and Pancreatin Resistance
2.6. Lysozyme Resistance
2.7. Amylolytic Activity
2.8. Cellulolytic Activity
2.9. Proteolytic Activity
2.10. Lipolytic Activity
2.11. Assessment of Antibiotic Susceptibility
2.12. Antagonistic Activity
2.13. Antioxidant Activity
2.14. Statistical Analysis
3. Results
3.1. Purification and Biochemical Characterization of Bacterial Isolates
3.2. Enzymatic Potential of Lactobacillus Strains
3.3. Antibiotic Suscenptibility
3.4. Determination of Cell Viability under Different Probiotic Parameters
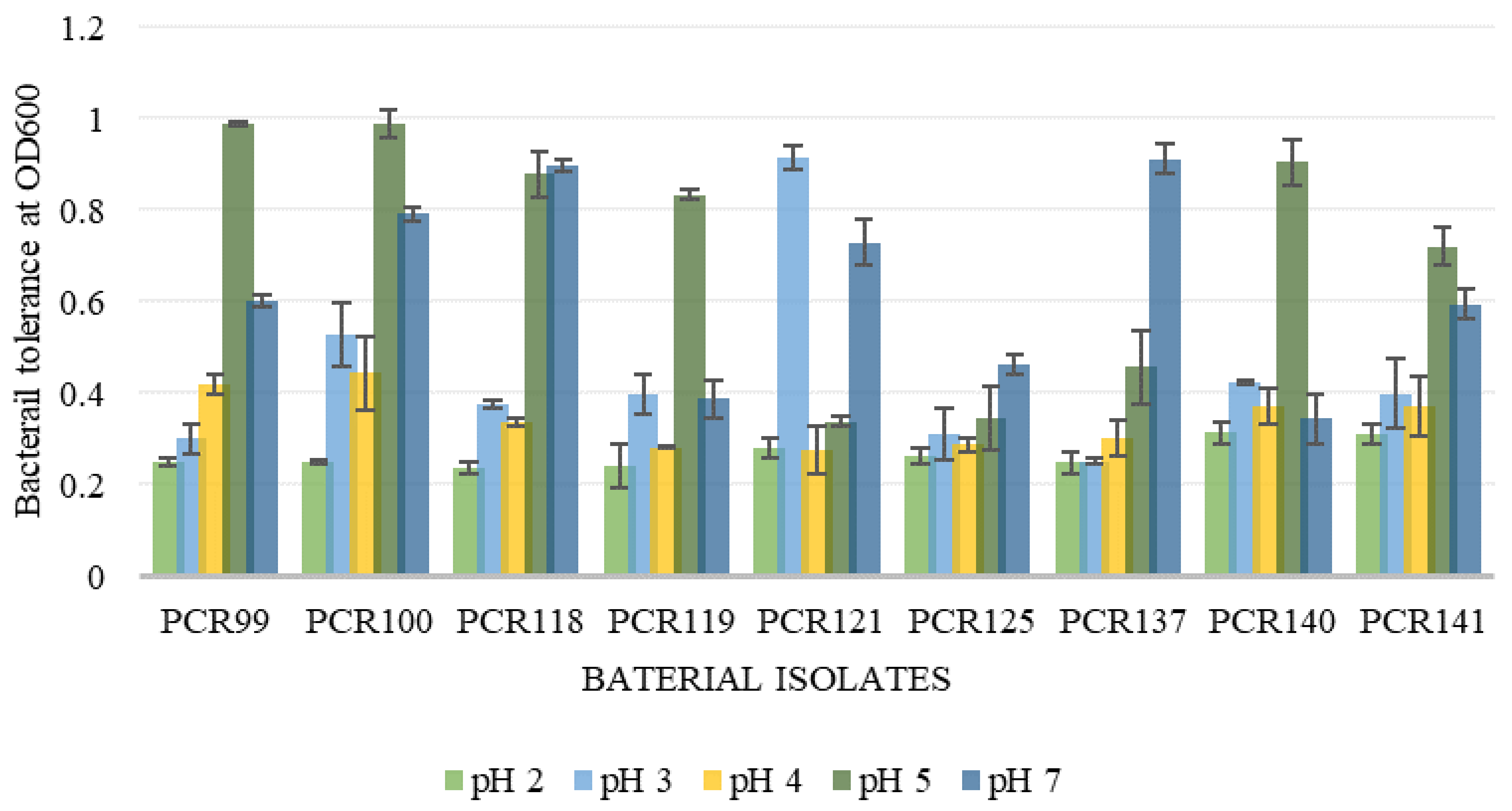
3.4.1. pH Tolerance Assay
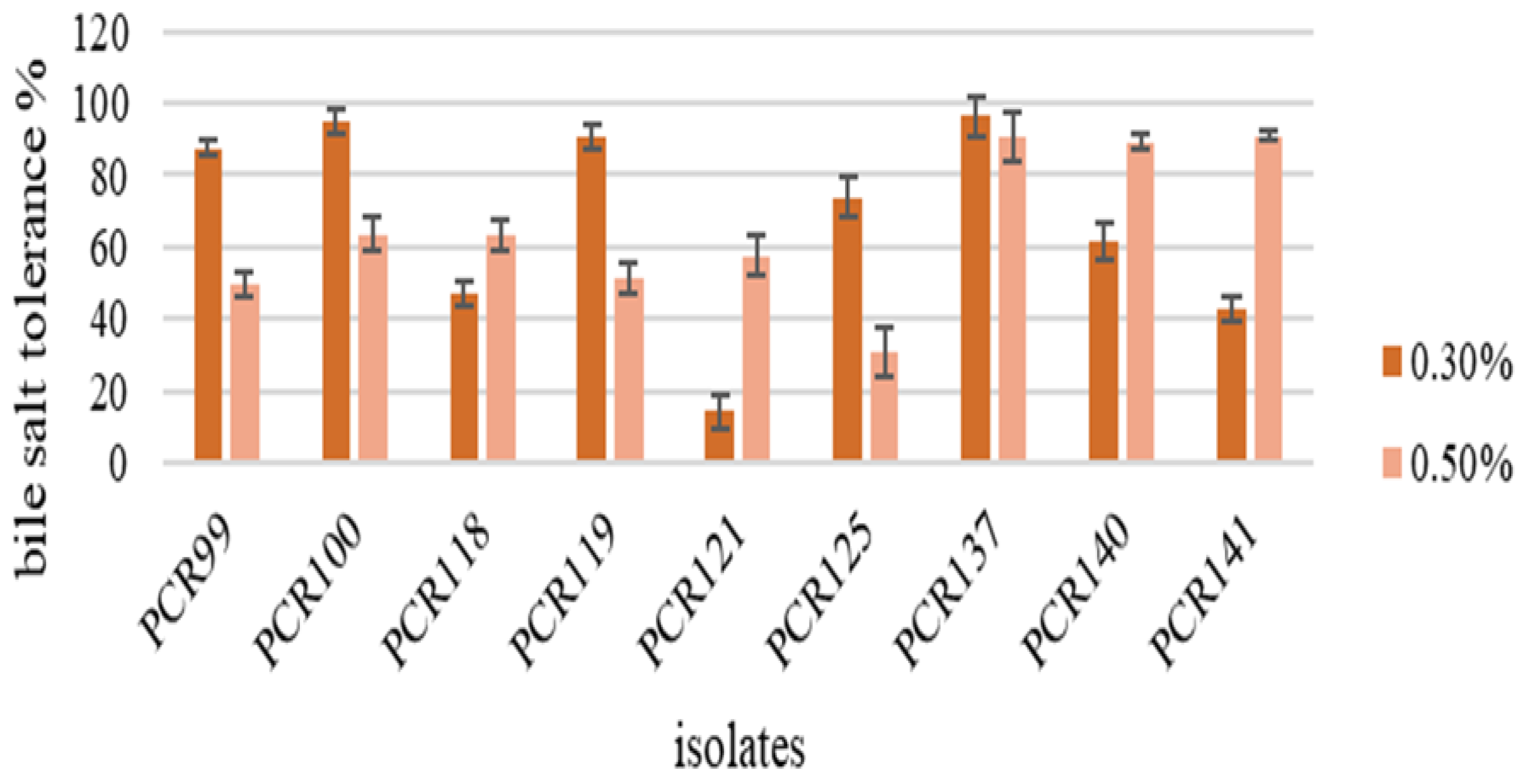
3.4.2. Bile Salt Tolerance Assay
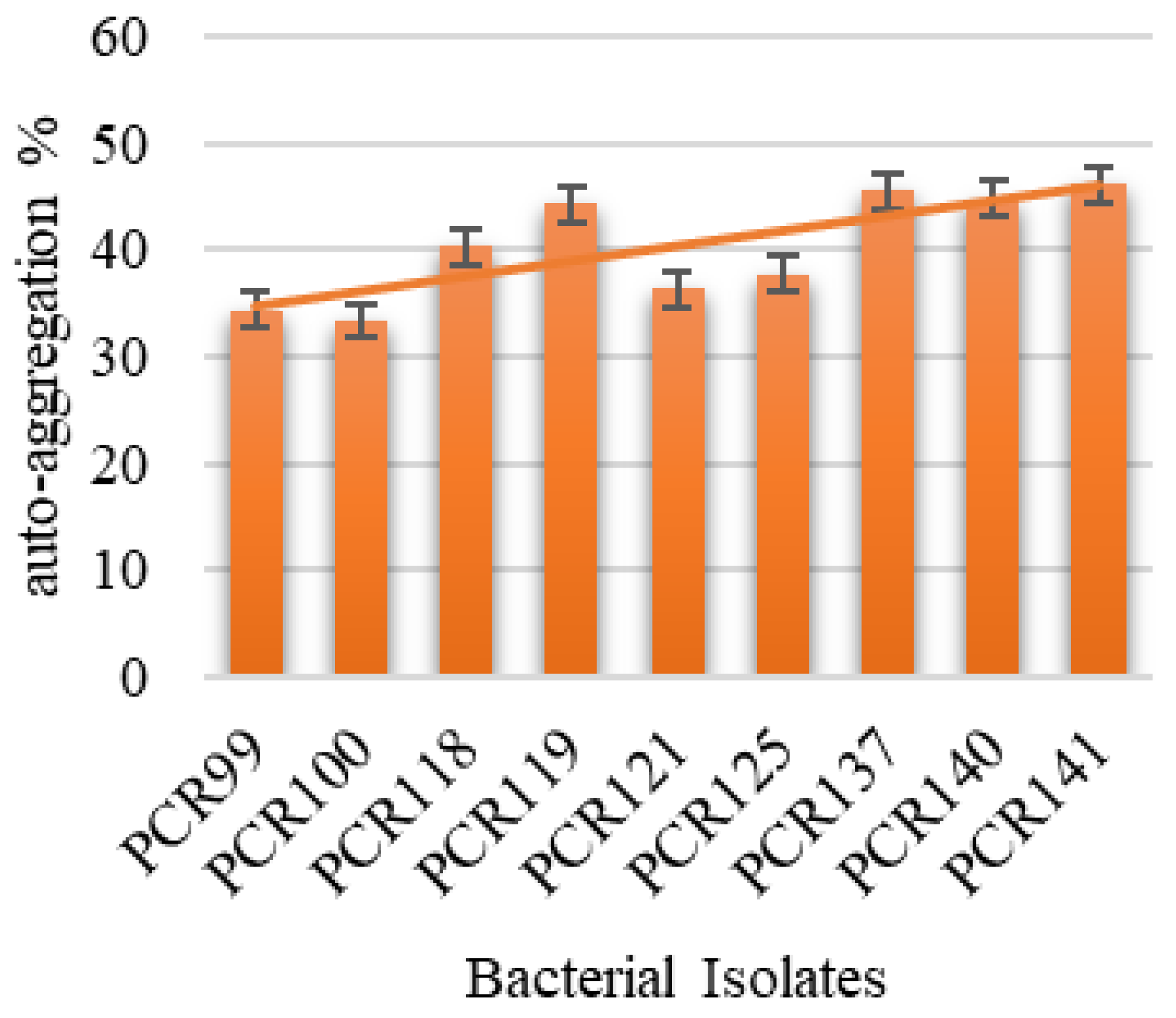
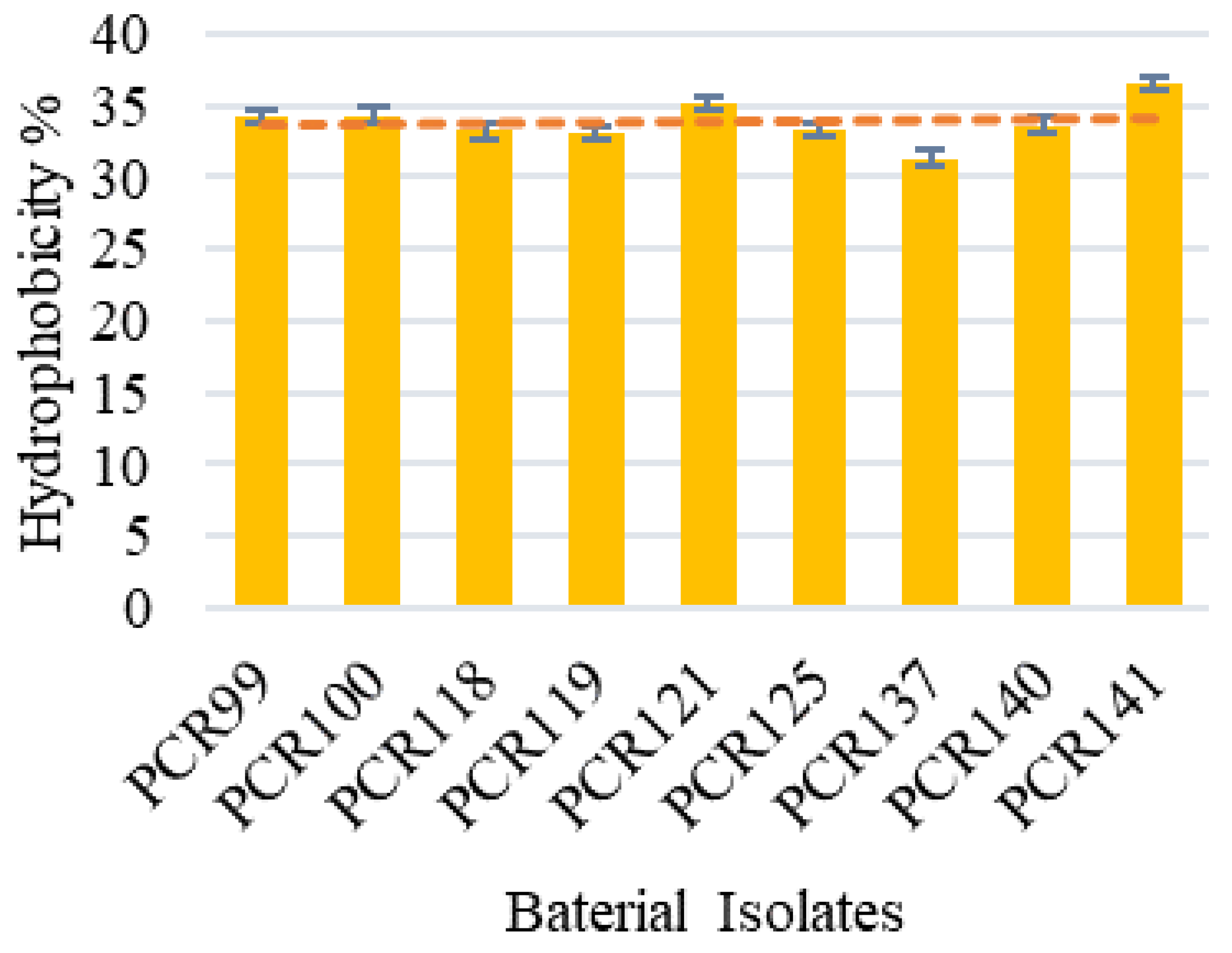
3.4.3. Auto-Aggregation and Cell Surface Hydrophobicity Determination
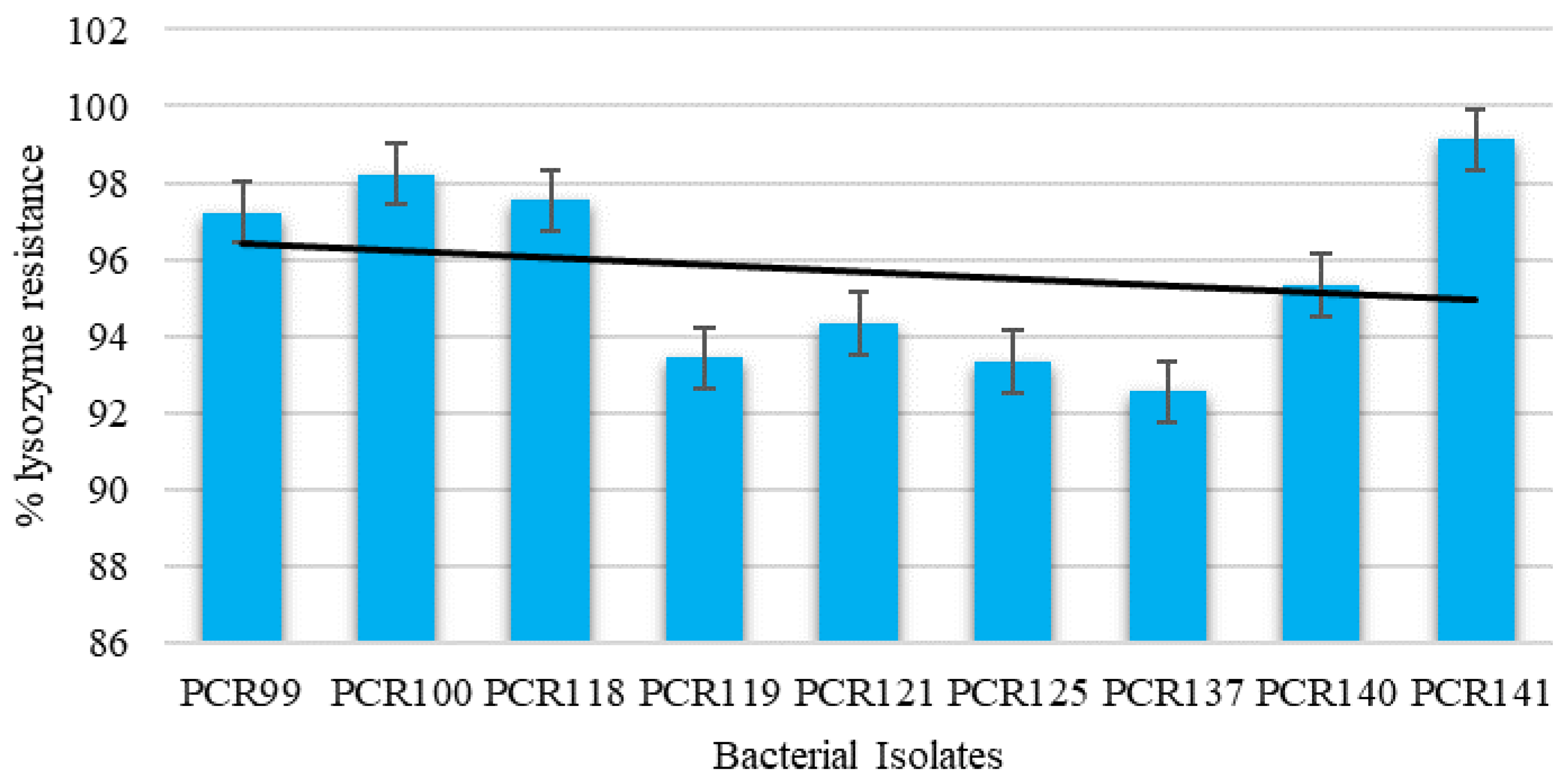
3.4.4. Effect of Lysozyme on Cell Viability
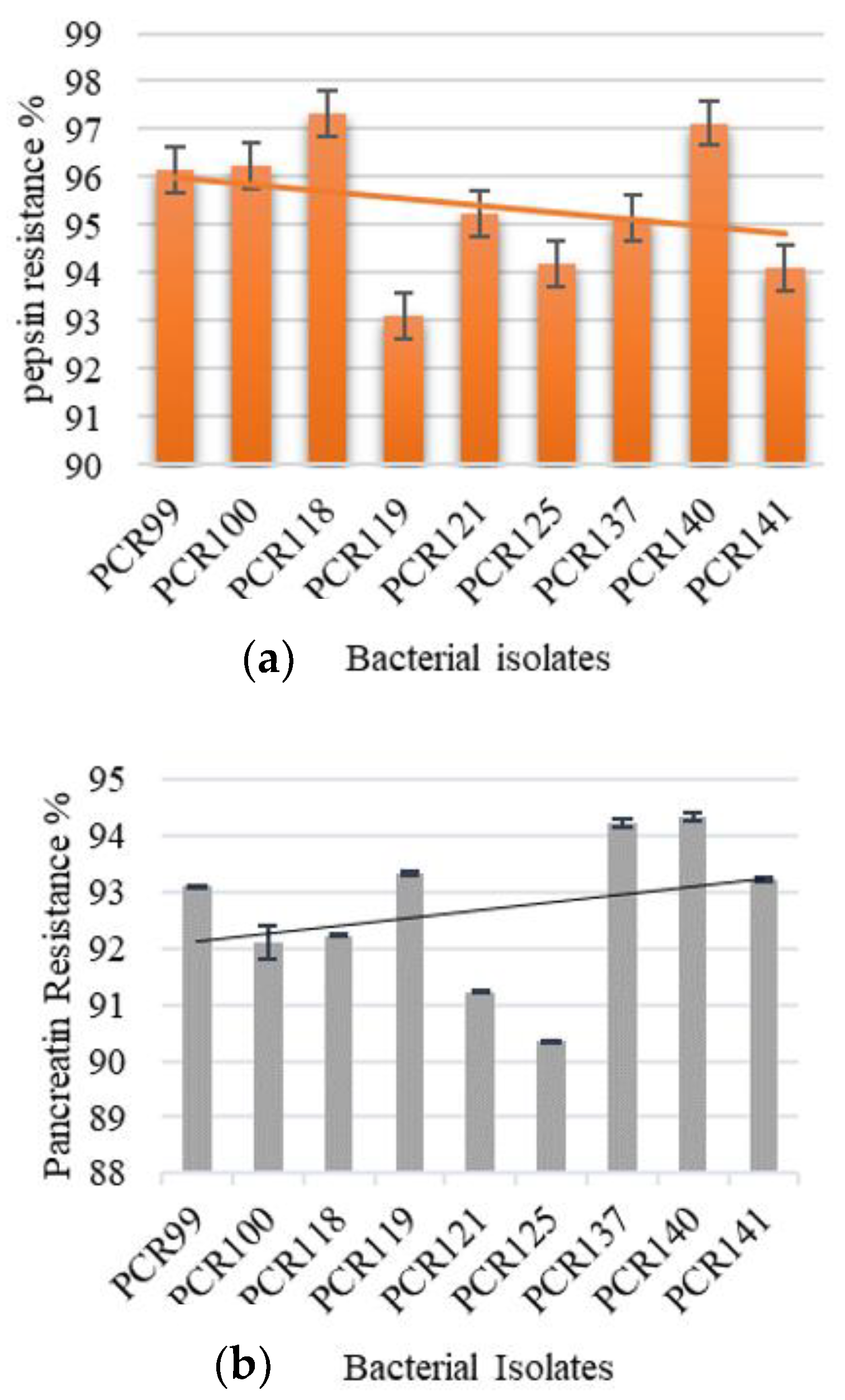
3.4.5. Pepsin and Pancreatin Resistance Assay
3.4.6. Antioxidant Activity against DPPH
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sayın, F.K. The effect of pickling on total phenolic contents and antioxidant activity of 10 vegetables. J. Food Sci. 2015, 1, 135–141. [Google Scholar] [CrossRef]
- Lee, E.; Jung, S.-R.; Lee, S.-Y.; Lee, N.-K.; Paik, H.-D.; Lim, S.-I. Lactiplantibacillus plantarum strain LN4 attenuates diet-induced obesity, insulin resistance, and changes in hepatic mrna levels associated with glucose and lipid metabolism. Nutrients 2018, 10, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaskheli, G.B.; Zuo, F.L.; Yu, R.; Chen, S.W. Overexpression of small heat shock protein enhances heat- and salt-stress tolerance of Bifidobacterium Longum NCC2705. Curr. Microbiol. 2015, 71, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Monika, S.; Kumar, V.; Kumari, A.; Angmo, K.; Bhalla, T.C. Isolation and characterization of lactic acid bacteria from traditional pickles of Himachal Pradesh, India. J. Food Sci. Technol. 2017, 54, 1945–1952. [Google Scholar] [CrossRef]
- Perczak, A.; Goliński, P.; Bryła, M.; Waśkiewicz, A. The efficiency of lactic acid bacteria against pathogenic fungi and mycotoxins. Arch. Ind. Hyg. Toxicol. 2018, 69, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Alhashem, Y.N.; Farid, A.; Al Mohaini, M.; Muzammal, M.; Khan, M.H.; Dadrasnia, A.; Alsalman, A.J.; Al Hawaj, M.A.; Ghazanfar, S.; Almusalami, E.M.; et al. Protein Isolation and Separation Techniques of Pasteurella multocidavia One-and Two-Dimensional Gel Electrophoresis. Int. J. Cur. Res. Rev. 2022, 14, 1–8. [Google Scholar] [CrossRef]
- Choi, I.H.; Noh, J.S.; Han, J.-S.; Kim, H.J.; Han, E.-S.; Song, Y.O. Kimchi, a fermented vegetable, improves serum lipid profiles in healthy young adults: Randomized clinical trial. J. Med. Food 2013, 16, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.-P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common vegetables in Singapore. Food Chem. 2010, 120, 993–1003. [Google Scholar] [CrossRef]
- Takebayashi, J.; Oki, T.; Watanabe, J.; Yamasaki, K.; Chen, J.; Sato-Furukawa, M.; Tsubota-Utsugi, M.; Taku, K.; Goto, K.; Matsumoto, T.; et al. Hydrophilic antioxidant capacities of vegetables and fruits commonly consumed in Japan and estimated average daily intake of hydrophilic antioxidants from these foods. J. Food Compos. Anal. 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Klein, M.; Sanders, M.E.; Duong, T.; Young, H.A. Probiotics: From bench to market. Ann. N. Y. Acad. Sci. 2010, 1212, E1–E14. [Google Scholar] [CrossRef]
- Zuo, F.; Yu, R.; Feng, X.; Chen, L.; Zeng, Z.; Khaskheli, G.B.; Ma, H.; Chen, S. Characterization and in vitro properties of potential probiotic bifidobacterium strains isolated from breast-fed infant feces. Ann. Microbiol. 2015, 66, 1027–1037. [Google Scholar] [CrossRef]
- Olaoye, O.A.; Ndife, J.; Raymond, V.I. Use of Lactiplantibacillus plantarum as starter culture and its influence on physicochemical, microbiological, and sensory characteristics of kunnu-aya produced from Sorghum and tigernut. J. Food Qual. 2017, 2017, 6738137. [Google Scholar] [CrossRef] [Green Version]
- Akabanda, F.; Owusu-Kwarteng, J.; Tano-Debrah, K.; Parkouda, C.; Jespersen, L. The use of lactic acid bacteria starter culture in the production of Nunu, a spontaneously fermented milk product in Ghana. Int. J. Food Sci. 2014, 2014, 721067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaseen, M.; Kamran, M.; Farid, A.; Ismail, S.; Muzammal, M.; Amir, K.A.; Rashid, S.A. Antibacterial, Hemagglutination, and Insecticidal Activity Studies on the Solvent Extracts of the Roots of Olea ferruginea. Makara J. Sci. 2022, 26, 8. [Google Scholar]
- Salomskiene, J.; Jonkuviene, D.; Macioniene, I.; Abraitiene, A.; Zeime, J.; Repeckiene, J.; Vaiciulyte-Funk, L. Differences in the occurence and efficiency of antimicrobial compounds produced by lactic acid bacteria. Eur. Food Res. Technol. 2019, 245, 569–579. [Google Scholar] [CrossRef]
- Kanwal, H.; Di Cerbo, A.; Zulfiqar, F.; Sabia, C.; Nawaz, A.; Siddiqui, F.M.; Aqeel, M.; Ghazanfar, S. Probiotic characterization and population diversity analysis of gut-associated Pediococcus acidilactici for its potential use in the dairy industry. Appl. Sci. 2021, 11, 9586. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H. New Perspectives on probiotics in health and disease. Food Sci. Hum. Wellness 2015, 4, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Mohanty, U.; Majumdar, R.K. Isolation and characterization of lactic acid bacteria from traditional fermented fish product Shidal of India with reference to their probiotic potential. LWT 2021, 146, 111641. [Google Scholar] [CrossRef]
- Ogunbanwo, S.T.; Sanni, A.I.; Onilude, A.A. Characterization of bacteriocin produced by Lactiplantibacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol. 2003, 2, 219–227. [Google Scholar]
- Mohammed, S.; Çon, A.H. Isolation and characterization of potential probiotic lactic acid bacteria from traditional cheese. LWT 2021, 152, 112319. [Google Scholar] [CrossRef]
- Prawan, K.; Bhima, B. Isolation and characterization of lactic acid bacteria for probiotic application from plant sources. Int. J. Adv. Res. 2017, 5, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Breed, R.S.; Bergey, D.H. Bergey’s Manual of Determinative Bacteriology, 7th ed.; Williams & Wilkins Co.: Baltimore, MD, USA, 1957. [Google Scholar]
- Guo, X.-H.; Kim, J.-M.; Nam, H.-M.; Park, S.-Y.; Kim, J.-M. Screening lactic acid bacteria from swine origins for multistrain probiotics based on in vitro functional properties. Anaerobe 2010, 16, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Singhal, N.; Singh, N.S.; Mohanty, S.; Kumar, M.; Virdi, J.S. Rhizospheric lactobacillus plantarum (Lactiplantibacillus plantarum) strains exhibit bile salt hydrolysis, hypocholestrolemic and probiotic capabilities in vitro. Sci. Rep. 2021, 11, 15288. [Google Scholar] [CrossRef] [PubMed]
- Kılıç, G.B.; Karahan, A.G. Identification of lactic acid bacteria isolated from the fecal samples of healthy humans and patients with dyspepsia, and determination of their ph, bile, and antibiotic tolerance properties. J. Mol. Microbiol. Biotechnol. 2010, 18, 220–229. [Google Scholar] [CrossRef]
- Yadav, R.; Puniya, A.K.; Shukla, P. Probiotic properties of lactobacillus plantarum rypr1 from an indigenous fermented beverage Raabadi. Front. Microbiol. 2016, 7, 1683. [Google Scholar] [CrossRef] [Green Version]
- Gharbi, Y.; Fhoula, I.; Ruas-Madiedo, P.; Afef, N.; Boudabous, A.; Gueimonde, M.; Ouzari, H.-I. In-vitro characterization of potentially probiotic lactobacillus strains isolated from human microbiota: Interaction with pathogenic bacteria and the enteric cell line HT29. Ann. Microbiol. 2018, 69, 61–72. [Google Scholar] [CrossRef]
- Jamaly, N. Probiotic potential of lactobacillus strains isolated from known popular traditional Moroccan dairy products. Br. Microbiol. Res. J. 2011, 1, 79–94. [Google Scholar] [CrossRef]
- Ghazanfar, S. Study on the Effect of Dietary Supplementation of Saccharomyces cerevisiae on Performance of Dairy Cattle and Heifers. Ph.D. Thesis, Department of Microbiology, Quaid-i-Azam University, Islamabad, Pakistan, 2016. [Google Scholar]
- Nadri, M.H.; Salim, Y.; Basar, N.; Yahya, A.; Zulkifli, R.M. Antioxidant activities and tyrosinase inhibition effects of phaleria macrocarpa extracts. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Al Mohaini, M.; Farid, A.; Muzammal, M.; Dadrasnia, A.; Alsalman, A.J.; Al Hawaj, M.A.; Alhashem, Y.N.; Ismail, S. Pathological study of Pasteurella Multocida Recombinant Clone ABA392. Pak. J. Med. Health Sci. 2022, 16, 1112–1116. [Google Scholar] [CrossRef]
- Adesulu-Dahunsi, A.T.; Jeyaram, K.; Sanni, A.I. Probiotic and technological properties of exopolysaccharide producing lactic acid bacteria isolated from cereal-based Nigerian fermented food products. Food Control 2018, 92, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Hattingh, M.; Alexander, A.; Meijering, I.; Van, R.C.; Dicks, L.M. Amylolytic strains of lactobacillus plantarum isolated from Barley. Afr. J. Biotechnol. 2015, 14, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Khalil, T.; Okla, M.K.; Al-Qahtani, W.H.; Ali, F.; Zahra, M.; Shakeela, Q.; Ahmed, S.; Akhtar, N.; AbdElgawad, H.; Asif, R.; et al. Tracing probiotic producing bacterial species from gut of buffalo (bubalus bubalis), south-east-Asia. Braz. J. Biol. 2022, 84, e259094. [Google Scholar] [CrossRef]
- Atanassova, M. Isolation and partial biochemical characterization of a proteinaceous anti-bacteria and anti-yeast compound produced by Lacticaseibacillus paracasei subsp. paracasei strain M3. Int. J. Food Microbiol. 2003, 87, 63–73. [Google Scholar] [CrossRef]
- Khan, A.N.; Yasmin, H.; Ghazanfar, S.; Hassan, M.N.; Keyani, R.; Khan, I.; Gohar, M.; Shahzad, A.; Hashim, M.J.; Ahmad, A. Antagonistic, anti-oxidant, anti-inflammatory and anti-diabetic probiotic potential of lactobacillus agilis isolated from the rhizosphere of the medicinal plants. Saudi J. Biol. Sci. 2021, 28, 6069–6076. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Joshi, D.; Adsul, M.; Varma, A.; Gokhale, D. D-(−)-lactic acid production from cellobiose and cellulose by lactobacillus lactis mutant RM2-24. Curr. Green Chem. 2010, 12, 1106–1109. [Google Scholar] [CrossRef]
- Dinçer, E.; Kıvanç, M. Lipolytic activity of lactic acid bacteria isolated from Turkish pastırma. Anadolu Univ. J. Sci. Technol.–C Life Sci. Biotechnol. 2018, 7, 12–19. [Google Scholar] [CrossRef]
- Farid, A.; Shah, A.H.; Ayaz, M.; Amin, A.; Yaseen, M.; Ullah, H.; Haq, F. Comparative study of biological activity of glutathione, sodium tungstate and glutathione-tungstate mixture. Afr. J. Biotechnol. 2012, 11, 10431–10437. [Google Scholar]
- Abid, R.; Ghazanfar, S.; Farid, A.; Sulaman, S.M.; Idrees, M.; Amen, R.A.; Muzammal, M.; Shahzad, M.K.; Mohamed, M.O.; Khaled, A.A.; et al. Pharmacological Properties of 4′, 5,7-Trihydroxyflavone (Apigenin) and Its Impact on Cell Signaling Pathways. Molecules 2022, 27, 4304. [Google Scholar] [CrossRef]
- Patel, M.; Siddiqui, A.J.; Hamadou, W.S.; Surti, M.; Awadelkareem, A.M.; Ashraf, S.A.; Alreshidi, M.; Snoussi, M.; Rizvi, S.M.; Bardakci, F.; et al. Inhibition of bacterial adhesion and antibiofilm activities of a glycolipid biosurfactant from lactobacillus rhamnosus with its physicochemical and functional properties. Antibiotics 2021, 10, 1546. [Google Scholar] [CrossRef]
- Abriouel, H.; Casado Muñoz, M.d.C.; Lavilla Lerma, L.; Pérez Montoro, B.; Bockelmann, W.; Pichner, R.; Kabisch, J.; Cho, G.-S.; Franz, C.M.A.P.; Gálvez, A.; et al. New insights in antibiotic resistance of lactobacillus species from fermented foods. Int. Food Res. J. 2015, 78, 465–481. [Google Scholar] [CrossRef]
- Alsalman, A.J.; Farid, A.; Al Mohaini, M.; Al Hawaj, M.A.; Muzammal, M.; Khan, M.H.; Dadrasnia, A.; Alhashem, Y.N.; Ghazanfar, S.; Almusalami, E.M.; et al. Analysis and Characterization of Chitinase in Bacillus salmalaya Strain 139SI. Int. J. Curr. Res. Rev. 2022, 14, 31–36. [Google Scholar] [CrossRef]
- Al Hawaj, M.A.; Farid, A.; Al Mohaini, M.; Alsalman, A.J.; Muzammal, M.; Khan, M.H.; Dadrasnia, A.; Alhashem, Y.N.; Ghazanfar, S.; Almusalami, E.M.; et al. Biosurfactant Screening and Antibiotic Analysis of Bacillus salmalaya. Int. J. Cur. Res. Rev. 2022, 14, 56–64. [Google Scholar] [CrossRef]
- Al Mohaini, M.; Farid, A.; Alsalman, A.J.; Al Hawaj, M.A.; Alhashem, Y.N.; Ghazanfar, S.; Muzammal, M.; Khan, M.H.; Dadrasnia, A.; Ismail, S. Screening of Anticancer and Immunomodulatory Properties of Recombinant pQE-HAS113 Clone Derived from Streptococcus Equi. Pak. J. Med. Health Sci. 2022, 16, 1100. [Google Scholar] [CrossRef]
- Angmo, K.; Kumari, A.; Savitri; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT—Food Sci. Tech. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- FAO/WHO. Probiotics in food. In Health and Nutritional Properties and Guidelines for Evaluation; FAO Food and Nutrition Paper 85; FAO: Rome, Italy, 2006. [Google Scholar]
- Saadullah, M.; Asif, M.; Farid, A.; Naseem, F.; Rashid, S.A.; Ghazanfar, S.; Muzammal, M.; Ahmad, S.; Bin Jardan, Y.A.; Alshaya, H.; et al. A Novel Distachionate from Breynia distachia Treats Inflammations by Modulating COX-2 and Inflammatory Cytokines in Rat Liver Tissue. Molecules 2022, 27, 2596. [Google Scholar] [CrossRef]
- Borah, D.; Gogoi, O.; Adhikari, C.; Kakoti, B.B. Isolation and characterization of the new indigenous Staphylococcus sp. DBOCP06 as a probiotic bacterium from traditionally fermented fish and meat products of Assam State. Egypt. J. Basic Appl. Sci. 2016, 3, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Al Mohaini, M.; Farid, A.; Muzammal, M.; Gazanffar, S.; Dadrasnia, A.; Alsalman, A.J.; Al Hawaj, M.A.; Alhashem, Y.N.; Ismail, S. Enhancing Lipase Production of Bacillus salmalay Strain 139SI Using Different Carbon Sources and Surfactants. Appl. Microbiol. 2022, 2, 237–247. [Google Scholar] [CrossRef]
- Ghalouni, E.; Hassaine, O.; Karam, N.-E. Phenotypic identification and technological characterization of lactic acid bacteria isolated from l’ben, an Algerian traditional fermented cow milk. J. Pure Appl. Microbiol. 2018, 12, 521–532. [Google Scholar] [CrossRef]
- Alsalman, A.J.; Farid, A.; Al Mohaini, M.; Muzammal, M.; Khan, M.H.; Dadrasnia, A.; Ismail, S. Chitinase Activity by Chitin Degrading Strain (Bacillus Salmalaya) in Shrimp Waste. Int. J. Curr. Res. Rev. 2022, 14, 11–17. [Google Scholar] [CrossRef]
- Tokatlı, M.; Gülgör, G.; Bağder Elmacı, S.; Arslankoz İşleyen, N.; Özçelik, F. In vitro properties of potential probiotic indigenous lactic acid bacteria originating from traditional pickles. Biomed Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Patel, M.; Hadi, S. Functional and health promoting inherent attributes ofenterococcus hiraeF2 as a novel probiotic isolated from the digestive tract of the freshwater fishcatla catla. PeerJ 2017, 5, e3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abid, S.; Farid, A.; Abid, R.; Rehman, M.U.; Alsanie, W.F.; Alhomrani, M.; Alamri, A.S.; Asdaq, S.M.B.; Hefft, D.I.; Saqib, S.; et al. Identification, Biochemical Characterization, and Safety Attributes of Locally Isolated Lactobacillus fermentum from Bubalus bubalis (Buffalo) Milk as a Probiotic. Microorganisms 2022, 10, 954. [Google Scholar] [CrossRef]
- Shivangi, S.; Devi, P.B.; Ragul, K.; Shetty, P.H. Probiotic Potential of Bacillus Strains Isolated from an Acidic Fermented Food Idli. Probiotics Antimicrob. Proteins 2020, 12, 1502–1513. [Google Scholar]
- Fatima, S.; Muzammal, M.; Khan, M.A.; Farid, A.; Kamran, M.; Qayum, J.; Qureshi, M.; Khan, M.N.; Khan, M.A. CRISPR/Cas9 endonucleases: A new era of genetic engineering. Abasyn J. life Sci. 2021, 4, 29–39. [Google Scholar] [CrossRef]
- Farid, A.; Javed, R.; Hayat, M.; Muzammal, M.; Khan, M.H.; Ismail, S.; Rashid, S.A. Screening of Strobilanthes urticifolia wall.ex kuntze for Antitermite and insecticidal activities. Abasyn J. life Sci. 2021, 4, 40–45. [Google Scholar] [CrossRef]
- Amaretti, A.; Di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Hameed, A.; Condò, C.; Tauseef, I.; Idrees, M.; Ghazanfar, S.; Farid, A.; Muzammal, M.; Al Mohaini, M.; Alsalman, A.J.; Al Hawaj, M.A.; et al. Isolation and Characterization of a Cholesterol-Lowering Bacteria from Bubalus bubalis Raw Milk. Fermentation 2022, 8, 163. [Google Scholar] [CrossRef]
- Ghazanfar, S. Understanding the Mechanism of Action of Indigenous Target Probiotic Yeast: Linking the Manipulation of Gut Microbiota and Performance in Animals. In Saccharomyces; Intech Open: London, UK, 2021. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Isolates | Amylase Activity | Lipase Activity | Cellulolytic Activity | Proteolysis Activity |
|---|---|---|---|---|
| PCR99 | −− | + | − | − |
| PCR100 | − | + | − | − |
| PCR118 | − | + | − | − |
| PCR119 | − | + | − | + |
| PCR121 | − | − | − | − |
| PCR125 | + | − | − | − |
| PCR137 | − | − | − | − |
| PCR140 | − | − | − | − |
| PCR141 | − | + | − | − |
| Antibiotics [Anti-Microbial Agents] | PCR99 | PCR100 | PCR118 | PCR119 | PCR121 | PCR125 | PCR137 | PCR140 | PCR141 |
|---|---|---|---|---|---|---|---|---|---|
| Penicillin G | S | I | R | R | R | R | R | R | I |
| Polymyxin B | R | R | R | S | S | S | S | R | I |
| Chloramphenicol | S | S | S | S | S | S | S | S | S |
| Ampicillin | I | R | R | S | S | S | S | S | R |
| Bacitracin | S | S | R | S | S | S | S | S | R |
| Kanamycin | R | R | S | S | S | S | S | S | S |
| Cephalexin | I | S | S | S | S | S | S | S | R |
| Tetracycline | I | R | S | S | S | S | S | S | S |
| Amoxycillin | S | R | R | S | S | S | S | S | S |
| Metronidazole | R | R | R | R | R | R | R | R | R |
| Vancomycin | S | R | S | S | S | S | R | S | S |
| Cefuroxime | I | R | R | S | S | S | S | S | S |
| Streptomycin | R | I | S | S | R | S | S | R | S |
| Gentamycin | S | I | S | S | S | S | S | S | I |
| Nalidixic Acid | S | R | S | S | I | S | R | S | R |
| ID | Treatments | Inhibition % |
|---|---|---|
| PCR140 | Supernatant | 67.000 ± 0.82 |
| Ascorbic acid [drug] | 71.1 ± 0.83 | |
| Control | 0 ± 0.81 |
| Strain ID | Strain Name/Genus | Length of 16s RNA | Bacterial Taxonomy | Similarity % of 16s RNA Gene Sequencing | Accession Numbers |
|---|---|---|---|---|---|
| PCR118 | Lactobacillus | 354 | Lacticaseibacillus paracasei | 99.08% | MN088781.1 |
| PCR119 | Lactobacillus | 417 | Lacticaseibacillus paracasei | 99.53% | MN088853.1 |
| PCR121 | Lactobacillus | 334 | Lacticaseibacillus paracasei | 99.30% | MN088850.1 |
| PCR125 | Lactobacillus | 338 | Lactiplantibacillus plantarum | 99.53% | MN089481.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akmal, U.; Ghori, I.; Elasbali, A.M.; Alharbi, B.; Farid, A.; Alamri, A.S.; Muzammal, M.; Asdaq, S.M.B.; Naiel, M.A.E.; Ghazanfar, S. Probiotic and Antioxidant Potential of the Lactobacillus Spp. Isolated from Artisanal Fermented Pickles. Fermentation 2022, 8, 328. https://doi.org/10.3390/fermentation8070328
Akmal U, Ghori I, Elasbali AM, Alharbi B, Farid A, Alamri AS, Muzammal M, Asdaq SMB, Naiel MAE, Ghazanfar S. Probiotic and Antioxidant Potential of the Lactobacillus Spp. Isolated from Artisanal Fermented Pickles. Fermentation. 2022; 8(7):328. https://doi.org/10.3390/fermentation8070328
Chicago/Turabian StyleAkmal, Urva, Ifra Ghori, Abdelbaset Mohamed Elasbali, Bandar Alharbi, Arshad Farid, Abdulhakeem S. Alamri, Muhammad Muzammal, Syed Mohammed Basheeruddin Asdaq, Mohammed A. E. Naiel, and Shakira Ghazanfar. 2022. "Probiotic and Antioxidant Potential of the Lactobacillus Spp. Isolated from Artisanal Fermented Pickles" Fermentation 8, no. 7: 328. https://doi.org/10.3390/fermentation8070328
APA StyleAkmal, U., Ghori, I., Elasbali, A. M., Alharbi, B., Farid, A., Alamri, A. S., Muzammal, M., Asdaq, S. M. B., Naiel, M. A. E., & Ghazanfar, S. (2022). Probiotic and Antioxidant Potential of the Lactobacillus Spp. Isolated from Artisanal Fermented Pickles. Fermentation, 8(7), 328. https://doi.org/10.3390/fermentation8070328