


Recent Advances in Marine Microalgae Production: Highlighting Human Health Products from Microalgae in View of the Coronavirus Pandemic (COVID-19)



Abstract
:1. Introduction
2. Build-Up Process
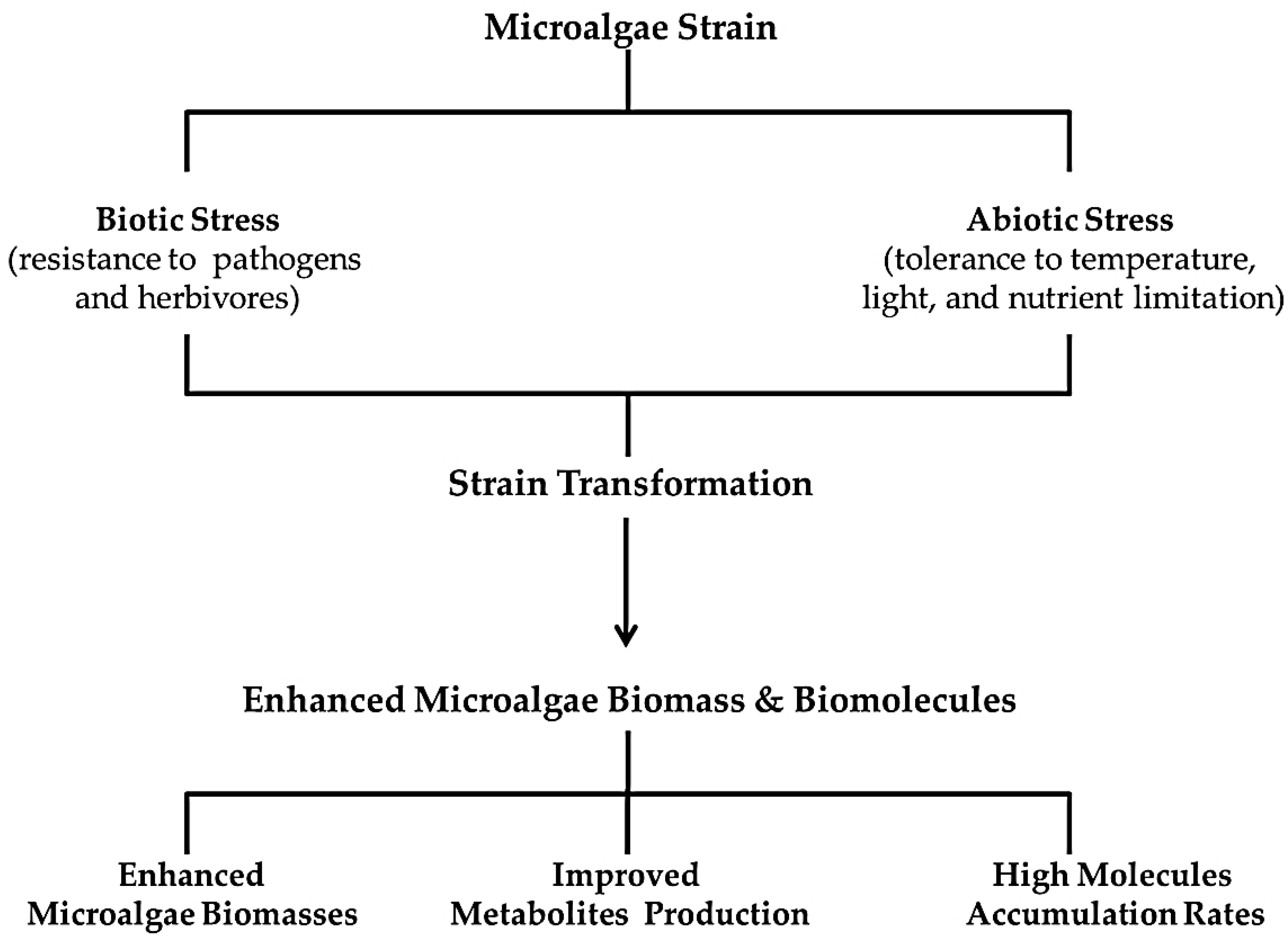
2.1. Super-Strain Concept
2.1.1. Strain Selection
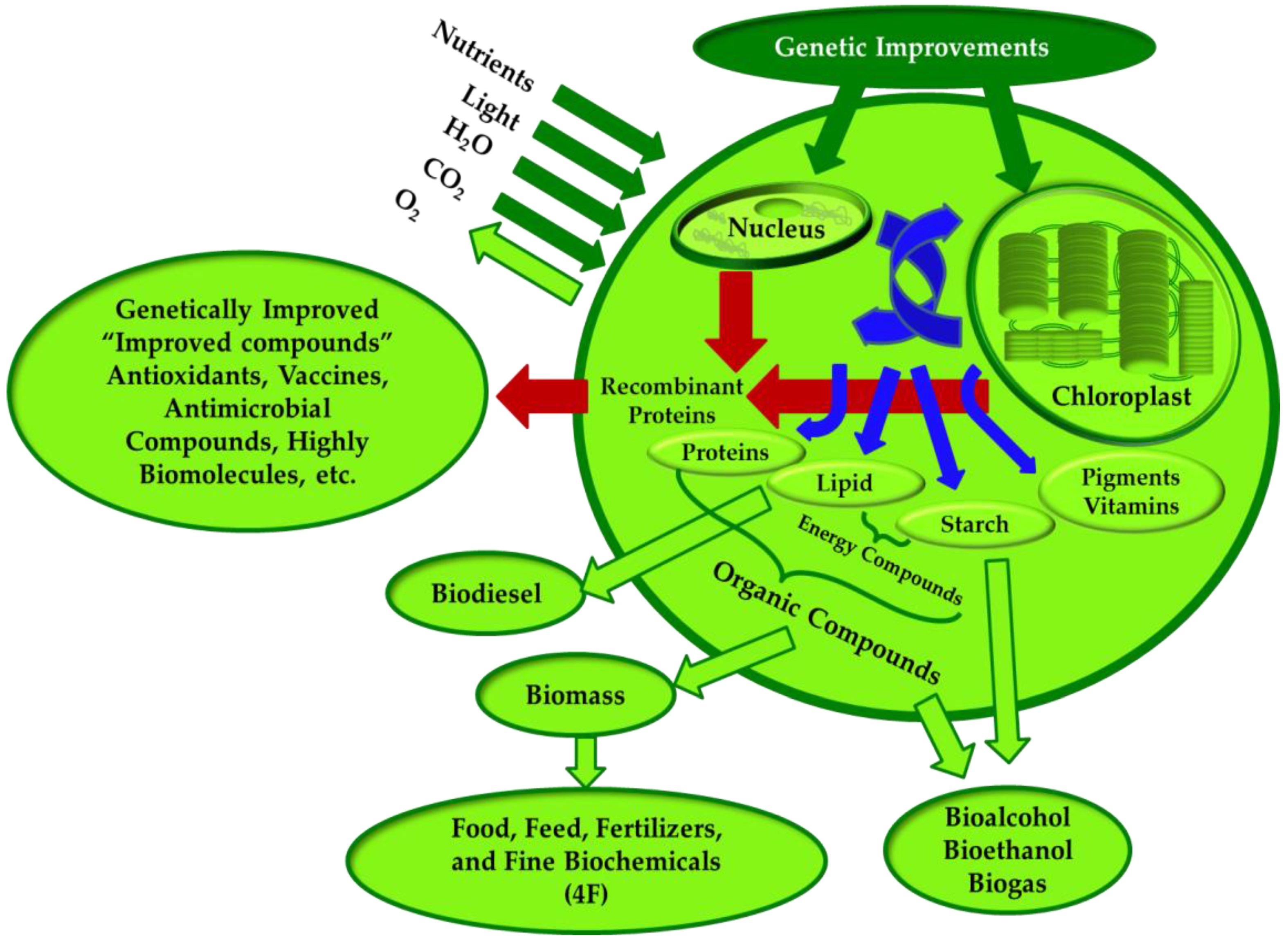
2.1.2. Strain Transformation
2.2. Cultivation Aspects
2.2.1. Cultivation Systems
2.2.2. Stress via Environmental Growth Conditions
2.2.3. Stress via Nutrient Availability and/or Limitations
2.2.4. Microalgae–Bacteria Interaction
3. Pull-Down Process
3.1. Harvesting Technologies
3.2. Drying Technologies
3.3. Extraction Technologies
4. Microalgae Derivatives against the COVID-19 Cytokine Storm
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abbas, E.M.; Ali, F.S.; Desouky, M.G.; Ashour, M.; El-Shafei, A.; Maaty, M.M.; Sharawy, Z.Z. Novel Comprehensive Molecular and Ecological Study Introducing Coastal Mud Shrimp (Solenocera Crassicornis) Recorded at the Gulf of Suez, Egypt. J. Mar. Sci. Eng. 2020, 9, 9. [Google Scholar] [CrossRef]
- Ashour, M.; Mabrouk, M.M.; Abo-Taleb, H.A.; Sharawy, Z.Z.; Ayoub, H.F.; Van Doan, H.; Davies, S.J.; El-Haroun, E.; Goda, A.M.S.A. A liquid seaweed extract (TAM®) improves aqueous rearing environment, diversity of zooplankton community, whilst enhancing growth and immune response of Nile tilapia, Oreochromis niloticus, challenged by Aeromonas hydrophila. Aquaculture 2021, 543, 736915. [Google Scholar] [CrossRef]
- El-Shenody, R.A.; Ashour, M.; Ghobara, M.M.E. Evaluating the chemical composition and antioxidant activity of three Egyptian seaweeds: Dictyota dichotoma, Turbinaria decurrens, and Laurencia obtusa. Braz. J. Food Technol. 2019, 22. [Google Scholar] [CrossRef]
- Elshobary, M.E.; El-Shenody, R.A.; Ashour, M.; Zabed, H.M.; Qi, X. Antimicrobial and antioxidant characterization of bioactive components from Chlorococcum minutum. Food Biosci. 2020, 35, 100567. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Osman, M.E.; Abo-Shady, A.M.; Komatsu, E.; Perreault, H.; Sorensen, J.; Piercey-Normore, M.D. Algal carbohydrates affect polyketide synthesis of the lichen-forming fungus Cladonia rangiferina. Mycologia 2016, 108, 646–656. [Google Scholar] [CrossRef]
- Valdovinos-García, E.M.; Petriz-Prieto, M.A.; Olán-Acosta, M.d.l.Á.; Barajas-Fernández, J.; Guzmán-López, A.; Bravo-Sánchez, M.G. Production of microalgal biomass in photobioreactors as feedstock for bioenergy and other uses: A techno-economic study of harvesting stage. Appl. Sci. 2021, 11, 4386. [Google Scholar] [CrossRef]
- Alam, M.A.; Xu, J.-L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Santhakumaran, P.; Ayyappan, S.M.; Ray, J.G. Nutraceutical applications of twenty-five species of rapid-growing green-microalgae as indicated by their antibacterial, antioxidant and mineral content. Algal Res. 2020, 47, 101878. [Google Scholar] [CrossRef]
- El-Shouny, W.; Sharaf, M.; Abomohra, A.; Abo-Eleneen, M. Production enhancement of some valuable compounds of Arthrospira Platensis. J. Basic Environ. Sci. 2015, 2, 74–83. [Google Scholar]
- Barra, L.; Chandrasekaran, R.; Corato, F.; Brunet, C. The challenge of ecophysiological biodiversity for biotechnological applications of marine microalgae. Mar. Drugs 2014, 12, 1641–1675. [Google Scholar] [CrossRef]
- Kamani, M.H.; Eş, I.; Lorenzo, J.M.; Remize, F.; Roselló-Soto, E.; Barba, F.J.; Clark, J.; Khaneghah, A.M. Advances in plant materials, food by-products, and algae conversion into biofuels: Use of environmentally friendly technologies. Green Chem. 2019, 21, 3213–3231. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Abu-Saied, M.A.; Mansour, A.T.; Ashour, M. Studying the Adsorptive Behavior of Poly(Acrylonitrile-co-Styrene) and Carbon Nanotubes (Nanocomposites) Impregnated with Adsorbent Materials towards Methyl Orange Dye. Nanomaterials 2021, 11, 1144. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, H.S.; Ibrahim, H.A.; Beltagy, E.A.; Khairy, H.M. Effects of short term feeding of some marine microalgae on the microbial profile associated with Dicentrarchus labrax post larvae. Egypt. J. Aquat. Res. 2014, 40, 251–260. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Abbas, E.; Ashry, O.; Helal, M.; Nazmi, H.; Kelany, M.; Kamel, A.; Hassaan, M.; Rossi, W.; et al. Effects of dietary marine microalgae, Tetraselmis suecica, on production, gene expression, protein markers and bacterial count of Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 2216–2228. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Labena, A.; Alsaqufi, A.; Mansour, A.; Abbas, E. Effects of dietary Arthrospira platensis nanoparticles on growth performance, feed utilization, and growth-related gene expression of Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2022, 551, 737905. [Google Scholar] [CrossRef]
- Mansour, A.T.; Ashour, M.; Abbas, E.M.; Alsaqufi, A.S.; Kelany, M.S.; El-Sawy, M.A.; Sharawy, Z.Z. Growth Performance, Immune-Related and Antioxidant Genes Expression, and Gut Bacterial Abundance of Pacific White Leg Shrimp, Litopenaeus vannamei, Dietary Supplemented With Natural Astaxanthin. Front. Physiol. 2022, 13. [Google Scholar] [CrossRef]
- Abdelrhman, A.M.; Ashour, M.; Al-Zahaby, M.A.; Sharawy, Z.Z.; Nazmi, H.; Zaki, M.A.; Ahmed, N.H.; Ahmed, S.R.; El-Haroun, E.; Van Doan, H. Effect of polysaccharides derived from brown macroalgae Sargassum dentifolium on growth performance, serum biochemical, digestive histology and enzyme activity of hybrid red tilapia. Aquac. Rep. 2022, 25, 101212. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Almutairi, A.W. A close-loop integrated approach for microalgae cultivation and efficient utilization of agar-free seaweed residues for enhanced biofuel recovery. Bioresour. Technol. 2020, 317, 124027. [Google Scholar] [CrossRef]
- Ashour, M.; Mabrouk, M.M.; Ayoub, H.F.; El-Feky, M.; Zaki, S.Z.; Hoseinifar, S.H.; Rossi, W.; Van Doan, H.; El-Haroun, E.; Goda, A.M.A.S. Effect of dietary seaweed extract supplementation on growth, feed utilization, hematological indices, and non-specific immunity of Nile Tilapia, Oreochromis niloticus challenged with Aeromonas hydrophila. J. Appl. Phycol. 2020, 32, 3467–3479. [Google Scholar] [CrossRef]
- Ashour, M.; El-Shafei, A.A.; Khairy, H.M.; Abd-Elkader, D.Y.; Mattar, M.A.; Alataway, A.; Hassan, S.M. Effect of Pterocladia capillacea Seaweed Extracts on Growth Parameters and Biochemical Constituents of Jew’s Mallow. Agronomy 2020, 10, 420. [Google Scholar] [CrossRef]
- Ashour, M.; Hassan, S.M.; Elshobary, M.E.; Ammar, G.A.G.; Gaber, A.; Alsanie, W.F.; Mansour, A.T.; El-Shenody, R. Impact of Commercial Seaweed Liquid Extract (TAM®) Biostimulant and Its Bioactive Molecules on Growth and Antioxidant Activities of Hot Pepper (Capsicum annuum). Plants 2021, 10. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Sakai, N.; Zhang, L.; Hassanien, H.A.; Gaber, A.; Ammar, G. Impact of Seaweed Liquid Extract Biostimulant on Growth, Yield, and Chemical Composition of Cucumber (Cucumis sativus). Agriculture 2021, 11, 320. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Soliman, A. Anticancer Activity, Antioxidant Activity, Mineral Contents, Vegetative and Yield of Eruca sativa Using Foliar Application of Autoclaved Cellular Extract of Spirulina platensis Extract, Comparing to NPK Fertilizers. J. Plant Prod. 2017, 8, 529–536. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Soliman, A.A.F.; Hassanien, H.A.; Alsanie, W.F.; Gaber, A.; Elshobary, M.E. The Potential of a New Commercial Seaweed Extract in Stimulating Morpho-Agronomic and Bioactive Properties of Eruca vesicaria (L.) Cav. Sustainability 2021, 13, 4485. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Elsayed, M.; Esakkimuthu, S.; El-Sheekh, M.; Hanelt, D. Potential of fat, oil and grease (FOG) for biodiesel production: A critical review on the recent progress and future perspectives. Prog. Energy Combust. Sci. 2020, 81, 100868. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Zheng, X.; Wang, Q.; Huang, J.; Ebaid, R. Enhancement of biodiesel yield and characteristics through in-situ solvo-thermal co-transesterification of wet microalgae with spent coffee grounds. Bioresour. Technol. 2020, 323, 124640. [Google Scholar] [CrossRef]
- Abomohra, A.E.F.; El-Sheekh, M.; Hanelt, D. Pilot cultivation of the chlorophyte microalga Scenedesmus obliquus as a promising feedstock for biofuel. Biomass Bioenergy 2014, 64, 237–244. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Elshobary, M. Biodiesel, bioethanol, and biobutanol production from microalgae. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Springer: Singapore, 2019; pp. 293–321. [Google Scholar]
- Grigorenko, A.; Kostyukevich, Y.I.; Chernova, N.; Kiseleva, S.; Kiseleva, E.; Popel, O.; Vladimirov, G.; Nikolaev, E.; Kumar, V.; Vlaskin, M. Hydrothermal liquefaction of Arthrospira platensis for bio-oil production and study of chemical composition for bio-oil and its gasoline fraction. Russ. J. Appl. Chem. 2019, 92, 1480–1486. [Google Scholar] [CrossRef]
- Vlaskin, M.S.; Grigorenko, A.V.; Chernova, N.I.; Kiseleva, S.V.; Lipatova, I.A.; Popel, O.S.; DombrovsKy, L.A. The hydrothermal liquefaction as a promising procedure for microalgae-to-biofuel conversion: A general review and some thermophysical problems to be solved. High Temp. High Press. 2019, 48, 309–351. [Google Scholar] [CrossRef]
- Alprol, A.E.; Ashour, M.; Mansour, A.T.; Alzahrani, O.M.; Mahmoud, S.F.; Gharib, S.M. Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. J. Mar. Sci. Eng. 2021, 9, 1328. [Google Scholar] [CrossRef]
- Alprol, A.E.; El-Metwally, M.; Amer, A. Sargassum latifolium as eco-friendly materials for treatment of toxic nickel (II) and lead (II) ions from aqueous solution. Egypt. J. Aquat. Biol. Fish. 2019, 23, 285–299. [Google Scholar] [CrossRef]
- Alprol, A.E.; Heneash, A.M.M.; Soliman, A.M.; Ashour, M.; Alsanie, W.F.; Gaber, A.; Mansour, A.T. Assessment of Water Quality, Eutrophication, and Zooplankton Community in Lake Burullus, Egypt. Diversity 2021, 13, 268. [Google Scholar] [CrossRef]
- Al-Rub, F.A.; El-Naas, M.; Ashour, I.; Al-Marzouqi, M. Biosorption of copper on Chlorella vulgaris from single, binary and ternary metal aqueous solutions. Process Biochem. 2006, 41, 457–464. [Google Scholar] [CrossRef]
- Mansour, A.T.; Alprol, A.E.; Abualnaja, K.M.; El-Beltagi, H.S.; Ramadan, K.M.A.; Ashour, M. Dried Brown Seaweed’s Phytoremediation Potential for Methylene Blue Dye Removal from Aquatic Environments. Polymers 2022, 14, 1375. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Alprol, A.E.; Abualnaja, K.M.; El-Beltagi, H.S.; Ramadan, K.M.A.; Ashour, M. The Using of Nanoparticles of Microalgae in Remediation of Toxic Dye from Industrial Wastewater: Kinetic and Isotherm Studies. Materials 2022, 15, 3922. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Alprol, A.E.; Ashour, M.; Ramadan, K.M.; Alhajji, A.H.; Abualnaja, K.M. Do Red Seaweed Nanoparticles Enhance Bioremediation Capacity of Toxic Dyes from Aqueous Solution? Gels 2022, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Alprol, A.E.; Khedawy, M.; Abualnaja, K.M.; Shalaby, T.A.; Rayan, G.; Ramadan, K.M.; Ashour, M. Green Synthesis of Zinc Oxide Nanoparticles Using Red Seaweed for the Elimination of Organic Toxic Dye from an Aqueous Solution. Materials 2022, 15, 5169. [Google Scholar] [CrossRef]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant activity of extracts from marine macroalgae, wild-collected and cultivated, in an integrated multi-trophic aquaculture system. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- Ardiles, P.; Cerezal-Mezquita, P.; Salinas-Fuentes, F.; Órdenes, D.; Renato, G.; Ruiz-Domínguez, M.C. Biochemical Composition and Phycoerythrin Extraction from Red Microalgae: A Comparative Study Using Green Extraction Technologies. Processes 2020, 8, 1628. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Aguilar-Manjarrez, J.; Cornish, L.; Dabbadie, L.; Desrochers, A.; Diffey, S.; Garrido Gamarro, E.; Geehan, J.; Hurtado, A. Seaweeds and Microalgae: AnOverview for Unlocking Their Potential in Global Aquaculture Development; FAO: Rome, Italy, 2021. [Google Scholar]
- World Fisheries and Aquaculture. 2020. Available online: https://www.aqua.cl/wp-content/uploads/2020/06/FAO-SOFIA-2020-Brief.pdf (accessed on 29 July 2022).
- Larson, S.; Stoeckl, N.; Fachry, M.E.; Mustafa, M.D.; Lapong, I.; Purnomo, A.H.; Rimmer, M.A.; Paul, N.A. Women’s well-being and household benefits from seaweed farming in Indonesia. Aquaculture 2021, 530, 735711. [Google Scholar] [CrossRef]
- Vigani, M.; Parisi, C.; Rodríguez-Cerezo, E.; Barbosa, M.J.; Sijtsma, L.; Ploeg, M.; Enzing, C. Food and feed products from micro-algae: Market opportunities and challenges for the EU. Trends Food Sci. Technol. 2015, 42, 81–92. [Google Scholar] [CrossRef]
- Callegari, A.; Bolognesi, S.; Cecconet, D.; Capodaglio, A.G. Production technologies, current role, and future prospects of biofuels feedstocks: A state-of-the-art review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 384–436. [Google Scholar] [CrossRef]
- Walsh, M.J.; Van Doren, L.G.; Sills, D.L.; Archibald, I.; Beal, C.M.; Lei, X.G.; Huntley, M.E.; Johnson, Z.; Greene, C.H. Algal food and fuel coproduction can mitigate greenhouse gas emissions while improving land and water-use efficiency. Environ. Res. Lett. 2016, 11, 114006. [Google Scholar] [CrossRef]
- Zaky, A.S. Introducing a Marine Biorefinery System for the Integrated Production of Biofuels, High-Value-Chemicals, and Co-Products: A Path Forward to a Sustainable Future. Processes 2021, 9, 1841. [Google Scholar] [CrossRef]
- Zaky, A.S.; Kumar, S.; Welfle, A.J. Integrated Approaches and Future Perspectives. In Waste-to-Energy; Springer: Berlin/Heidelberg, Germany, 2022; pp. 613–651. [Google Scholar]
- Zhu, L. Biorefinery as a promising approach to promote microalgae industry: An innovative framework. Renew. Sustain. Energy Rev. 2015, 41, 1376–1384. [Google Scholar] [CrossRef]
- Béchet, Q.; Plouviez, M.; Chambonnière, P.; Guieysse, B. Environmental impacts of full-scale algae cultivation. In Microalgae-Based Biofuels and Bioproducts; Elsevier: Amsterdam, The Netherlands, 2017; pp. 505–525. [Google Scholar]
- Mayers, J.J.; Ekman Nilsson, A.; Svensson, E.; Albers, E. Integrating microalgal production with industrial outputs—Reducing process inputs and quantifying the benefits. Ind. Biotechnol. 2016, 12, 219–234. [Google Scholar] [CrossRef]
- Usher, P.K.; Ross, A.B.; Camargo-Valero, M.A.; Tomlin, A.S.; Gale, W.F. An overview of the potential environmental impacts of large-scale microalgae cultivation. Biofuels 2014, 5, 331–349. [Google Scholar] [CrossRef]
- Mansour, A.T.; Ashour, M.; Alprol, A.E.; Alsaqufi, A.S. Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution. Sustainability 2022, 14, 3257. [Google Scholar] [CrossRef]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M. Current status of the algae production industry in Europe: An emerging sector of the Blue Bioeconomy. Front. Mar. Sci. 2021, 7, 1247. [Google Scholar] [CrossRef]
- Muhammad, G.; Alam, M.; Xiong, W.; Lv, Y.; Xu, J.-L. Microalgae biomass production: An overview of dynamic operational methods. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Singapore, 2020; pp. 415–432. [Google Scholar]
- Talukdar, J.; Dasgupta, S.; Nagle, V.; Bhadra, B. COVID-19: Potential of microalgae derived natural astaxanthin as adjunctive supplement in alleviating cytokine storm. SSRN. 2020. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3579738 (accessed on 29 July 2022).
- Alam, M.A.; Parra-Saldivar, R.; Bilal, M.; Afroze, C.A.; Ahmed, M.N.; Iqbal, H.M.; Xu, J. Algae-derived bioactive molecules for the potential treatment of SARS-CoV-2. Molecules 2021, 26, 2134. [Google Scholar] [CrossRef]
- Ziyaei, K.; Ataie, Z.; Mokhtari, M.; Adrah, K.; Daneshmehr, M.A. An insight to the therapeutic potential of algae-derived sulfated polysaccharides and polyunsaturated fatty acids: Focusing on the COVID-19. Int. J. Biol. Macromol. 2022, 209, 244–257. [Google Scholar] [CrossRef]
- Tzachor, A.; Rozen, O.; Khatib, S.; Jensen, S.; Avni, D. Photosynthetically controlled spirulina, but not solar spirulina, inhibits TNF-α secretion: Potential implications for COVID-19-related cytokine storm therapy. Mar. Biotechnol. 2021, 23, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Rajvanshi, M.; Sayre, R. Recent advances in algal biomass production. In Biotechnological Applications of Biomass; Intech Open: London, UK, 2020. [Google Scholar]
- Guiry, M.; Guiry, G. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2013. [Google Scholar]
- Yew, G.Y.; Lee, S.Y.; Show, P.L.; Tao, Y.; Law, C.L.; Nguyen, T.T.C.; Chang, J.-S. Recent advances in algae biodiesel production: From upstream cultivation to downstream processing. Bioresour. Technol. Rep. 2019, 7, 100227. [Google Scholar] [CrossRef]
- Neofotis, P.; Huang, A.; Sury, K.; Chang, W.; Joseph, F.; Gabr, A.; Twary, S.; Qiu, W.; Holguin, O.; Polle, J.E. Characterization and classification of highly productive microalgae strains discovered for biofuel and bioproduct generation. Algal Res. 2016, 15, 164–178. [Google Scholar] [CrossRef]
- Saini, D.K.; Chakdar, H.; Pabbi, S.; Shukla, P. Enhancing production of microalgal biopigments through metabolic and genetic engineering. Crit. Rev. Food Sci. Nutr. 2020, 60, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, M.; Chew, K.W.; Show, P.L.; Ling, T.C.; Ng, I.-S.; Chang, J.-S. Genetic engineering of microalgae for enhanced biorefinery capabilities. Biotechnol. Adv. 2020, 43, 107554. [Google Scholar] [CrossRef]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Factories 2018, 17, 173. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Lee, P.A.; Shen, Z.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust expression and secretion of Xylanase1 in Chlamydomonas reinhardtii by fusion to a selection gene and processing with the FMDV 2A peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Hannon, M.J.; Marcuschi, M.; Wu, S.; Botsch, K.; Lewis, A.J.; Hyun, J.; Mendez, M.; Mayfield, S.P. Production of recombinant enzymes in the marine alga Dunaliella tertiolecta. Algal Res. 2013, 2, 2–9. [Google Scholar] [CrossRef]
- Hou, Q.; Qiu, S.; Liu, Q.; Tian, J.; Hu, Z.; Ni, J. Selenoprotein-transgenic Chlamydomonas reinhardtii. Nutrients 2013, 5, 624–636. [Google Scholar] [CrossRef]
- Manuell, A.L.; Beligni, M.V.; Elder, J.H.; Siefker, D.T.; Tran, M.; Weber, A.; McDonald, T.L.; Mayfield, S.P. Robust expression of a bioactive mammalian protein in Chlamydomonas chloroplast. Plant Biotechnol. J. 2007, 5, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Muto, M.; Lee, P.A.; Jager, M.; Cardoso, R.M.; Behnke, C.A.; Kirk, P.; Hokanson, C.A.; Crea, R.; Mendez, M. Production of therapeutic proteins in algae, analysis of expression of seven human proteins in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2010, 8, 719–733. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Feng, W.; Zhao, L.; Gu, H.; Li, Q.; Shi, K.; Guo, S.; Zhang, N. Preparation of transgenic Dunaliella salina for immunization against white spot syndrome virus in crayfish. Arch. Virol. 2014, 159, 519–525. [Google Scholar] [CrossRef]
- Cha, T.-S.; Chen, C.-F.; Yee, W.; Aziz, A.; Loh, S.-H. Cinnamic acid, coumarin and vanillin: Alternative phenolic compounds for efficient Agrobacterium-mediated transformation of the unicellular green alga, Nannochloropsis sp. J. Microbiol. Methods 2011, 84, 430–434. [Google Scholar] [CrossRef]
- He, Y.; Peng, H.; Liu, J.; Chen, F.; Zhou, Y.; Ma, X.; Chen, H.; Wang, K. Chlorella sp. transgenic with Scy-hepc enhancing the survival of Sparus macrocephalus and hybrid grouper challenged with Aeromonas hydrophila. Fish Shellfish Immunol. 2018, 73, 22–29. [Google Scholar] [CrossRef]
- Lau, C.C.; Loh, S.H.; Aziz, A.; San Cha, T. Effects of disrupted omega-3 desaturase gene construct on fatty acid composition and expression of four fatty acid biosynthetic genes in transgenic Chlorella vulgaris. Algal Res. 2017, 26, 143–152. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Gerken, H.; Huang, J.; Jiang, Y.; Chen, F. Genetic engineering of the green alga Chlorella zofingiensis: A modified norflurazon-resistant phytoene desaturase gene as a dominant selectable marker. Appl. Microbiol. Biotechnol. 2014, 98, 5069–5079. [Google Scholar] [CrossRef]
- Ma, X.; Pan, K.; Zhang, L.; Zhu, B.; Yang, G.; Zhang, X. Genetic transformation of Nannochloropsis oculata with a bacterial phleomycin resistance gene as dominant selective marker. J. Ocean. Univ. China 2016, 15, 351–356. [Google Scholar] [CrossRef]
- Prasad, B.; Vadakedath, N.; Jeong, H.-J.; General, T.; Cho, M.-G.; Lein, W. Agrobacterium tumefaciens-mediated genetic transformation of haptophytes (Isochrysis species). Appl. Microbiol. Biotechnol. 2014, 98, 8629–8639. [Google Scholar] [CrossRef]
- Prasad, B. Agrobacterium-mediated Nuclear Transformation of Haptophyte and Rhodophyte Species. Ph.D. Thesis, University of Erlangen-Nuremberg, Erlangen, Germany, 2017. [Google Scholar]
- Kim, D.-H.; Kim, Y.T.; Cho, J.J.; Bae, J.-H.; Hur, S.-B.; Hwang, I.; Choi, T.-J. Stable integration and functional expression of flounder growth hormone gene in transformed microalga, Chlorella ellipsoidea. Mar. Biotechnol. 2002, 4, 63–73. [Google Scholar] [CrossRef]
- Liu, J.; Gerken, H.; Huang, J.; Chen, F. Engineering of an endogenous phytoene desaturase gene as a dominant selectable marker for Chlamydomonas reinhardtii transformation and enhanced biosynthesis of carotenoids. Process Biochem. 2013, 48, 788–795. [Google Scholar] [CrossRef]
- Lin, W.-R.; Lai, Y.-C.; Sung, P.-K.; Tan, S.-I.; Chang, C.-H.; Chen, C.-Y.; Chang, J.-S.; Ng, I.-S. Enhancing carbon capture and lipid accumulation by genetic carbonic anhydrase in microalgae. J. Taiwan Inst. Chem. Eng. 2018, 93, 131–141. [Google Scholar] [CrossRef]
- Wei, L.; Wang, Q.; Xin, Y.; Lu, Y.; Xu, J. Enhancing photosynthetic biomass productivity of industrial oleaginous microalgae by overexpression of RuBisCO activase. Algal Res. 2017, 27, 366–375. [Google Scholar] [CrossRef]
- Endo, H.; Yoshida, M.; Uji, T.; Saga, N.; Inoue, K.; Nagasawa, H. Stable nuclear transformation system for the coccolithophorid alga Pleurochrysis carterae. Sci. Rep. 2016, 6, 22252. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.-J.; Chen, H.-X.; Xu, W.-Q.; Xu, Y.-W. Expression of soybean Kunitz trypsin inhibitor gene SKTI in Dunaliella salina. J. Appl. Phycol. 2013, 25, 139–144. [Google Scholar] [CrossRef]
- Yang, Y.; Chai, Z.; Wang, Q.; Chen, W.; He, Z.; Jiang, S. Cultivation of seaweed Gracilaria in Chinese coastal waters and its contribution to environmental improvements. Algal Res. 2015, 9, 236–244. [Google Scholar] [CrossRef]
- Te, M.R.; Miller, D.J. Genetic transformation of dinoflagellates (Amphidinium and Symbiodinium): Expression of GUS in microalgae using heterologous promoter constructs. Plant, J. 1998, 13, 427–435. [Google Scholar] [CrossRef]
- Dunahay, T. Transformation of Chlamydomonas reinhardtii with silicon carbide whiskers. Biotechniques 1993, 15, 452–455, 457. [Google Scholar] [PubMed]
- Dong, B.; Cheng, R.-Q.; Liu, Q.-Y.; Wang, J.; Fan, Z.-C. Multimer of the antimicrobial peptide Mytichitin-A expressed in Chlamydomonas reinhardtii exerts a broader antibacterial spectrum and increased potency. J. Biosci. Bioeng. 2018, 125, 175–179. [Google Scholar] [CrossRef]
- Sizova, I.; Greiner, A.; Awasthi, M.; Kateriya, S.; Hegemann, P. Nuclear gene targeting in Chlamydomonas using engineered zinc-finger nucleases. Plant J. 2013, 73, 873–882. [Google Scholar] [CrossRef]
- Cui, Y.; Jiang, P.; Wang, J.; Li, F.; Chen, Y.; Zheng, G.; Qin, S. Genetic transformation of Platymonas (Tetraselmis) subcordiformis (Prasinophyceae, Chlorophyta) using particle bombardment and glass-bead agitation. Chin. J. Oceanol. Limnol. 2012, 30, 471–475. [Google Scholar] [CrossRef]
- Kathiresan, S.; Chandrashekar, A.; Ravishankar, G.; Sarada, R. Regulation of astaxanthin and its intermediates through cloning and genetic transformation of β-carotene ketolase in Haematococcus pluvialis. J. Biotechnol. 2015, 196, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Hempel, F.; Lau, J.; Klingl, A.; Maier, U.G. Algae as protein factories: Expression of a human antibody and the respective antigen in the diatom Phaeodactylum tricornutum. PLoS ONE 2011, 6, e28424. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Tamoi, M.; Shigeoka, S. Overexpression of a cyanobacterial fructose-1, 6-/sedoheptulose-1, 7-bisphosphatase in tobacco enhances photosynthesis and growth. Nat. Biotechnol. 2001, 19, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, K.K.; Banerjee, I.; Singh, D.; Sajwan, P.; Chhetri, V. Ecological stress stimulus to improve microalgae biofuel generation: A review. Octa. J. Biosci. 2020, 8, 48–54. [Google Scholar]
- Madkour, F.F.; Kamil, A.E.-W.; Nasr, H.S. Production and nutritive value of Spirulina platensis in reduced cost media. Egypt. J. Aquat. Res. 2012, 38, 51–57. [Google Scholar] [CrossRef]
- González-González, L.M.; de-Bashan, L.E. Toward the enhancement of microalgal metabolite production through microalgae–bacteria consortia. Biology 2021, 10, 282. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- de Carvalho, J.C.; Sydney, E.B.; Tessari, L.F.A.; Soccol, C.R. Culture media for mass production of microalgae. In Biofuels from Algae; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–50. [Google Scholar]
- De Godos, I.; Mendoza, J.; Acién, F.; Molina, E.; Banks, C.; Heaven, S.; Rogalla, F. Evaluation of carbon dioxide mass transfer in raceway reactors for microalgae culture using flue gases. Bioresour. Technol. 2014, 153, 307–314. [Google Scholar] [CrossRef]
- Zerrouki, D.; Henni, A. Outdoor Microalgae Cultivation for Wastewater Treatment. In Application of Microalgae in Wastewater Treatment; Springer: Berlin/Heidelberg, Germany, 2019; pp. 81–99. [Google Scholar]
- Kumar, K.; Mishra, S.K.; Shrivastav, A.; Park, M.S.; Yang, J.-W. Recent trends in the mass cultivation of algae in raceway ponds. Renew. Sustain. Energy Rev. 2015, 51, 875–885. [Google Scholar] [CrossRef]
- Al-lwayzy, S.H.; Yusaf, T.; Al-Juboori, R.A. Biofuels from the fresh water microalgae Chlorella vulgaris (FWM-CV) for diesel engines. Energies 2014, 7, 1829–1851. [Google Scholar] [CrossRef]
- Rahul, M.; Sundaramahalingam, M.; Shivamthi, C.; Shyam, K.; Varalkshmi, P.; Karthikumar, S.; Kanimozhi, J.; Vinoth, K.; Sabarathinam, S.; Ganesh, M. Insights about sustainable biodiesel production from microalgae biomass: A review. Int. J. Energy Res. 2021, 45, 17028–17056. [Google Scholar] [CrossRef]
- Vickers, N.J. Animal communication: When i’m calling you, will you answer too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.; Cañavate, J.P.; García-González, M. Assessment of carotenoid production by Dunaliella salina in different culture systems and operation regimes. J. Biotechnol. 2011, 151, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Bošnjaković, M.; Sinaga, N. The perspective of large-scale production of algae biodiesel. Appl. Sci. 2020, 10, 8181. [Google Scholar] [CrossRef]
- Li, J.; Zhu, D.; Niu, J.; Shen, S.; Wang, G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef]
- Chandra, R.; Iqbal, H.M.; Vishal, G.; Lee, H.-S.; Nagra, S. Algal biorefinery: A sustainable approach to valorize algal-based biomass towards multiple product recovery. Bioresour. Technol. 2019, 278, 346–359. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Oslan, S.N.; Mohamad, R.; Tan, J.S.; Yusoff, A.H.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction. Fermentation 2022, 8, 179. [Google Scholar] [CrossRef]
- Barati, B.; Gan, S.-Y.; Lim, P.-E.; Beardall, J.; Phang, S.-M. Green algal molecular responses to temperature stress. Acta Physiol. Plant. 2019, 41, 26. [Google Scholar] [CrossRef]
- Ras, M.; Steyer, J.-P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Bio Technol. 2013, 12, 153–164. [Google Scholar] [CrossRef]
- Covarrubias, Y.; Cantoral-Uriza, E.A.; Casas-Flores, J.S.; García-Meza, J.V. Thermophile mats of microalgae growing on the woody structure of a cooling tower of a thermoelectric power plant in Central Mexico. Rev. Mex. Biodivers. 2016, 87, 277–287. [Google Scholar] [CrossRef]
- Ferro, L.; Gorzsás, A.; Gentili, F.G.; Funk, C. Subarctic microalgal strains treat wastewater and produce biomass at low temperature and short photoperiod. Algal Res. 2018, 35, 160–167. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Ho, S.-H.; Ye, X.; Hasunuma, T.; Chang, J.-S.; Kondo, A. Perspectives on engineering strategies for improving biofuel production from microalgae—A critical review. Biotechnol. Adv. 2014, 32, 1448–1459. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.F.; Menegol, T.; Rech, R. Biochemical composition of green microalgae Pseudoneochloris marina grown under different temperature and light conditions. Biocatal. Agric. Biotechnol. 2019, 18, 101032. [Google Scholar] [CrossRef]
- El-Khouly, M.E.; El-Mohsnawy, E.; Fukuzumi, S. Solar energy conversion: From natural to artificial photosynthesis. J. Photochem. Photobiol. C Photochem. Rev. 2017, 31, 36–83. [Google Scholar] [CrossRef]
- Amini Khoeyi, Z.; Seyfabadi, J.; Ramezanpour, Z. Effect of light intensity and photoperiod on biomass and fatty acid composition of the microalgae, Chlorella vulgaris. Aquac. Int. 2012, 20, 41–49. [Google Scholar] [CrossRef]
- Sforza, E.; Barbera, E.; Bertucco, A. Improving the photoconversion efficiency: An integrated photovoltaic-photobioreactor system for microalgal cultivation. Algal Res. 2015, 10, 202–209. [Google Scholar] [CrossRef]
- Mohsenpour, S.F.; Willoughby, N. Luminescent photobioreactor design for improved algal growth and photosynthetic pigment production through spectral conversion of light. Bioresour. Technol. 2013, 142, 147–153. [Google Scholar] [CrossRef]
- Das, P.; Lei, W.; Aziz, S.S.; Obbard, J.P. Enhanced algae growth in both phototrophic and mixotrophic culture under blue light. Bioresour. Technol. 2011, 102, 3883–3887. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Solórzano, G.G.; Campos, J.E.; Martínez-García, M.; Morales, M. Characterization of Scenedesmus obtusiusculus AT-UAM for high-energy molecules accumulation: Deeper insight into biotechnological potential of strains of the same species. Biotechnol. Rep. 2018, 17, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Gifuni, I.; Olivieri, G.; Pollio, A.; Marzocchella, A. Identification of an industrial microalgal strain for starch production in biorefinery context: The effect of nitrogen and carbon concentration on starch accumulation. New Biotechnol. 2018, 41, 46–54. [Google Scholar] [CrossRef] [PubMed]
- de Farias Silva, C.E.; Sforza, E.; Bertucco, A. Stability of carbohydrate production in continuous microalgal cultivation under nitrogen limitation: Effect of irradiation regime and intensity on Tetradesmus obliquus. J. Appl. Phycol. 2018, 30, 261–270. [Google Scholar] [CrossRef]
- Coronel, C.D.; Do Nascimento, M.; Curatti, L. Effect of matching microalgal strains origin and regional weather condition on biomass productivity in environmental photobioreactors. Bioresour. Technol. Rep. 2019, 5, 104–112. [Google Scholar] [CrossRef]
- García-Cubero, R.; Cabanelas, I.T.D.; Sijtsma, L.; Kleinegris, D.M.; Barbosa, M.J. Production of exopolysaccharide by Botryococcus braunii CCALA 778 under laboratory simulated Mediterranean climate conditions. Algal Res. 2018, 29, 330–336. [Google Scholar] [CrossRef]
- de Farias Silva, C.E.; Barbera, E.; Bertucco, A. Biorefinery as a promising approach to promote ethanol industry from microalgae and cyanobacteria. In Bioethanol Production from Food Crops; Elsevier: Amsterdam, The Netherlands, 2019; pp. 343–359. [Google Scholar]
- Samiee-Zafarghandi, R.; Karimi-Sabet, J.; Abdoli, M.A.; Karbassi, A. Increasing microalgal carbohydrate content for hydrothermal gasification purposes. Renew. Energy 2018, 116, 710–719. [Google Scholar] [CrossRef]
- Selvakumar, P.; Umadevi, K. Mass cultivation of marine micro alga Nannochloropsis gaditana KF410818 isolated from Visakhapatnam offshore and fatty acid profile analysis for biodiesel production. J. Algal Biomass Util. 2014, 5, 28–37. [Google Scholar]
- Wu, L.F.; Chen, P.C.; Lee, C.M. The effects of nitrogen sources and temperature on cell growth and lipid accumulation of microalgae. Int. Biodeterior. Biodegrad. 2013, 85, 506–510. [Google Scholar] [CrossRef]
- Ashour, M.; Kamel, A. Enhance growth and biochemical composition of Nannochloropsis oceanica, cultured under nutrient limitation, using commercial agricultural fertilizers. J. Mar. Sci. Res. Dev 2017, 7, 233. [Google Scholar]
- Lai, Y.H.; Puspanadan, S.; Lee, C.K. Nutritional optimization of Arthrospira platensis for starch and total carbohydrates production. Biotechnol. Prog. 2019, 35, e2798. [Google Scholar] [CrossRef]
- Hanifzadeh, M.; Garcia, E.C.; Viamajala, S. Production of lipid and carbohydrate from microalgae without compromising biomass productivities: Role of Ca and Mg. Renew. Energy 2018, 127, 989–997. [Google Scholar] [CrossRef]
- Mathiot, C.; Ponge, P.; Gallard, B.; Sassi, J.-F.; Delrue, F.; Le Moigne, N. Microalgae starch-based bioplastics: Screening of ten strains and plasticization of unfractionated microalgae by extrusion. Carbohydr. Polym. 2019, 208, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Dammak, M.; Hadrich, B.; Miladi, R.; Barkallah, M.; Hentati, F.; Hachicha, R.; Laroche, C.; Michaud, P.; Fendri, I.; Abdelkafi, S. Effects of nutritional conditions on growth and biochemical composition of Tetraselmis sp. Lipids Health Dis. 2017, 16, 41. [Google Scholar] [CrossRef] [PubMed]
- Hernández-García, A.; Velásquez-Orta, S.B.; Novelo, E.; Yáñez-Noguez, I.; Monje-Ramírez, I.; Ledesma, M.T.O. Wastewater-leachate treatment by microalgae: Biomass, carbohydrate and lipid production. Ecotoxicol. Environ. Saf. 2019, 174, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Hong, M.-E.; Chang, W.S.; Sim, S.J. Autotrophic biodiesel production from the thermotolerant microalga Chlorella sorokiniana by enhancing the carbon availability with temperature adjustment. Biotechnol. Bioprocess Eng. 2019, 24, 223–231. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Bicarbonate supplementation enhanced biofuel production potential as well as nutritional stress mitigation in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 193, 315–323. [Google Scholar] [CrossRef]
- Varshney, P.; Beardall, J.; Bhattacharya, S.; Wangikar, P.P. Isolation and biochemical characterisation of two thermophilic green algal species-Asterarcys quadricellulare and Chlorella sorokiniana, which are tolerant to high levels of carbon dioxide and nitric oxide. Algal Res. 2018, 30, 28–37. [Google Scholar] [CrossRef]
- Sengupta, S.; Nawaz, T.; Beaudry, J. Nitrogen and phosphorus recovery from wastewater. Curr. Pollut. Rep. 2015, 1, 155–166. [Google Scholar] [CrossRef]
- Walsh, M.J.; Van Doren, L.G.; Shete, N.; Prakash, A.; Salim, U. Financial tradeoffs of energy and food uses of algal biomass under stochastic conditions. Appl. Energy 2018, 210, 591–603. [Google Scholar] [CrossRef]
- Lerat, Y.; Cornish, M.; Critchley, A.T. Applications of algal biomass in global food and feed markets: From traditional usage to the potential for functional products. Blue Biotechnol. Prod. Use Mar. Mol. 2018, 1, 143–189. [Google Scholar]
- Hawrot-Paw, M.; Koniuszy, A.; Gałczyńska, M.; Zając, G.; Szyszlak-Bargłowicz, J. Production of microalgal biomass using aquaculture wastewater as growth medium. Water 2019, 12, 106. [Google Scholar] [CrossRef]
- Sfez, S.; Van Den Hende, S.; Taelman, S.E.; De Meester, S.; Dewulf, J. Environmental sustainability assessment of a microalgae raceway pond treating aquaculture wastewater: From up-scaling to system integration. Bioresour. Technol. 2015, 190, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Boopathy, R.; Fontenot, Q.; Kilgen, M.B. Biological treatment of sludge from a recirculating aquaculture system using a sequencing batch reactor. J. World Aquac. Soc. 2005, 36, 542–545. [Google Scholar] [CrossRef]
- Boopathy, R.; Bonvillain, C.; Fontenot, Q.; Kilgen, M. Biological treatment of low-salinity shrimp aquaculture wastewater using sequencing batch reactor. Int. Biodeterior. Biodegrad. 2007, 59, 16–19. [Google Scholar] [CrossRef]
- Schulz, C.; Gelbrecht, J.; Rennert, B. Constructed wetlands with free water surface for treatment of aquaculture effluents. J. Appl. Ichthyol. 2004, 20, 64–70. [Google Scholar] [CrossRef]
- Mirzoyan, N.; Parnes, S.; Singer, A.; Tal, Y.; Sowers, K.; Gross, A. Quality of brackish aquaculture sludge and its suitability for anaerobic digestion and methane production in an upflow anaerobic sludge blanket (UASB) reactor. Aquaculture 2008, 279, 35–41. [Google Scholar] [CrossRef]
- Suhr, K.I.; Pedersen, L.-F.; Nielsen, J.L. End-of-pipe single-sludge denitrification in pilot-scale recirculating aquaculture systems. Aquac. Eng. 2014, 62, 28–35. [Google Scholar] [CrossRef]
- Kumar, V.; Gururani, P.; Parveen, A.; Verma, M.; Kim, H.; Vlaskin, M.; Grigorenko, A.V.; Rindin, K.G. Dairy Industry wastewater and stormwater energy valorization: Effect of wastewater nutrients on microalgae-yeast biomass. Biomass Convers. Biorefinery 2022, 1–10. [Google Scholar] [CrossRef]
- Yogev, U.; Sowers, K.R.; Mozes, N.; Gross, A. Nitrogen and carbon balance in a novel near-zero water exchange saline recirculating aquaculture system. Aquaculture 2017, 467, 118–126. [Google Scholar] [CrossRef]
- Barbu, M.; Ceangă, E.; Caraman, S. Water quality modeling and control in recirculating aquaculture systems. Urban Agric. 2018, 2, 64. [Google Scholar]
- Pittman, J.K.; Dean, A.P.; Osundeko, O. The potential of sustainable algal biofuel production using wastewater resources. Bioresour. Technol. 2011, 102, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Rawat, I.; Kumar, R.R.; Mutanda, T.; Bux, F. Dual role of microalgae: Phycoremediation of domestic wastewater and biomass production for sustainable biofuels production. Appl. Energy 2011, 88, 3411–3424. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Lay, C.-H.; Chen, C.-C.; Wu, S.-Y. Lipid accumulating microalgae cultivation in textile wastewater: Environmental parameters optimization. J. Taiwan Inst. Chem. Eng. 2017, 79, 1–6. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; Mohamed, L.A. Bioremediation of the textile waste effluent by Chlorella vulgaris. Egypt. J. Aquat. Res. 2014, 40, 301–308. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Ramamurthy, D. Dyeing industry effluent system as lipid production medium of Neochloris sp. for biodiesel feedstock preparation. BioMed Res. Int. 2014, 2014, 529560. [Google Scholar] [CrossRef]
- Parikh, A.; Madamwar, D. Textile dye decolorization using cyanobacteria. Biotechnol. Lett. 2005, 27, 323–326. [Google Scholar] [CrossRef]
- Verma, K.; Kumar, P.K.; Krishna, S.V.; Himabindu, V. Phycoremediation of Sewage-Contaminated Lake Water Using Microalgae–Bacteria Co-Culture. Water Air Soil Pollut. 2020, 231, 299. [Google Scholar] [CrossRef]
- Luo, H.; Moran, M.A. Evolutionary ecology of the marine Roseobacter clade. Microbiol. Mol. Biol. Rev. 2014, 78, 573–587. [Google Scholar] [CrossRef]
- Paddock, M.B.; Fernández-Bayo, J.D.; VanderGheynst, J.S. The effect of the microalgae-bacteria microbiome on wastewater treatment and biomass production. Appl. Microbiol. Biotechnol. 2020, 104, 893–905. [Google Scholar] [CrossRef]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef]
- Peng, H.; de-Bashan, L.E.; Bashan, Y.; Higgins, B.T. Indole-3-acetic acid from Azosprillum brasilense promotes growth in green algae at the expense of energy storage products. Algal Res. 2020, 47, 101845. [Google Scholar] [CrossRef]
- Sandhya, S.; Vijayan, K. Symbiotic association among marine microalgae and bacterial flora: A study with special reference to commercially important Isochrysis galbana culture. J. Appl. Phycol. 2019, 31, 2259–2266. [Google Scholar] [CrossRef]
- Xue, L.; Shang, H.; Ma, P.; Wang, X.; He, X.; Niu, J.; Wu, J. Analysis of growth and lipid production characteristics of Chlorella vulgaris in artificially constructed consortia with symbiotic bacteria. J. Basic Microbiol. 2018, 58, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Park, B.S.; Wang, P.; Patidar, S.K.; Kim, J.H.; Kim, S.-H.; Han, M.-S. Phycospheric native bacteria Pelagibaca bermudensis and Stappia sp. ameliorate biomass productivity of Tetraselmis striata (KCTC1432BP) in co-cultivation system through mutualistic interaction. Front. Plant Sci. 2017, 8, 289. [Google Scholar] [CrossRef]
- Higgins, B.T.; VanderGheynst, J.S. Effects of Escherichia coli on mixotrophic growth of Chlorella minutissima and production of biofuel precursors. PLoS ONE 2014, 9, e96807. [Google Scholar] [CrossRef]
- Paul, C.; Mausz, M.A.; Pohnert, G. A co-culturing/metabolomics approach to investigate chemically mediated interactions of planktonic organisms reveals influence of bacteria on diatom metabolism. Metabolomics 2013, 9, 349–359. [Google Scholar] [CrossRef]
- Do Nascimento, M.; de los Angeles Dublan, M.; Ortiz-Marquez, J.C.F.; Curatti, L. High lipid productivity of an Ankistrodesmus–Rhizobium artificial consortium. Bioresour. Technol. 2013, 146, 400–407. [Google Scholar] [CrossRef]
- Le Chevanton, M.; Garnier, M.; Bougaran, G.; Schreiber, N.; Lukomska, E.; Bérard, J.-B.; Fouilland, E.; Bernard, O.; Cadoret, J.-P. Screening and selection of growth-promoting bacteria for Dunaliella cultures. Algal Res. 2013, 2, 212–222. [Google Scholar] [CrossRef]
- Choix, F.J.; De-Bashan, L.E.; Bashan, Y. Enhanced accumulation of starch and total carbohydrates in alginate-immobilized Chlorella spp. induced by Azospirillum brasilense: II. Heterotrophic conditions. Enzym. Microb. Technol. 2012, 51, 300–309. [Google Scholar] [CrossRef]
- Rivas, M.O.; Vargas, P.; Riquelme, C.E. Interactions of Botryococcus braunii cultures with bacterial biofilms. Microb. Ecol. 2010, 60, 628–635. [Google Scholar] [CrossRef]
- Butler, T.; Golan, Y. Astaxanthin production from microalgae. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020; pp. 175–242. [Google Scholar]
- Mathimani, T.; Mallick, N. A comprehensive review on harvesting of microalgae for biodiesel—Key challenges and future directions. Renew. Sustain. Energy Rev. 2018, 91, 1103–1120. [Google Scholar] [CrossRef]
- Han, P.; Lu, Q.; Fan, L.; Zhou, W. A review on the use of microalgae for sustainable aquaculture. Appl. Sci. 2019, 9, 2377. [Google Scholar] [CrossRef]
- Levine, I.; Fleurence, J. Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Alam, M.A.; Wang, Z. Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Zhou, W.; Min, M.; Hu, B.; Ma, X.; Liu, Y.; Wang, Q.; Shi, J.; Chen, P.; Ruan, R. Filamentous fungi assisted bio-flocculation: A novel alternative technique for harvesting heterotrophic and autotrophic microalgal cells. Sep. Purif. Technol. 2013, 107, 158–165. [Google Scholar] [CrossRef]
- Chen, J.; Leng, L.; Ye, C.; Lu, Q.; Addy, M.; Wang, J.; Liu, J.; Chen, P.; Ruan, R.; Zhou, W. A comparative study between fungal pellet-and spore-assisted microalgae harvesting methods for algae bioflocculation. Bioresour. Technol. 2018, 259, 181–190. [Google Scholar] [CrossRef]
- Liu, H.; Lu, Q.; Wang, Q.; Liu, W.; Wei, Q.; Ren, H.; Ming, C.; Min, M.; Chen, P.; Ruan, R. Isolation of a bacterial strain, Acinetobacter sp. from centrate wastewater and study of its cooperation with algae in nutrients removal. Bioresour. Technol. 2017, 235, 59–69. [Google Scholar] [CrossRef]
- Naaz, F.; Bhattacharya, A.; Pant, K.K.; Malik, A. Impact of heavy metal laden algal biomass on hydrothermal liquefaction and biorefinery approach. Process Saf. Environ. Prot. 2021, 145, 141–149. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, Z.; Zhu, L.; Ye, T.; Zuo, J.; Li, X.; Xiao, B.; Jin, S. Vertical-algal-biofilm enhanced raceway pond for cost-effective wastewater treatment and value-added products production. Water Res. 2018, 139, 144–157. [Google Scholar] [CrossRef]
- de Farias Neves, F.; Demarco, M.; Tribuzi, G. Drying and quality of microalgal powders for human alimentation. In Microalgae-from Physiology to Application; IntechOpen: London, UK, 2019. [Google Scholar]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Chen, C.-L.; Chang, J.-S.; Lee, D.-J. Dewatering and drying methods for microalgae. Dry. Technol. 2015, 33, 443–454. [Google Scholar] [CrossRef]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Saratale, R.G.; Ferreira, L.F.R.; Bilal, M.; Chandra, R.; Bharagava, R.N. Ecotoxicological and health concerns of persistent coloring pollutants of textile industry wastewater and treatment approaches for environmental safety. J. Environ. Chem. Eng. 2021, 9, 105012. [Google Scholar] [CrossRef]
- Tan, X.B.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Wong, C.Y.; Lee, K.T. Cultivation of microalgae for biodiesel production: A review on upstream and downstream processing. Chin. J. Chem. Eng. 2018, 26, 17–30. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Marticorena, P.; Sepúlveda, C.; Salinas, F.; Cerezal, P.; Riquelme, C. Effect of drying methods on lutein content and recovery by supercritical extraction from the microalga Muriellopsis sp.(MCH35) cultivated in the arid north of Chile. Mar. Drugs 2020, 18, 528. [Google Scholar] [CrossRef] [PubMed]
- Stramarkou, M.; Papadaki, S.; Kyriakopoulou, K.; Krokida, M. Effect of drying and extraction conditions on the recovery of bioactive compounds from Chlorella vulgaris. J. Appl. Phycol. 2017, 29, 2947–2960. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Monteiro, M.; Santos, R.; Iglesias, P.; Couto, A.; Serra, C.; Gouvinhas, I.; Barros, A.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of extraction method and solvent system on the phenolic content and antioxidant activity of selected macro-and microalgae extracts. J. Appl. Phycol. 2020, 32, 349–362. [Google Scholar] [CrossRef]
- Zhang, R.; Parniakov, O.; Grimi, N.; Lebovka, N.; Marchal, L.; Vorobiev, E. Emerging techniques for cell disruption and extraction of valuable bio-molecules of microalgae Nannochloropsis sp. Bioprocess Biosyst. Eng. 2019, 42, 173–186. [Google Scholar] [CrossRef]
- Onay, M.; Sonmez, C.; Oktem, H.A.; Yucel, M. Evaluation of various extraction techniques for efficient lipid recovery from thermo-resistant microalgae, Hindakia, Scenedesmus and Micractinium species—Comparison of lipid extraction methods from microalgae. Am. J. Anal. Chem. 2016, 7, 141. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- De Castro, M.L.; Priego-Capote, F. Soxhlet extraction: Past and present panacea. J. Chromatogr. A 2010, 1217, 2383–2389. [Google Scholar] [CrossRef]
- Koberg, M.; Cohen, M.; Ben-Amotz, A.; Gedanken, A. Bio-diesel production directly from the microalgae biomass of Nannochloropsis by microwave and ultrasound radiation. Bioresour. Technol. 2011, 102, 4265–4269. [Google Scholar] [CrossRef]
- Cui, Y.; Liang, Y. Direct transesterification of wet Cryptococcus curvatus cells to biodiesel through use of microwave irradiation. Appl. Energy 2014, 119, 438–444. [Google Scholar] [CrossRef]
- Cheng, J.; Yu, T.; Li, T.; Zhou, J.; Cen, K. Using wet microalgae for direct biodiesel production via microwave irradiation. Bioresour. Technol. 2013, 131, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Khoiroh, I.; Vo, D.-V.N.; Senthil Kumar, P.; Show, P.L. Techniques of lipid extraction from microalgae for biofuel production: A review. Environ. Chem. Lett. 2021, 19, 231–251. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.-H.; Chen, C.-N.N.; Chen, C.-Y.; Jing, K.; Ng, I.-S.; Chen, J.; Chang, J.-S.; Lu, Y. Disruption of thermo-tolerant Desmodesmus sp. F51 in high pressure homogenization as a prelude to carotenoids extraction. Biochem. Eng. J. 2016, 109, 243–251. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.; Zhang, J. Science of Marine Shellfish Culture; Ocean University of China: Qingdao, China, 2008. [Google Scholar]
- Rivera, E.C.; Montalescot, V.; Viau, M.; Drouin, D.; Bourseau, P.; Frappart, M.; Monteux, C.; Couallier, E. Mechanical cell disruption of Parachlorella kessleri microalgae: Impact on lipid fraction composition. Bioresour. Technol. 2018, 256, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Yin, J.; Gao, Z.; Huang, H.; Ji, X.; Dou, C. Disruption of Chlorella vulgaris cells for the release of biodiesel-producing lipids: A comparison of grinding, ultrasonication, bead milling, enzymatic lysis, and microwaves. Appl. Biochem. Biotechnol. 2011, 164, 1215–1224. [Google Scholar] [CrossRef]
- Greenly, J.M.; Tester, J.W. Ultrasonic cavitation for disruption of microalgae. Bioresour. Technol. 2015, 184, 276–279. [Google Scholar] [CrossRef]
- Lee, A.K.; Lewis, D.M.; Ashman, P.J. Microalgal cell disruption by hydrodynamic cavitation for the production of biofuels. J. Appl. Phycol. 2015, 27, 1881–1889. [Google Scholar] [CrossRef]
- Khedkar, S.V.; Chavan, Y.; Bhagat, S. Extraction of oil from algae by solvent extraction and oil expeller method. Sadguru Publ. 2011, 9, 1746–1750. [Google Scholar]
- Thoisen, C.; Hansen, B.W.; Nielsen, S.L. A simple and fast method for extraction and quantification of cryptophyte phycoerythrin. MethodsX 2017, 4, 209–213. [Google Scholar] [CrossRef] [PubMed]
- González-González, L.M.; Astals, S.; Pratt, S.; Jensen, P.D.; Schenk, P.M. Impact of osmotic shock pre-treatment on microalgae lipid extraction and subsequent methane production. Bioresour. Technol. Rep. 2019, 7, 100214. [Google Scholar] [CrossRef]
- Günerken, E.; D’hondt, E.; Eppink, M.H.; Wijffels, R.H.; Elst, K. Disruption of microalgae with a novel continuous explosive decompression device. Algal Res. 2019, 39, 101376. [Google Scholar] [CrossRef]
- Gim, G.H.; Kim, S.W. Optimization of cell disruption and transesterification of lipids from Botryococcus braunii LB572. Biotechnol. Bioprocess Eng. 2018, 23, 550–556. [Google Scholar] [CrossRef]
- Sheng, J.; Vannela, R.; Rittmann, B. Disruption of Synechocystis PCC 6803 for lipid extraction. Water Sci. Technol. 2012, 65, 567–573. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K. Influence of acid pre-treatment on microalgal biomass for bioethanol production. Process Biochem. 2011, 46, 304–309. [Google Scholar] [CrossRef]
- Sert, B.Ş.; İnan, B.; Özçimen, D. Effect of chemical pre-treatments on bioethanol production from Chlorella minutissima. Acta Chim. Slov. 2018, 65, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Zuorro, A.; Maffei, G.; Lavecchia, R. Optimization of enzyme-assisted lipid extraction from Nannochloropsis microalgae. J. Taiwan Inst. Chem. Eng. 2016, 67, 106–114. [Google Scholar] [CrossRef]
- Armenta, S.; Garrigues, S.; Esteve-Turrillas, F.A.; de la Guardia, M. Green extraction techniques in green analytical chemistry. TrAC Trends Anal. Chem. 2019, 116, 248–253. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Yoo, C.; Jun, S.-Y.; Ahn, C.-Y.; Oh, H.-M. Comparison of several methods for effective lipid extraction from microalgae. Bioresour. Technol. 2010, 101, S75–S77. [Google Scholar] [CrossRef]
- Chu, W.-L.; Lim, Y.-W.; Radhakrishnan, A.K.; Lim, P.-E. Protective effect of aqueous extract from Spirulina platensis against cell death induced by free radicals. BMC Complement. Altern. Med. 2010, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Jin, E.S.; Melis, A. Microalgal biotechnology: Carotenoid production by the green algae Dunaliella salina. Biotechnol. Bioprocess Eng. 2003, 8, 331–337. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Reis, A.; Gouveia, L.; Veloso, V.; Fernandes, H.L.; Empis, J.; Novais, J.M. Eicosapentaenoic acid-rich biomass production by the microalga Phaeodactylum tricornutum in a continuous-flow reactor. Bioresour. Technol. 1996, 55, 83–88. [Google Scholar] [CrossRef]
- Asgharpour, M.; Rodgers, B.; Hestekin, J.A. Eicosapentaenoic acid from Porphyridium cruentum: Increasing growth and productivity of microalgae for pharmaceutical products. Energies 2015, 8, 10487–10503. [Google Scholar] [CrossRef]
- Mendes, A.; Reis, A.; Vasconcelos, R.; Guerra, P.; Lopes da Silva, T. Crypthecodinium cohnii with emphasis on DHA production: A review. J. Appl. Phycol. 2009, 21, 199–214. [Google Scholar] [CrossRef]
- Forján Lozano, E.; Garbayo Nores, I.; Casal Bejarano, C.; Vílchez Lobato, C. Enhancement of carotenoid production in Nannochloropsis by phosphate and sulphur limitation. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Formatex: Badajoz, Spain, 2007. [Google Scholar]
- Mišurcová, L.; Škrovánková, S.; Samek, D.; Ambrožová, J.; Machů, L. Health benefits of algal polysaccharides in human nutrition. Adv. Food Nutr. Res. 2012, 66, 75–145. [Google Scholar]
- Rahman, K.M. Food and high value products from microalgae: Market opportunities and challenges. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Singapore, 2020; pp. 3–27. [Google Scholar]
- Eckelberry, R. Algae—Food or chemical grade. Algae Ind. Mag. 2011. Available online: http://www.algaeindustrymagazine.com/algae-business-algae-food-or-chemical-grade/ (accessed on 29 July 2022).
- Taufiqurrahmi, N.; Religia, P.; Mulyani, G.; Suryana, D.; Tanjung, F.; Arifin, Y. Phycocyanin extraction in Spirulina produced using agricultural waste. Proc. IOP Conf. Ser. Mater. Sci. Eng. 2017, 296, 012097. [Google Scholar] [CrossRef]
- Zanella, L.; Alam, M. Extracts and bioactives from microalgae (sensu stricto): Opportunities and challenges for a new generation of cosmetics. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Singapore, 2020; pp. 295–349. [Google Scholar]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Mittermaier, M.; Pickerodt, P.; Kurth, F.; de Jarcy, L.B.; Uhrig, A.; Garcia, C.; Machleidt, F.; Pergantis, P.; Weber, S.; Li, Y. Evaluation of PEEP and prone positioning in early COVID-19 ARDS. EClinicalMedicine 2020, 28, 100579. [Google Scholar] [CrossRef] [PubMed]
- Khavari, F.; Saidijam, M.; Taheri, M.; Nouri, F. Microalgae: Therapeutic potentials and applications. Mol. Biol. Rep. 2021, 48, 4757–4765. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Dwivedi, V.; Sanyal, D.; Dasgupta, S. Therapeutic and nutritional potential of Spirulina in combating COVID-19 infection. AIJR Preprints 2020, 49, 2–8. [Google Scholar]
- Bao, J.; Li, C.; Zhang, K.; Kang, H.; Chen, W.; Gu, B. Comparative analysis of laboratory indexes of severe and non-severe patients infected with COVID-19. Clin. Chim. Acta 2020, 509, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, H.; Zhan, M.; Jiang, J.; Yin, H.; Dauphars, D. Preventing mortality in COVID-19 patients: Which cytokine to target in a raging storm? Front. Cell. Dev. Biol. 2020, 8, 677. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of theCytokine Storm’in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Shimizu, M. Clinical features of cytokine storm syndrome. In Cytokine Storm Syndrome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 31–41. [Google Scholar]
- Wan Afifudeen, C.L.; Teh, K.Y.; Cha, T.S. Bioprospecting of microalgae metabolites against cytokine storm syndrome during COVID-19. Mol. Biol. Rep. 2021, 49, 1475–1490. [Google Scholar] [CrossRef]
- Moldoveanu, B.; Otmishi, P.; Jani, P.; Walker, J.; Sarmiento, X.; Guardiola, J.; Saad, M.; Yu, J. Inflammatory mechanisms in the lung. J. Inflamm. Res. 2009, 2, 1. [Google Scholar]
- Chia, W.Y.; Kok, H.; Chew, K.W.; Low, S.S.; Show, P.L. Can algae contribute to the war with Covid-19? Bioengineered 2021, 12, 1226–1237. [Google Scholar] [CrossRef]
- Barrantes, M.G.; Espinoza, O.G.; Montero, K.N. Microalgae-based approaches to overcome the effects of the COVID-19 pandemic. Tecnol. Marcha 2022, 35, 84–93. [Google Scholar]
- Norambuena, F.; Hermon, K.; Skrzypczyk, V.; Emery, J.A.; Sharon, Y.; Beard, A.; Turchini, G.M. Algae in fish feed: Performances and fatty acid metabolism in juvenile Atlantic salmon. PLoS ONE 2015, 10, e0124042. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine bioactive compounds and their health benefits: A review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. Thessalon. 2014, 21, 6. [Google Scholar] [CrossRef]
- Wang, J.; Hu, X.; Chen, J.; Wang, T.; Huang, X.; Chen, G. The Extraction of β-Carotene from Microalgae for Testing Their Health Benefits. Foods 2022, 11, 502. [Google Scholar] [CrossRef]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Mathur, M. Bioactive Molecules of: A Food Supplement. In Bioactive Molecules in Food; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1621–1642. [Google Scholar]
- Morais, M.G.d.; Vaz, B.d.S.; Morais, E.G.d.; Costa, J.A.V. Biological effects of Spirulina (Arthrospira) biopolymers and biomass in the development of nanostructured scaffolds. BioMed Res. Int. 2014, 2014, 762705. [Google Scholar] [CrossRef]
- de Sousa e Silva, A.; de Magalhaes, W.T.; Moreira, L.M.; Rocha, M.V.P.; Bastos, A.K.P. Microwave-assisted extraction of polysaccharides from Arthrospira (Spirulina) platensis using the concept of green chemistry. Algal Res. 2018, 35, 178–184. [Google Scholar] [CrossRef]
- Nuhu, A.A. Spirulina (Arthrospira): An important source of nutritional and medicinal compounds. J. Mar. Biol. 2013, 2013, 325636. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, C.; Ajeesh, C.M.; Balaji, S.; Shalini, M.; Ramamoorthy, S.; Ranjan, D.; Fulzele, D.P.; Kalaivani, T. Effect of modified Zarrouk’s medium on growth of different Spirulina strains. Walailak J. Sci. Technol. 2016, 13, 67–75. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Transformation Methods | Microalgae Species | Refs. |
|---|---|---|
| PEG mediated | Pleurochrysis carterae | [85] |
| Dunaliella salina | [86] | |
| Enzyme-Mediated | Chlorella vulgaris | [87] |
| Silicon carbide fibers | Amphidinium sp. | [88] |
| Chlamydomonas reinhardtii | [89] | |
| Electroporation | Chlamydomonas reinhardtii | [90] |
| Chlorella sp. | [75] | |
| Nannochloropsis oculata | [78] | |
| Glass beads | Dunaliella salina | [73] |
| Chlamydomonas reinhardtii | [91] | |
| Platymonas subcordiformis | [92] | |
| Agrobacterium-mediated | Chlorella vulgaris | [76] |
| Haematococcus pluvialis | [93] | |
| Nannochloropsis sp. | [74] | |
| Microprojectile bombardment | Chlorella zofingiensis | [77] |
| Phaeodactylum tricornutum | [94] | |
| Chaetoceros sp. | [95] |
| Properties | Open Pond Cultivation | Closed Pond Cultivation |
|---|---|---|
| Required space | High | Low |
| Water loss | Very high | Low |
| Biomass quality | Variable | Variable and able to increase |
| Biomass concentration (g L−1) | Low (0.1 and 0.5) | High (0.5 and 8) |
| Efficiency of light utilization | Low | Excellent |
| Temperature | Variable | Controlled |
| CO2 loss | High (depend on pond depth) | Low (controlled) |
| Contamination | High | Low |
| Cleaning | None | Required |
| Process control | Limited | Possible |
| Weather dependence | High | Low |
| Start-up | 1.5–2 Months | 0.5–1 Month |
| Capital expenses | High | higher |
| Operating costs | Low | High |
| Efficiency of harvesting | Low | High |
| Irradiance Stress (μmol m−2 s−1) | Microalgal Strain | Biomass Production (mg L−1) | Carbohydrate Productivity (mg L−1 Day−1) | Metabolite Improvement (%) | Refs. |
|---|---|---|---|---|---|
| 890 | S. obtusiusculus | 978 | 280 | 31 | [124] |
| 300 | C. sorokiniana | 2800 | 170 | - | [125] |
| 650 | Tetradesmus obliquus | 1700 | 800 | 30 | [126] |
| 310 | Desmodesmus sp. | 2380 | - | 13.4 | [127] |
| 2000 | B. braunii | 1300 | 900 | - | [128] |
| Nutrient Limitation/Starvation | Microalgal Strain | Carbohydrate Productions (mg L−1) | Metabolite Improvements (%) | Biomass Productions (mg L−1) | Refs. |
|---|---|---|---|---|---|
| Nitrogen | A. platensis | 4.3 | 9.36 | 192 | [134] |
| Nitrogen | M. aeruginosa | - | 20 | 2.25 × 107 cell mL−1 | [135] |
| Phosphorus | A. platensis | 6.31 | 59.7 | 195 | [134] |
| Sulfur | C. reinhardtii | 5070 | 51 | - | [136] |
| vitamins, N, P, and metal | Tetraselmis sp. | 420 mg g−1 | 130 | 5720 | [137] |
| Calcium and magnesium | C. sorokiniana | 450 | 50 | - | [135] |
| Multiple nutrients | Desmodesmus sp. | 400 mg g−1 | 64 | 1950 | [138] |
| Stress Conditions | Microalgal Strain | Biomass Production (mg L−1) | Stress on Carbohydrate Content (%) | Stress on Protein Content (%) | Refs. | ||
|---|---|---|---|---|---|---|---|
| Before | After | Before | After | ||||
| 0.9 g L−1 NaHCO3 | Scenedesmus sp. | 28.32 | 18.5 | 31 | 47 | 49.5 | [140] |
| 5% Pentose | C. minutissima | 60 | 32.5 | 58.5 | 15.5 | 14 | [129] |
| 5% CO2 | A. quadricellulare | 900 mg L−1 | 31 | 71 | 20 | 14 | [141] |
| 5% CO2 | C. sorokiniana | 960 mg L−1 | 30.2 | 53 | 24 | 10 | [141] |
| Animal Type | TN (mg L−1) | NH3-N (mg L−1) | TP (mg L−1) | COD (mg L−1) | TS (g L−1) | Refs. |
|---|---|---|---|---|---|---|
| Shrimp | 361 | 90 | NA | 1321 | NA | [146] |
| Shrimp | >365 | 84 | NA | 1593 | NA | [147] |
| Shrimp | >395 | 102 | NA | 1201 | 13 | [148] |
| Rainbow trout | 1.2 | 0.27 | 0.2 | 17.6 | 0.01 | [149] |
| Crucian carp | 6 | 1 | >0.7 | NA | NA | [150] |
| nd | 111 | 0.07 | NA | 20 | NA | [151] |
| nd | 778 | 50 | 384 | 349 | 20 | [150] |
| Microalgae Species | Wastewater Type | Biomass Productivity (mg dw L−1 d−1) | Refs. |
|---|---|---|---|
| B. braunii | Municipal wastewater | 345.6 | [155,156] |
| S. obliquus | Municipal wastewater | 26 | [155,156] |
| P. carterae | Industrial wastewater | 33 | [155,156] |
| B. braunii | Industrial wastewater | 34 | [155,156] |
| C. pyrenoidosa | Industrial wastewater | 8.114 | [155,156] |
| C. saccharophila | Industrial wastewater | 23 | [155,156] |
| D. tertiolecta | Industrial wastewater | 28 | [155,156] |
| Chlorella sp. | Industrial wastewater | 0.00005 | [157] |
| C. vulgaris | Industrial wastewater | 0.0019 | [158] |
| Chlorella sp. | Agricultural wastewater | 81.4 | [155,156] |
| Neochloris sp. | Industrial wastewater | 0.109 | [159] |
| C. minutus | Synthetic wastewater | 2.04 | [160] |
| G. pleurocapsoides | Synthetic wastewater | 3.34 | [160] |
| Microalgal Strain | Bacterium Strain | Interactions (Microalgae Enhancement) | Refs. |
|---|---|---|---|
| C. sorokiniana | brasilense | 11% increase in cell density | [165] |
| A. protothecoides | A. brasilense | 90% increase in cell density | [165] |
| I. galbana | Alteromonas sp. | 52% increase in cell density | [166] |
| I. galbana | Labrenzia sp. | 71% increase in cell density | [166] |
| P. tricornutum | Stappia sp. | 72%, 144%, and 172% increase in growth, chlorophylls, and pigment, respectively | [166] |
| C. vulgaris | S. smaltophilia | 18%, 20%, and 22% increase in productivity, growth rate, and biomass, respectively | [167] |
| T. striata | P. bermudensis | 200% increase in biomass productivity | [168] |
| C. minutissima | Escherichia coli | 700% increase in biomass productivity | [169] |
| T. pseudonana | D. shibae | 35% increase in cell density | [170] |
| Ankistrodesmus sp. | Rhizobium sp. | 29% increase in dry weight | [171] |
| Dunaliella sp. | Muricauda sp | 7% increase in cell biovolume | [172] |
| C. sorokiniana | brasilense | 40% and 35% increase in cell density and growth rate, respectively | [173] |
| C. vulgaris | brasilense | 16% and 11% increase in cell density and growth rate, respectively | [173] |
| B. braunii | Rhizobium sp. | 55% increase in optical density | [174] |
| Method | Advantages | Disadvantages |
|---|---|---|
| Solar | Sustainable and no energy consumption | Dependence on the weather |
| Spray | Fast and economical method, suitable for algae production for human consumption | Degradation in the quality, operational cost |
| Freeze | Highly energy intensive | Applicable for small scale |
| Oven drying | Less energy intensive | Suitable for small scale |
| Crossflow air-drying | Economical and fast drying | Energy cost |
| Incinerator | Algal biomass burning can be avoided | High cost and complicated |
| Category | Technique | Mechanism | Refs. |
|---|---|---|---|
| Mechanical | High-pressure homogenization | When cells are pushed to flow via a narrow valve under tremendous pressure, they break. | [203] |
| Rotor–stator homogenization | Cells are disrupted by the shearing between a fixed outer stator and a rapid-spinning inner rotor when they are drawn into a long shaft | [204] | |
| Bead milling | Contact of cells with agitated beads crushes them. | [205] | |
| Grinding with mortar and pestle | Cells are crushed when they are cut between two hard surfaces, such as a stationary mortar and a rotating pestle. Before the grinding procedure, the sample is frozen by submerging it in liquid nitrogen. | [206] | |
| Ultrasonication | The cavitation, which is created by high-frequency sound waves, causes the cells to explode | [207] | |
| Hydrodynamic cavitation | Cells break down by the cavitation caused by a rapid shift in pressure | [208] | |
| Screw expeller pressing | When the dried cells are pressed through a barrel-like chamber, a large volume of them is crushed. Direct oil extraction is possible with this technique. | [209] | |
| Physical | Repeated freeze-thaw | Production of intracellular ice crystals during the freezing process and cell expansion during the thawing process both cause cell disruption. | [210] |
| Osmotic shock | When fluid comes into cells rapidly during a sudden osmotic transition, internal pressure builds up in the cells, causing the cells to burst. | [211] | |
| Explosive decompression | Gas bubbles escape out from the cells in point holes when pressure is released suddenly | [212] | |
| Pulsed electric field (PEF) | The effect of electroporation, caused by strong electric fields, causes cells to lysis | [177] | |
| Microwave | The intrusion of water vapor within the cells causes the cells to be disturbed. Microwaves create a fast fluctuating electric field that leads to the development of heat due to the frictional forces by inert-molecular movement. | [213] | |
| Thermolysis (autoclave) | When cells are heated to 121 °C for 30 min, they are lysed. Heat is transferred from the outside to the inside of the cells via the cell membrane. | [214] | |
| Chemical | Alkalis or acids | The cell membrane is solubilized through saponification with an alkali or acid | [215] |
| Detergent(surfactant) | Detergent chemicals cause cellular disruption by dissolving cell membrane proteins. | [216] | |
| Enzyme E | Through the process of enzymatic hydrolysis, the enzyme digests the cell wall. | [217] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashour, M.; Omran, A.M.M. Recent Advances in Marine Microalgae Production: Highlighting Human Health Products from Microalgae in View of the Coronavirus Pandemic (COVID-19). Fermentation 2022, 8, 466. https://doi.org/10.3390/fermentation8090466
Ashour M, Omran AMM. Recent Advances in Marine Microalgae Production: Highlighting Human Health Products from Microalgae in View of the Coronavirus Pandemic (COVID-19). Fermentation. 2022; 8(9):466. https://doi.org/10.3390/fermentation8090466
Chicago/Turabian StyleAshour, Mohamed, and Ashraf M. M. Omran. 2022. "Recent Advances in Marine Microalgae Production: Highlighting Human Health Products from Microalgae in View of the Coronavirus Pandemic (COVID-19)" Fermentation 8, no. 9: 466. https://doi.org/10.3390/fermentation8090466
APA StyleAshour, M., & Omran, A. M. M. (2022). Recent Advances in Marine Microalgae Production: Highlighting Human Health Products from Microalgae in View of the Coronavirus Pandemic (COVID-19). Fermentation, 8(9), 466. https://doi.org/10.3390/fermentation8090466