
Effects of Fermented Seaweed Fertilizer Treatment on Paddy Amino Acid Content and Rhizosphere Microbiome Community

 , , and
, , and 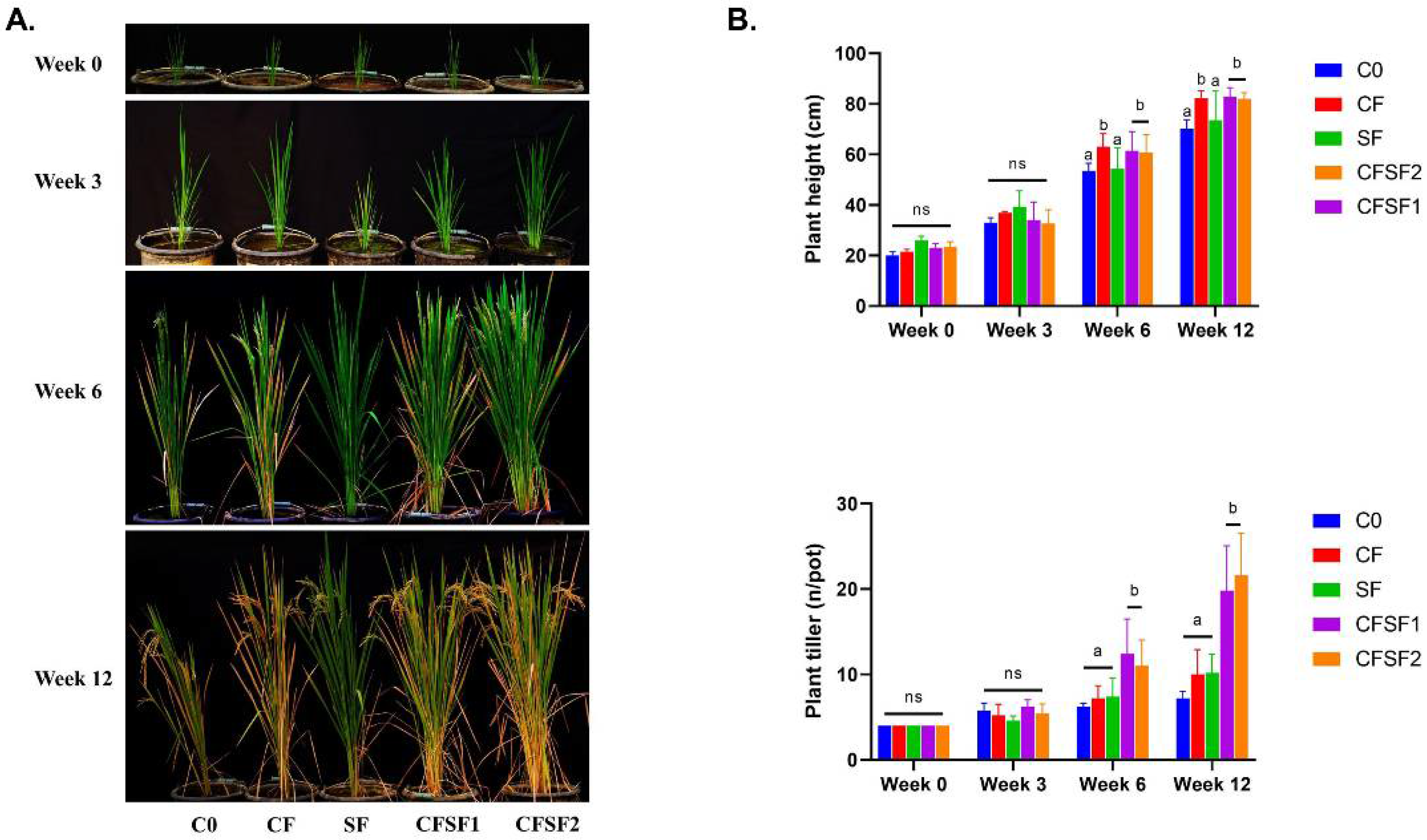{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seaweed Fertilizer Preparation
2.2. Pot Experimental Design
2.3. Determination of Soil, Paddy Leaves, and Rice Grain Macro- and Micronutrient Properties
2.4. Determination of Amino Acid Content in Rice Grain
2.5. Soil Microbial Community Analysis Based on the 16S rRNA Gene
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effects of Seaweed Organic Fertilizer on Growth of Paddy Plants
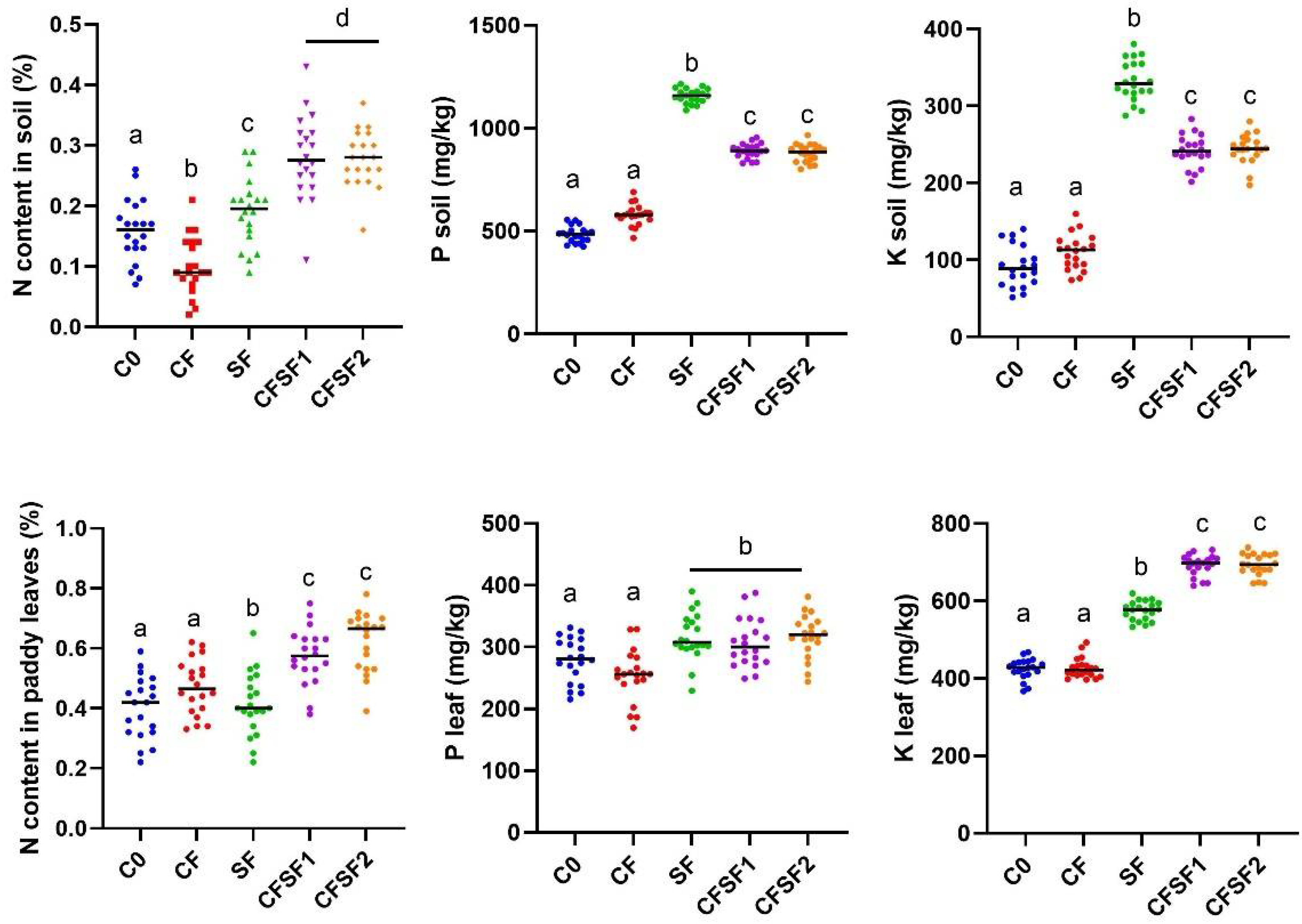
3.2. Effects of Seaweed Fertilizer on Paddy Macronutrient Uptake
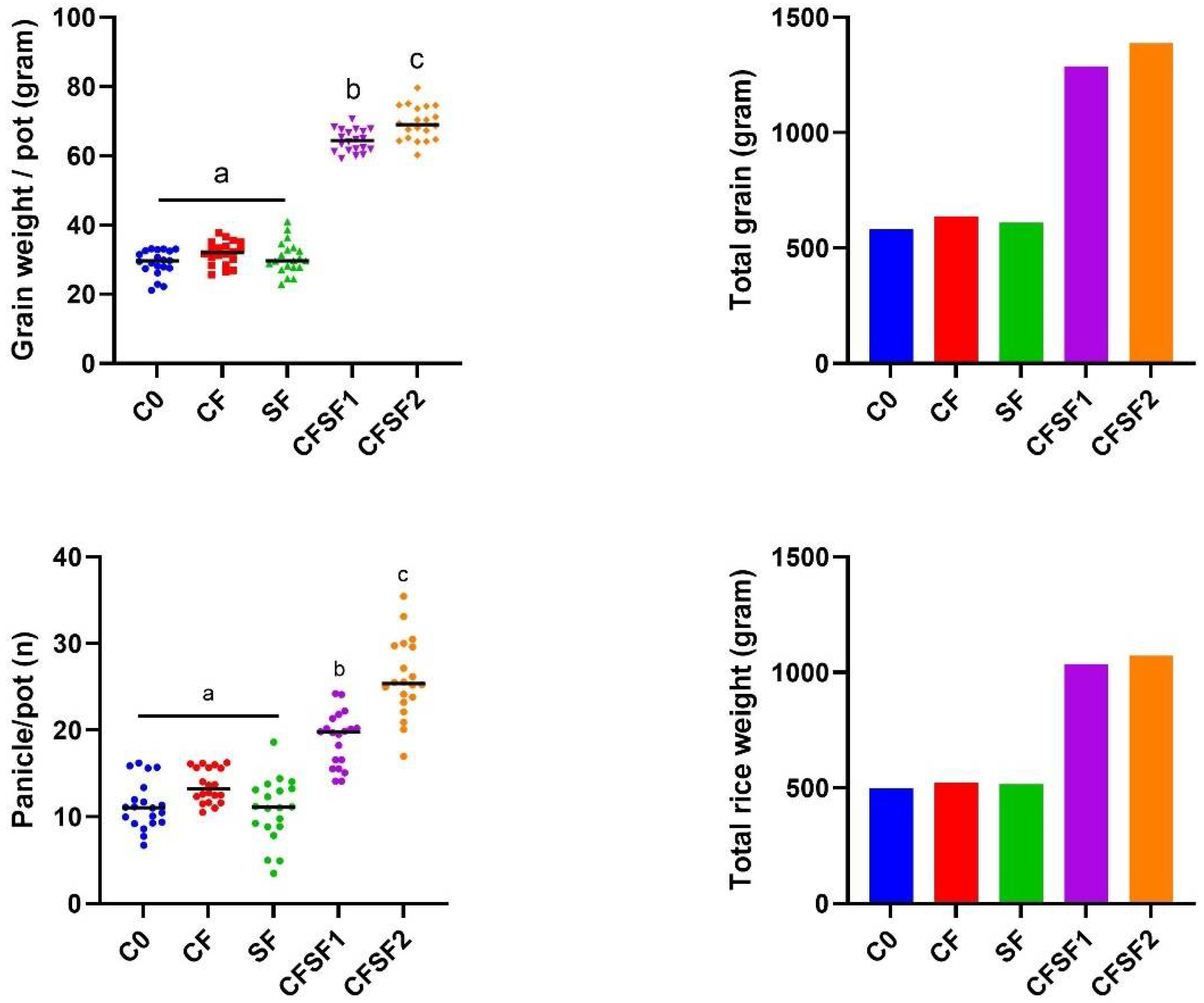
3.3. Effects of Seaweed Fertilizer on Grain Yield
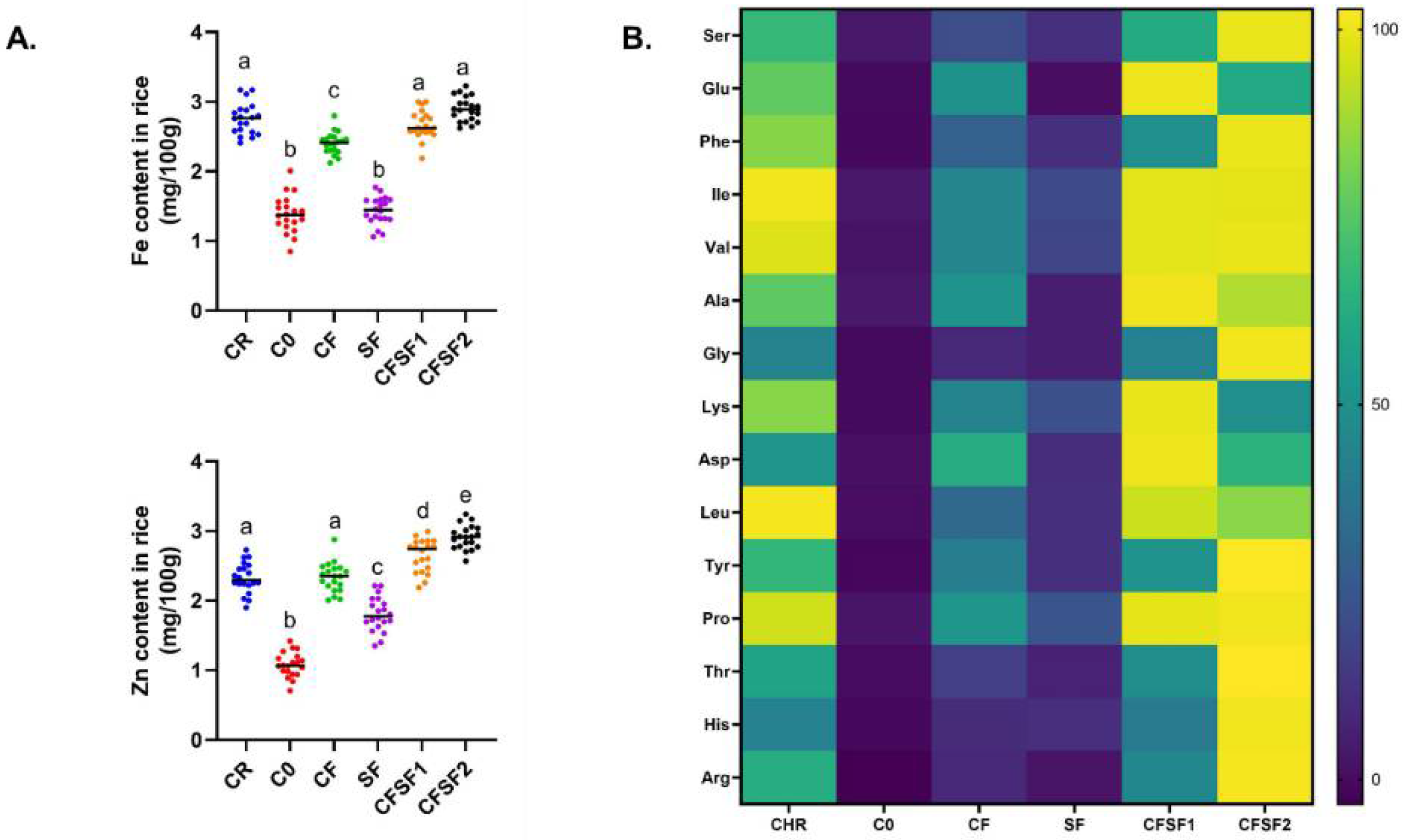
3.4. Effects of Seaweed Fertilizer on Rice Micronutrient and Amino Acid Composition
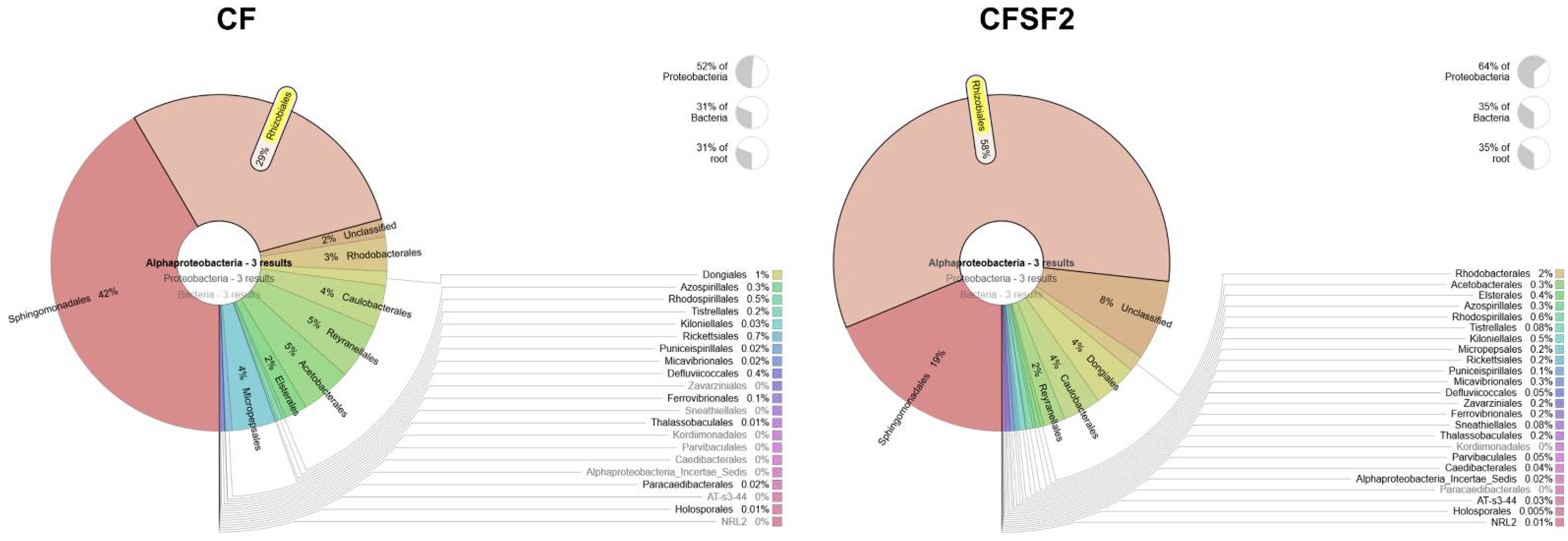
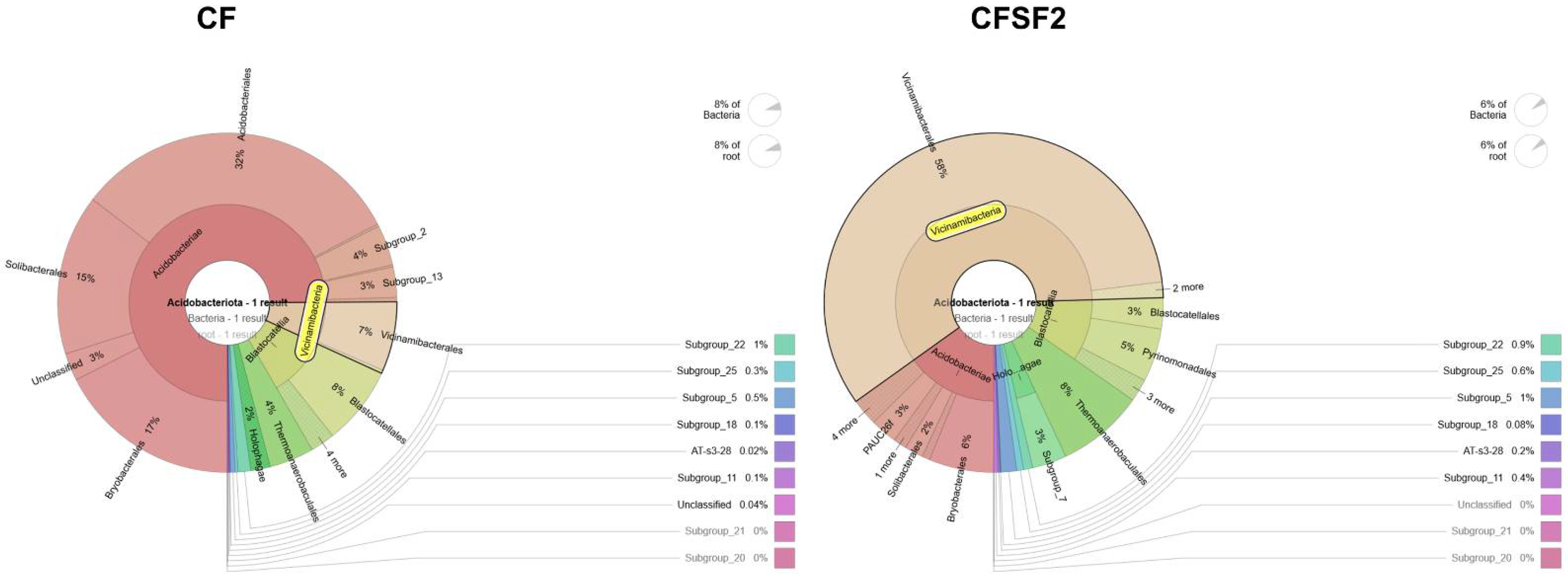
3.5. Soil Microbial Diversity of Rice Grown with Seaweed Fertilizer
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhavadharini, B.; Mohan, V.; Dehghan, M.; Rangarajan, S.; Swaminathan, S.; Rosengren, A.; Wielgosz, A.; Avezum, A.; Lopez-Jaramillo, P.; Lanas, F.; et al. White Rice Intake and Incident Diabetes: A Study of 132,373 Participants in 21 Countries. Diabetes Care 2020, 43, 2643–2650. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cui, W.; Che, Z.; Liang, F.; Wen, Y.; Zhan, M.; Dong, X.; Jin, W.; Dong, Z.; Song, H. Effects of Synthetic Nitrogen Fertilizer and Manure on Fungal and Bacterial Contributions to N2O Production along a Soil Acidity Gradient. Sci. Total Environ. 2021, 753, 142011. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine; Division on Earth and Life Studies; Environmental Health Matters Initiative. Reducing the Health Impacts of the Nitrogen Problem: Proceedings of a Workshop—in Brief; Johnson, A., Laney, K., Eds.; The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press (US): Washington, DC, USA, 2021; ISBN 978-0-309-09412-2. [Google Scholar]
- Salami, R.; Kordi, M.; Bolouri, P.; Delangiz, N.; Asgari Lajayer, B. Algae-Based Biorefinery as a Sustainable Renewable Resource. Circ. Econ. Sustain. 2021, 1, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Shen, Z.; Sun, Y.; Lyu, N.; Ruan, Y.; Li, R.; Shen, Q. Effects of continuous application of bio-organic fertilizer on banana production and cultural microflora of bulk soil in orchard with serious disease incidence. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2015, 26, 481–489. [Google Scholar]
- Du, S.; Ma, Z.; Chen, J.; Xue, L.; Tang, C.; Shareef, T.M.E.; Siddique, K.H.M. Effects of Organic Fertilizer Proportion on the Distribution of Soil Aggregates and Their Associated Organic Carbon in a Field Mulched with Gravel. Sci. Rep. 2022, 12, 11513. [Google Scholar] [CrossRef]
- Salam, M.A.; Sarker, M.N.I.; Sharmin, S. Do Organic Fertilizer Impact on Yield and Efficiency of Rice Farms? Empirical Evidence from Bangladesh. Heliyon 2021, 7, e07731. [Google Scholar] [CrossRef]
- Sunarpi, H.; Kurnianingsih, R.; Ghazali, M.; Fanani, R.A.; Sunarwidhi, A.L.; Widyastuti, S.; Nikmatullah, A.; Prasedya, E.S. Evidence for the Presence of Growth-Promoting Factors in Lombok Turbinaria Murayana Extract Stimulating Growth and Yield of Tomato Plants (Lycopersicum esculentum Mill.). J. Plant Nutr. 2020, 43, 1813–1823. [Google Scholar] [CrossRef]
- Cen, Y.; Guo, L.; Liu, M.; Gu, X.; Li, C.; Jiang, G. Using Organic Fertilizers to Increase Crop Yield, Economic Growth, and Soil Quality in a Temperate Farmland. PeerJ 2020, 8, e9668. [Google Scholar] [CrossRef]
- Elsallam, M.E.A.; El-Moslamy, S.H.; El-Al, A.A.; Zahran, H.F. Scaling-up Production of Cost-Effective and Eco-Friendly Bio-Fertilizer and Its Application on Barley Green Fodder via IoT Hydroponic System. J. Genet. Eng. Biotechnol. 2021, 19, 97. [Google Scholar] [CrossRef]
- Deniz, F.; Ersanli, E.T. An Ecofriendly Approach for Bioremediation of Contaminated Water Environment: Potential Contribution of a Coastal Seaweed Community to Environmental Improvement. Int. J. Phytoremediation 2018, 20, 256–263. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; van der Werf, A.; Karlova, R. The Power of Seaweeds as Plant Biostimulants to Boost Crop Production under Abiotic Stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef] [PubMed]
- Fatimi, A. Seaweed-Based Biofertilizers: A Patent Analysis. Recent Pat. Biotechnol. 2022, 16, 144–154. [Google Scholar] [CrossRef] [PubMed]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Liu, J.; Li, S.; Chen, Y.; Du, P.; Zhu, Y.; Liao, Y.; Chen, Q.; Shou, L.; Yan, X.; et al. Kelp Cultivation Effectively Improves Water Quality and Regulates Phytoplankton Community in a Turbid, Highly Eutrophic Bay. Sci. Total Environ. 2020, 707, 135561. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.U.; Hasan, O.; Rahman, M.M.; Akter, M.; Rahman, M.S.; Sarker, S. Seaweeds for the Sustainable Blue Economy Development: A Study from the South East Coast of Bangladesh. Heliyon 2022, 8, e09079. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Kurniawan, N.S.H.; Kirana, I.A.P.; Ardiana, N.; Abidin, A.S.; Ilhami, B.T.K.; Jupri, A.; Widyastuti, S.; Sunarpi, H.; Nikmatullah, A. Seaweed Fertilizer Prepared by EM-Fermentation Increases Abundance of Beneficial Soil Microbiome in Paddy (Oryzasativa L.) during Vegetative Stage. Fermentation 2022, 8, 46. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, Y.; Jiang, Y. Amino Acids in Rice Grains and Their Regulation by Polyamines and Phytohormones. Plants 2022, 11, 1581. [Google Scholar] [CrossRef]
- de Koning, T.J. Amino Acid Synthesis Deficiencies. J. Inherit. Metab. Dis. 2017, 40, 609–620. [Google Scholar] [CrossRef]
- Titaley, C.R.; Ariawan, I.; Hapsari, D.; Muasyaroh, A.; Dibley, M.J. Determinants of the Stunting of Children under Two Years Old in Indonesia: A Multilevel Analysis of the 2013 Indonesia Basic Health Survey. Nutrients 2019, 11, 1106. [Google Scholar] [CrossRef]
- Wang, M.; Yan, P.; Wang, X. Microbial Fermentation for Algal Fertilizer Production. In Encyclopedia of Marine Biotechnology; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 2247–2260. ISBN 978-1-119-14380-2. [Google Scholar]
- Nasmia; Rosyida, E.; Masyahoro, A.; Putera, F.H.A.; Natsir, S. The Utilization of Seaweed-Based Liquid Organic Fertilizer to Stimulate Gracilaria Verrucosa Growth and Quality. Int. J. Environ. Sci. Technol. 2021, 18, 1637–1644. [Google Scholar] [CrossRef]
- Bzdyk, R.M.; Olchowik, J.; Studnicki, M.; Oszako, T.; Sikora, K.; Szmidla, H.; Hilszczańska, D. The Impact of Effective Microorganisms (EM) and Organic and Mineral Fertilizers on the Growth and Mycorrhizal Colonization of Fagus Sylvatica and Quercus Robur Seedlings in a Bare-Root Nursery Experiment. Forests 2018, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Budiono, R.; Adinurani, P.G.; Soni, P. Effect of New NPK Fertilizer on Lowland Rice (Oryza Sativa L.) Growth. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 2nd International Conference on Natural Resources and Life Sciences (NRLS-2018), Surabaya, Indonesia, 23–24 August 2018; IOP Publishing: Bristol, UK, 2019; Volume 293, p. 012034. [Google Scholar] [CrossRef]
- Chen, L.; Flynn, D.F.B.; Jing, X.; Kühn, P.; Scholten, T.; He, J.-S. A Comparison of Two Methods for Quantifying Soil Organic Carbon of Alpine Grasslands on the Tibetan Plateau. PLoS ONE 2015, 10, e0126372. [Google Scholar] [CrossRef] [PubMed]
- Walkley, A.; Black, I.A. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Kleiber, T.; Krzyżaniak, M.; Świerk, D.; Haenel, A.; Gałecka, S. How Does the Content of Nutrients in Soil Affect the Health Status of Trees in City Parks? PLoS ONE 2019, 14, e0221514. [Google Scholar] [CrossRef] [PubMed]
- Anusha, G.; Rao, D.S.; Jaldhani, V.; Beulah, P.; Neeraja, C.N.; Gireesh, C.; Anantha, M.S.; Suneetha, K.; Santhosha, R.; Prasad, A.S.H.; et al. Grain Fe and Zn Content, Heterosis, Combining Ability and Its Association with Grain Yield in Irrigated and Aerobic Rice. Sci. Rep. 2021, 11, 10579. [Google Scholar] [CrossRef]
- Li, Z.; Xing, Y.; Guo, X.; Cui, Y. Development of an UPLC-MS/MS Method for Simultaneous Quantitation of 11 d-Amino Acids in Different Regions of Rat Brain: Application to a Study on the Associations of d-Amino Acid Concentration Changes and Alzheimer’s Disease. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1058, 40–46. [Google Scholar] [CrossRef]
- García-Alegría, A.M.; Anduro-Corona, I.; Pérez-Martínez, C.J.; Guadalupe Corella-Madueño, M.A.; Rascón-Durán, M.L.; Astiazaran-Garcia, H. Quantification of DNA through the NanoDrop Spectrophotometer: Methodological Validation Using Standard Reference Material and Sprague Dawley Rat and Human DNA. Int. J. Anal. Chem. 2020, 2020, 8896738. [Google Scholar] [CrossRef]
- Zhou, S.; Hill, C.S.; Clark, M.U.; Sheahan, T.P.; Baric, R.; Swanstrom, R. Primer ID Next-Generation Sequencing for the Analysis of a Broad Spectrum Antiviral Induced Transition Mutations and Errors Rates in a Coronavirus Genome. Bio-Protocol 2021, 11, e3938. [Google Scholar] [CrossRef]
- Sola-Leyva, A.; Andrés-León, E.; Molina, N.M.; Terron-Camero, L.C.; Plaza-Díaz, J.; Sáez-Lara, M.J.; Gonzalvo, M.C.; Sánchez, R.; Ruíz, S.; Martínez, L.; et al. Mapping the Entire Functionally Active Endometrial Microbiota. Hum. Reprod. Oxf. Engl. 2021, 36, 1021–1031. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Guo, S.; Tang, M.; Su, M.; Li, H. Financial Support for Agriculture, Chemical Fertilizer Use, and Carbon Emissions from Agricultural Production in China. Int. J. Environ. Res. Public Health 2022, 19, 7155. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.-J.; Zhang, L.-F.; Liu, Z.-B.; Zhang, W.-X.; Lan, X.-J.; Liu, X.-M.; Liu, J.; Liu, G.-R.; Li, Z.-Z.; Wang, P. Effects of Long-Term Application of Chemical Fertilizers and Organic Fertilizers on Heavy Metals and Their Availability in Reddish Paddy Soil. Huan Jing Ke Xue Huanjing Kexue 2021, 42, 2469–2479. [Google Scholar] [CrossRef] [PubMed]
- Alewell, C.; Ringeval, B.; Ballabio, C.; Robinson, D.A.; Panagos, P.; Borrelli, P. Global Phosphorus Shortage Will Be Aggravated by Soil Erosion. Nat. Commun. 2020, 11, 4546. [Google Scholar] [CrossRef]
- Nedumaran, T. Seaweed: A Fertilizer for Sustainable Agriculture. In Sustainable Agriculture towards Food Security; Dhanarajan, A., Ed.; Springer: Singapore, 2017; pp. 159–174. ISBN 978-981-10-6647-4. [Google Scholar]
- Arioli, T.; Mattner, S.W.; Winberg, P.C. Applications of Seaweed Extracts in Australian Agriculture: Past, Present and Future. J. Appl. Phycol. 2015, 27, 2007–2015. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, X.; Chen, B.; Zhang, M.; Ma, J. Seaweed Extract Improved Yields, Leaf Photosynthesis, Ripening Time, and Net Returns of Tomato (Solanum lycopersicum Mill.). ACS Omega 2020, 5, 4242–4249. [Google Scholar] [CrossRef]
- Vassileva, M.; Malusà, E.; Sas-Paszt, L.; Trzcinski, P.; Galvez, A.; Flor-Peregrin, E.; Shilev, S.; Canfora, L.; Mocali, S.; Vassilev, N. Fermentation Strategies to Improve Soil Bio-Inoculant Production and Quality. Microorganisms 2021, 9, 1254. [Google Scholar] [CrossRef]
- Monteiro, P.; Lomartire, S.; Cotas, J.; Pacheco, D.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry. Processes 2021, 9, 1953. [Google Scholar] [CrossRef]
- Liao, Z.; Yu, H.; Duan, J.; Yuan, K.; Yu, C.; Meng, X.; Kou, L.; Chen, M.; Jing, Y.; Liu, G.; et al. SLR1 Inhibits MOC1 Degradation to Coordinate Tiller Number and Plant Height in Rice. Nat. Commun. 2019, 10, 2738. [Google Scholar] [CrossRef]
- Yang, G.; Xing, Y.; Li, S.; Ding, J.; Yue, B.; Deng, K.; Li, Y.; Zhu, Y. Molecular Dissection of Developmental Behavior of Tiller Number and Plant Height and Their Relationship in Rice (Oryza Sativa L.). Hereditas 2006, 143, 236–245. [Google Scholar] [CrossRef]
- Stirk, W.A.; Van Staden, J. Chapter Five—Plant Growth Regulators in Seaweeds: Occurrence, Regulation and Functions. In Advances in Botanical Research; Plants; Bourgougnon, N., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 71, pp. 125–159. [Google Scholar]
- Saha, M.; Dove, S.; Weinberger, F. Chemically Mediated Microbial “Gardening” Capacity of a Seaweed Holobiont Is Dynamic. Microorganisms 2020, 8, 1893. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Huang, Z.; Su, G.; Li, X.; Sun, Z.; Qin, Y. Impact of Short-Term Application of Seaweed Fertilizer on Bacterial Diversity and Community Structure, Soil Nitrogen Contents, and Plant Growth in Maize Rhizosphere Soil. Folia Microbiol. 2020, 65, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Seo, J.; Koo, Y. Alginate and Fucoidan Changes the Bacterial Community in Different Directions and the Alginate or Fucoidan Degrading Bacteria Isolated from Paddy Soil Promotes the Plant Growth. Arch. Microbiol. 2021, 203, 5183–5192. [Google Scholar] [CrossRef] [PubMed]
- Tursun, A.O. Effect of Foliar Application of Seaweed (Organic Fertilizer) on Yield, Essential Oil and Chemical Composition of Coriander. PLoS ONE 2022, 17, e0269067. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; El-Shershaby, N.A. Algae as Bio-Fertilizers: Between Current Situation and Future Prospective. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef]
- Chen, C.-L.; Song, W.-L.; Sun, L.; Qin, S.; Ren, C.-G.; Yang, J.-C.; Feng, D.-W.; Liu, N.; Yan, J.; Cui, B.-B.; et al. Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages. Agronomy 2022, 12, 342. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Y. The Physiological Response of Photosynthesis to Nitrogen Deficiency. Plant Physiol. Biochem. PPB 2021, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- EL Boukhari, M.E.M.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in Seaweed Extract Based Biostimulants: Manufacturing Process and Beneficial Effect on Soil-Plant Systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef]
- Di Stasio, E.; Rouphael, Y.; Colla, G.; Raimondi, G.; Giordano, M.; Pannico, A.; El-Nakhel, C.; De Pascale, S. Influence of Ecklonia Maxima Seaweed Extract on Growth, Photosynthetic Activity and Mineral Composition of Brassica rapa L. Subsp. Sylvestris under Nutrient Stress Conditions. Eur. J. Hortic. Sci. 2018, 82, 286–293. [Google Scholar] [CrossRef]
- Saravana, P.S.; Choi, J.H.; Park, Y.B.; Woo, H.C.; Chun, B.S. Evaluation of the Chemical Composition of Brown Seaweed (Saccharina japonica) Hydrolysate by Pressurized Hot Water Extraction. Algal Res. 2016, 13, 246–254. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of Phosphorus Deficiency on the Photosynthetic Electron Transport Chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, N.S.H.; Kirana, I.A.P.; Abidin, A.S.; Jupri, A.; Widyastuti, S.; Hernawan, A.; Nikmatullah, A.; Sunarpi, H.; Prasedya, E.S. Analysis of Leaf Chlorophyll Content of Paddy Plants during Vegetative Stage Grown in Soil Media Containing Macroalgae Organic Fertilizer. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 4th International Conference on Bioscience and Biotechnology, Virtual, 16–18 August 2021; IOP Publishing: Bristol, UK, 2021; Volume 913, p. 012025. [Google Scholar] [CrossRef]
- Chen, X.-X.; Zhang, W.; Liang, X.-Y.; Liu, Y.-M.; Xu, S.-J.; Zhao, Q.-Y.; Du, Y.-F.; Zhang, L.; Chen, X.-P.; Zou, C.-Q. Physiological and Developmental Traits Associated with the Grain Yield of Winter Wheat as Affected by Phosphorus Fertilizer Management. Sci. Rep. 2019, 9, 16580. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Lin, Y.-S.; Song, X.-J.; Shen, J.-B.; Huang, W.; Shan, J.-X.; Zhu, M.-Z.; Jiang, L.; Gao, J.-P.; Lin, H.-X. The Novel Quantitative Trait Locus GL3.1 Controls Rice Grain Size and Yield by Regulating Cyclin-T1;3. Cell Res. 2012, 22, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting Nitrogen Metabolism and Transport Processes to Improve Plant Nitrogen Use Efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef]
- Kumar, A.; Sen, A.; Upadhyay, P.K.; Singh, R.K. Effect of Zinc, Iron and Manganese Levels on Quality, Micro and Macro Nutrients Content of Rice and Their Relationship with Yield. Commun. Soil Sci. Plant Anal. 2017, 48, 1539–1551. [Google Scholar] [CrossRef]
- Díaz-Benito, P.; Banakar, R.; Rodríguez-Menéndez, S.; Capell, T.; Pereiro, R.; Christou, P.; Abadía, J.; Fernández, B.; Álvarez-Fernández, A. Iron and Zinc in the Embryo and Endosperm of Rice (Oryza sativa L.) Seeds in Contrasting 2′-Deoxymugineic Acid/Nicotianamine Scenarios. Front. Plant Sci. 2018, 9, 1190. [Google Scholar] [CrossRef]
- Herath, H.M.T.; Rajapakse, D.; Wimalasena, S.; Weerasooriya, M.K.B. Iron Content and Availability Studies in Some Sri Lankan Rice Varieties. Int. J. Food Sci. Technol. 2011, 46, 1679–1684. [Google Scholar] [CrossRef]
- Lin, W.; Lin, M.; Zhou, H.; Wu, H.; Li, Z.; Lin, W. The Effects of Chemical and Organic Fertilizer Usage on Rhizosphere Soil in Tea Orchards. PLoS ONE 2019, 14, e0217018. [Google Scholar] [CrossRef]
- Wu, J.; Wang, H.; Li, G.; Ma, W.; Wu, J.; Gong, Y.; Xu, G. Vegetation Degradation Impacts Soil Nutrients and Enzyme Activities in Wet Meadow on the Qinghai-Tibet Plateau. Sci. Rep. 2020, 10, 21271. [Google Scholar] [CrossRef]
- Gregory, A.S.; Ritz, K.; McGrath, S.P.; Quinton, J.N.; Goulding, K.W.T.; Jones, R.J.A.; Harris, J.A.; Bol, R.; Wallace, P.; Pilgrim, E.S.; et al. A Review of the Impacts of Degradation Threats on Soil Properties in the UK. Soil Use Manag. 2015, 31, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, J.; Zhang, X.; Zhang, S.; Chen, Y. Organic Manure Input Improves Soil Water and Nutrients Use for Sustainable Maize (Zea Mays. L) Productivity on the Loess Plateau. PLoS ONE 2020, 15, e0238042. [Google Scholar] [CrossRef] [PubMed]
- Dahunsi, S.O.; Oranusi, S.; Efeovbokhan, V.E.; Adesulu-Dahunsi, A.T.; Ogunwole, J.O. Crop Performance and Soil Fertility Improvement Using Organic Fertilizer Produced from Valorization of Carica Papaya Fruit Peel. Sci. Rep. 2021, 11, 4696. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, R.A.; Hussein, M.H.; El-Naggar, N.E.-A.; Karim-Eldeen, M.A.; Alamer, K.H.; Saleh, M.A.; Al Masoudi, L.M.; Sharaf, E.M.; El-Azeem, R.M.A. Promoting Effect of Soluble Polysaccharides Extracted from Ulva Spp. on Zea mays L. Growth. Molecules 2022, 27, 1394. [Google Scholar] [CrossRef] [PubMed]
- Kalman, D.S. Amino Acid Composition of an Organic Brown Rice Protein Concentrate and Isolate Compared to Soy and Whey Concentrates and Isolates. Foods 2014, 3, 394–402. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, H.; Zhao, C.; Chen, G.; Zou, Y. Amino Acid Content in Rice Grains Is Affected by High Temperature during the Early Grain-Filling Period. Sci. Rep. 2019, 9, 2700. [Google Scholar] [CrossRef]
- Mahmoud, A.H.; El Anany, A.M. Nutritional and Sensory Evaluation of a Complementary Food Formulated from Rice, Faba Beans, Sweet Potato Flour, and Peanut Oil. Food Nutr. Bull. 2014, 35, 403–413. [Google Scholar] [CrossRef]
- Barłóg, P.; Grzebisz, W.; Łukowiak, R. Fertilizers and Fertilization Strategies Mitigating Soil Factors Constraining Efficiency of Nitrogen in Plant Production. Plants 2022, 11, 1855. [Google Scholar] [CrossRef]
- Goulding, K.W.T. Soil Acidification and the Importance of Liming Agricultural Soils with Particular Reference to the United Kingdom. Soil Use Manag. 2016, 32, 390–399. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; Saber, H.; Alwaleed, E.A.; Tran, L.-S.P. Sargassum Muticum and Jania Rubens Regulate Amino Acid Metabolism to Improve Growth and Alleviate Salinity in Chickpea. Sci. Rep. 2017, 7, 10537. [Google Scholar] [CrossRef]
- Smith, M.R.; Reis Hodecker, B.E.; Fuentes, D.; Merchant, A. Investigating Nutrient Supply Effects on Plant Growth and Seed Nutrient Content in Common Bean. Plants 2022, 11, 737. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Darveekaran Nair, S.S.; Zhu, P.; Li, S.; Huang, S.; Li, X.; Xu, J.; Yang, F. Impact of DNA Extraction Method and Targeted 16S-RRNA Hypervariable Region on Oral Microbiota Profiling. Sci. Rep. 2018, 8, 16321. [Google Scholar] [CrossRef] [PubMed]
- Erlacher, A.; Cernava, T.; Cardinale, M.; Soh, J.; Sensen, C.W.; Grube, M.; Berg, G. Rhizobiales as Functional and Endosymbiontic Members in the Lichen Symbiosis of Lobaria pulmonaria L. Front. Microbiol. 2015, 6, 53. [Google Scholar] [CrossRef]
- Neuvonen, M.-M.; Tamarit, D.; Näslund, K.; Liebig, J.; Feldhaar, H.; Moran, N.A.; Guy, L.; Andersson, S.G.E. The Genome of Rhizobiales Bacteria in Predatory Ants Reveals Urease Gene Functions but no Genes for Nitrogen Fixation. Sci. Rep. 2016, 6, 39197. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Rensing, C.; Han, D.; Xiao, K.-Q.; Dai, Y.; Tang, Z.; Liesack, W.; Peng, J.; Cui, Z.; Zhang, F. Genome-Resolved Metagenomics Reveals Distinct Phosphorus Acquisition Strategies between Soil Microbiomes. mSystems 2022, 7, e01107-21. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Peng, J.; Liu, P.; Bei, Q.; Rensing, C.; Li, Y.; Yuan, H.; Liesack, W.; Zhang, F.; Cui, Z. Metagenomic Insights into Nitrogen and Phosphorus Cycling at the Soil Aggregate Scale Driven by Organic Material Amendments. Sci. Total Environ. 2021, 785, 147329. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Fraga, R. Phosphate Solubilizing Bacteria and Their Role in Plant Growth Promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Poirier, Y.; Jaskolowski, A.; Clúa, J. Phosphate Acquisition and Metabolism in Plants. Curr. Biol. CB 2022, 32, R623–R629. [Google Scholar] [CrossRef]
- Ajmera, I.; Hodgman, T.C.; Lu, C. An Integrative Systems Perspective on Plant Phosphate Research. Genes 2019, 10, 139. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasedya, E.S.; Kurniawan, N.S.H.; Ardiana, N.; Ilhami, B.T.K.; Mulyaningsih, T.; Astuti, S.P.; Jupri, A.; Nikmatullah, A.; Jaya, I.K.D.; Widyastuti, S. Effects of Fermented Seaweed Fertilizer Treatment on Paddy Amino Acid Content and Rhizosphere Microbiome Community. Fermentation 2022, 8, 420. https://doi.org/10.3390/fermentation8090420
Prasedya ES, Kurniawan NSH, Ardiana N, Ilhami BTK, Mulyaningsih T, Astuti SP, Jupri A, Nikmatullah A, Jaya IKD, Widyastuti S. Effects of Fermented Seaweed Fertilizer Treatment on Paddy Amino Acid Content and Rhizosphere Microbiome Community. Fermentation. 2022; 8(9):420. https://doi.org/10.3390/fermentation8090420
Chicago/Turabian StylePrasedya, Eka Sunarwidhi, Nanda Sofian Hadi Kurniawan, Nur Ardiana, Bq Tri Khairina Ilhami, Tri Mulyaningsih, Sri Puji Astuti, Ahmad Jupri, Aluh Nikmatullah, I Komang Damar Jaya, and Sri Widyastuti. 2022. "Effects of Fermented Seaweed Fertilizer Treatment on Paddy Amino Acid Content and Rhizosphere Microbiome Community" Fermentation 8, no. 9: 420. https://doi.org/10.3390/fermentation8090420
APA StylePrasedya, E. S., Kurniawan, N. S. H., Ardiana, N., Ilhami, B. T. K., Mulyaningsih, T., Astuti, S. P., Jupri, A., Nikmatullah, A., Jaya, I. K. D., & Widyastuti, S. (2022). Effects of Fermented Seaweed Fertilizer Treatment on Paddy Amino Acid Content and Rhizosphere Microbiome Community. Fermentation, 8(9), 420. https://doi.org/10.3390/fermentation8090420