
Effect of a Multi-Strain Probiotic on Growth Performance, Lipid Panel, Antioxidant Profile, and Immune Response in Andaman Local Piglets at Weaning

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Area
2.2. Experimental Period
2.3. Experimental Animals
2.4. Source of the Probiotic
2.5. Study Design
2.6. Estimation of Diarrhea Rate in Piglets
2.7. Production Parameters
2.8. Blood Sampling
2.9. Lipid Profile Analysis
2.10. Measurement of Stress Biomarkers
2.11. Antioxidant and Oxidative Profile
2.11.1. Nitric Oxide Assay
2.11.2. Serum Cortisol Assay
2.11.3. Determination of Serum HSPs
2.12. Immune Parameters
2.13. Statistical Analysis
3. Results
3.1. Production Parameters and Diarrhea Incidence
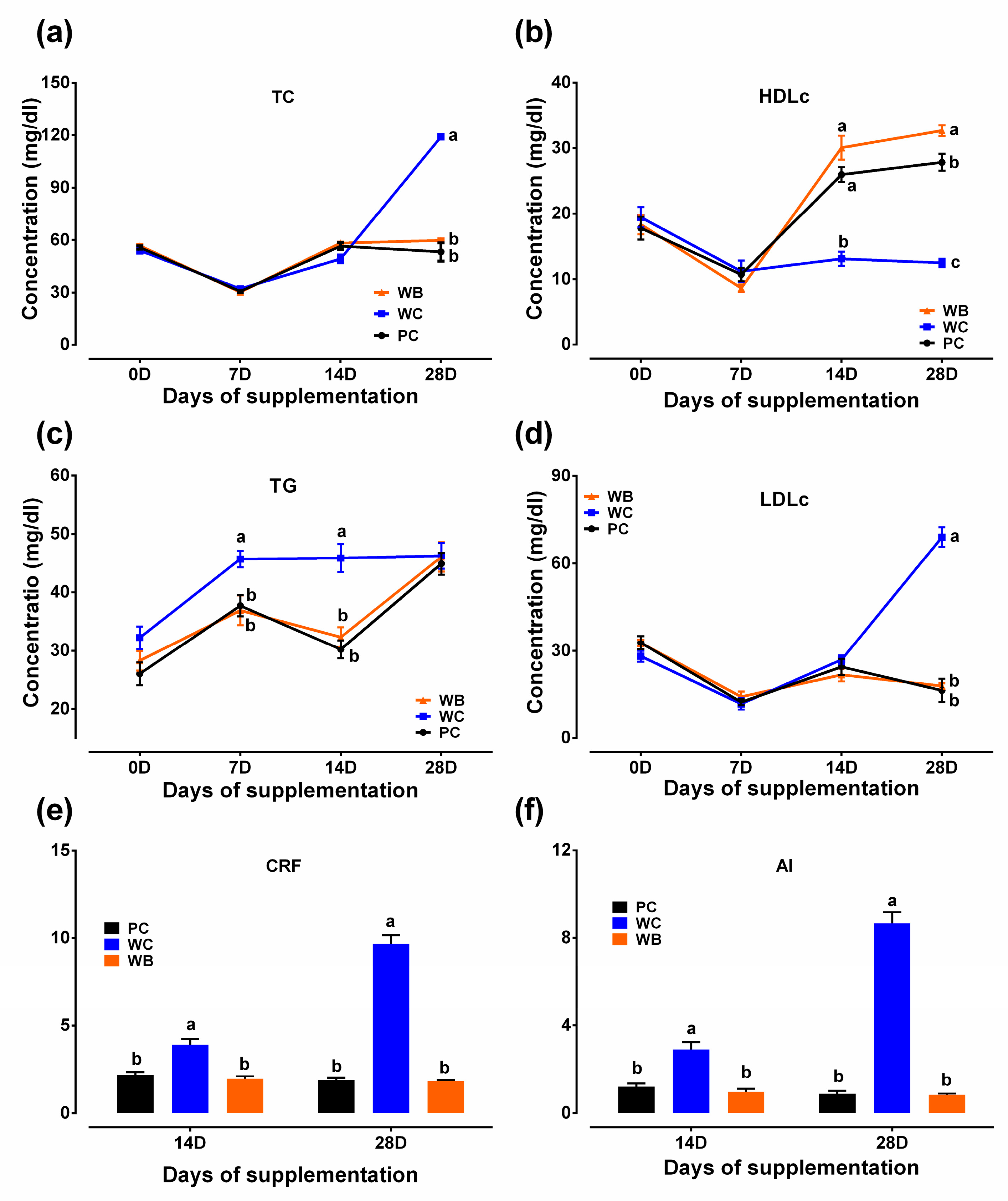
3.2. Lipid Panel Analysis
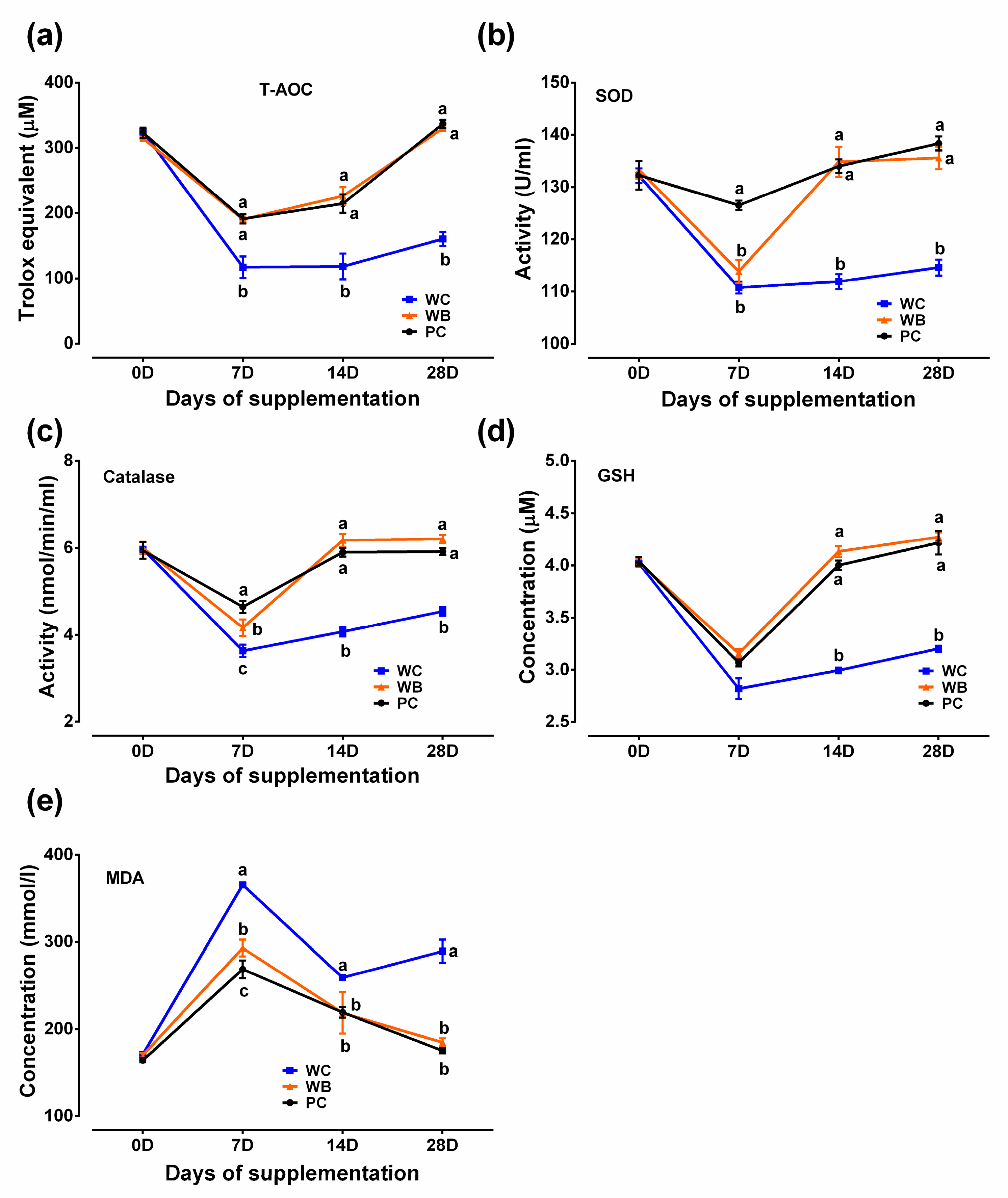
3.3. Antioxidant Profiles and Oxidative Stress Indicators
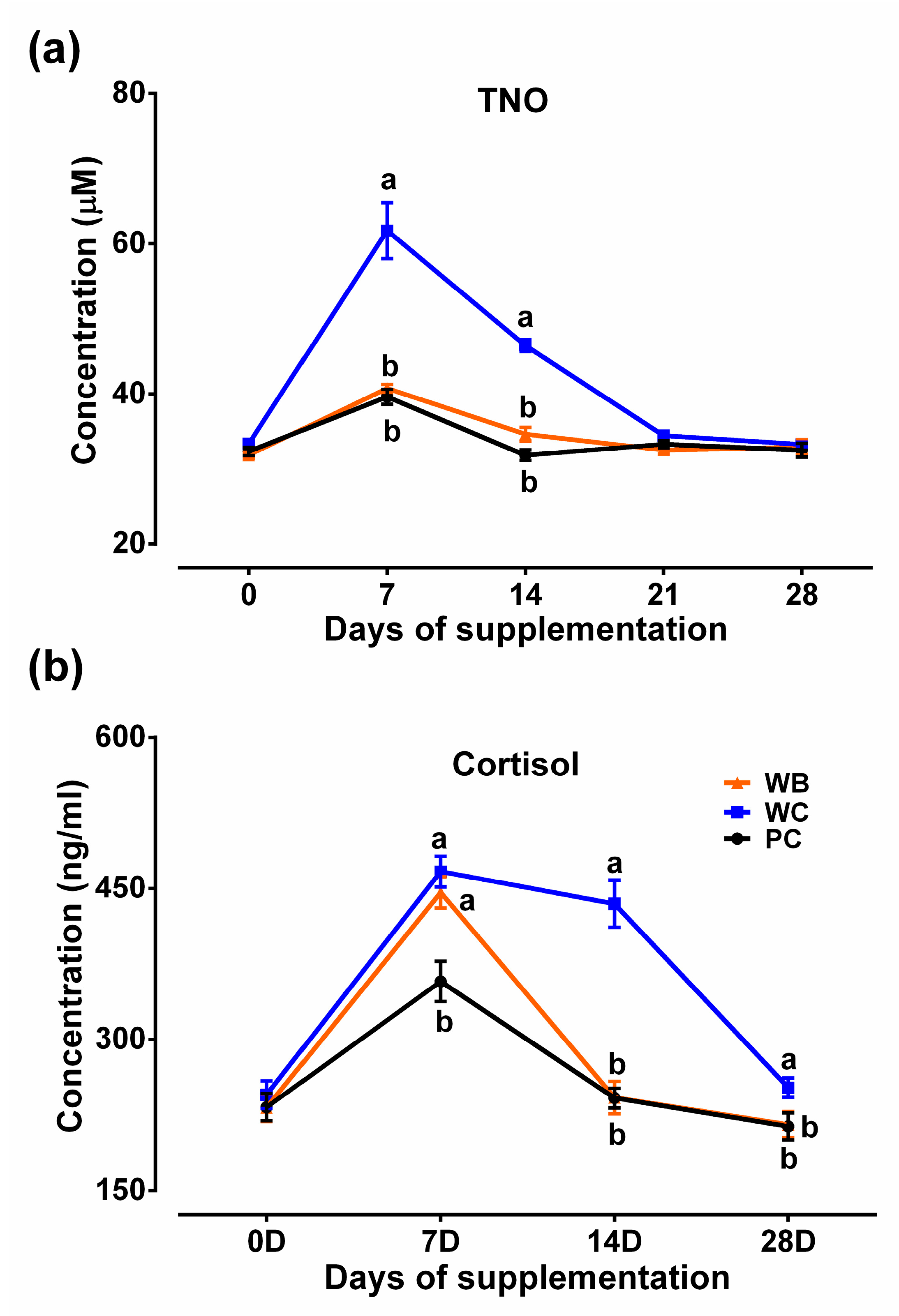
3.4. Evaluation of Stress Parameters
3.4.1. Total Serum Nitric Oxide Concentration
3.4.2. Serum Cortisol
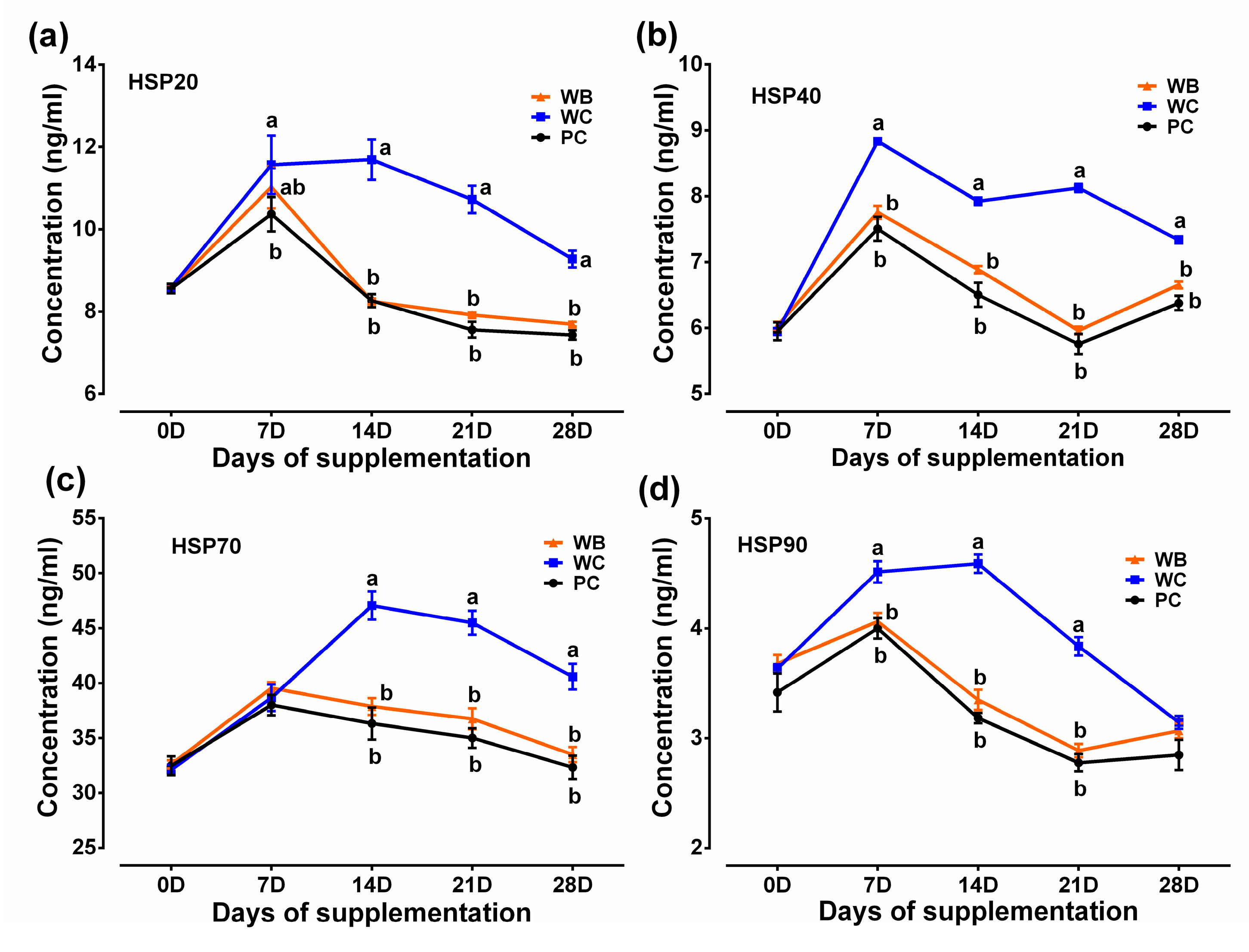
3.4.3. Serum Heat Shock Proteins (HSPs)
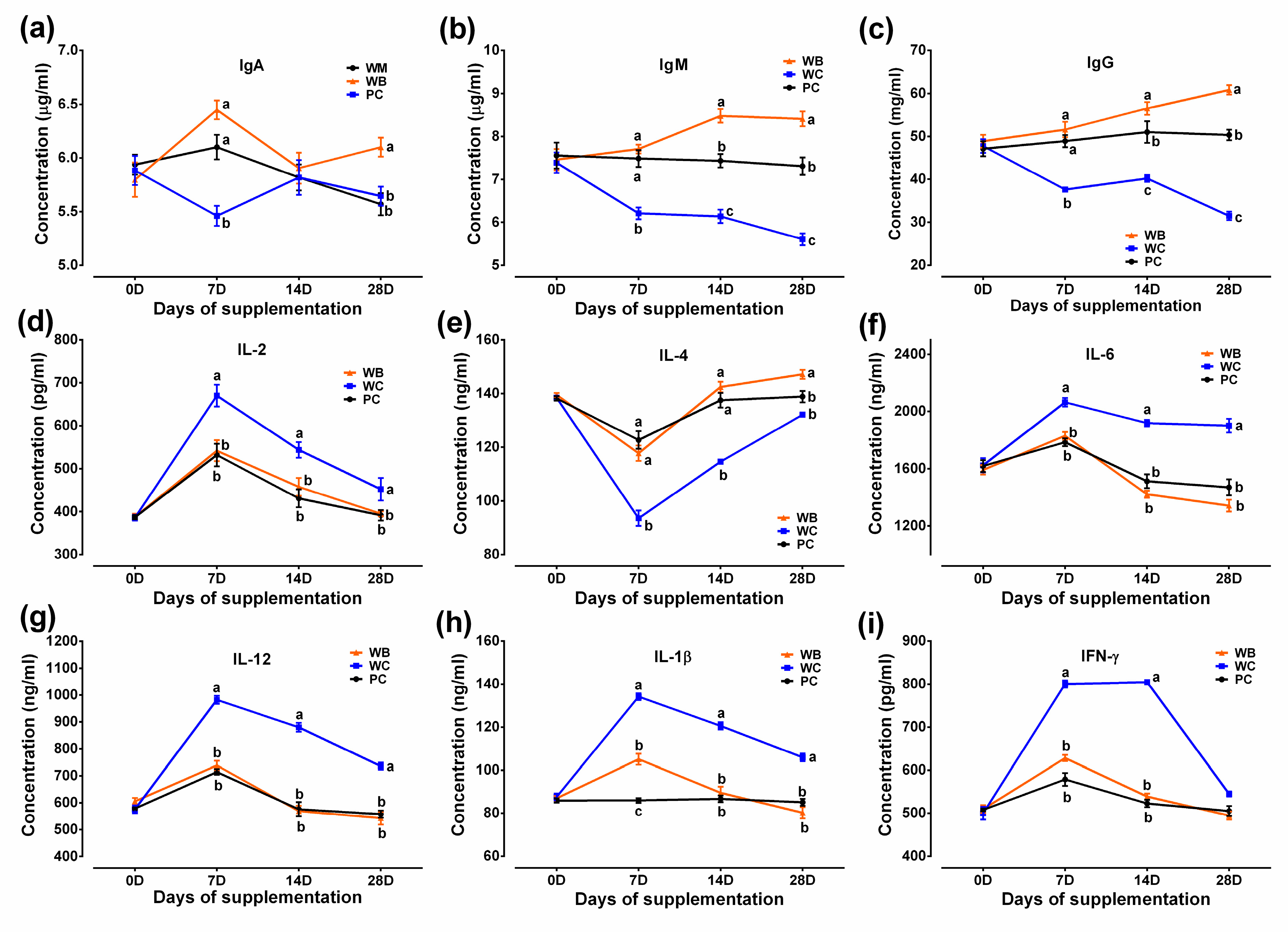
3.5. Immune Parameters of Serum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.Q.; Yin, F.G.; Zhu, C. Evaluation of probiotic bacteria for their effects on the growth performance and intestinal microbiota of newly-weaned pigs fed fermented high-moisture maize. Livest. Sci. 2012, 145, 79–86. [Google Scholar] [CrossRef]
- Jensen, P. Observations on the maternal behaviour of free-ranging domestic pigs. Appl. Anim. Behav. Sci. 1986, 16, 131–142. [Google Scholar] [CrossRef]
- Tang, X.; Xiong, K.; Fang, R.; Li, M. Weaning Stress and Intestinal Health of Piglets: A Review. Front. Immunol. 2022, 13, 1042778. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Heinonen, M.; Quesnel, H. High Physiological Demands in Intensively Raised Pigs: Impact on Health and Welfare. Animal 2010, 4, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.G.; Arthur, J.R.; Nicol, F.; Walker, M. Increased indices of lipid peroxidation in stress-susceptible pigs and effects of vitamin E. Res. Vet. Sci. 1989, 46, 226–230. [Google Scholar] [CrossRef]
- García, E.K.; de Souza, T.C.R.; Landín, G.M.; Barreyro, A.A.; Santos, M.G.B.; Soto, J.G.G. Microbial fermentation patterns, diarrhea incidence, and performance in weaned piglets fed a low protein diet supplemented with probiotics. Food Nutr. Sci. 2014, 5, 1776–1786. [Google Scholar] [CrossRef]
- Hao, Y.; Xing, M.; Gu, X. Research Progress on Oxidative Stress and Its Nutritional Regulation Strategies in Pigs. Animals 2021, 11, 1384. [Google Scholar] [CrossRef]
- Leibbrandt, V.D.; Ewan, R.C.; Speer, V.C.; Zimmerman, D.R. Effect of Weaning and Age at Weaning on Baby Pig Performance. J. Anim. Sci. 1975, 40, 1077–1080. [Google Scholar] [CrossRef]
- Faccin, J.E.G.; Laskoski, F.; Hernig, L.F.; Kummer, R.; Lima, G.F.R.; Orlando, U.A.D.; Goncalves, M.A.D.; Mellagi, A.P.G.; Ulguim, R.R.; Bortolozzo, F.P. Impact of Increasing Weaning Age on Pig Performance and Belly Nosing Prevalence in a Commercial Multisite Production System. J. Anim. Sci. 2020, 98, SKAA031. [Google Scholar] [CrossRef]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European Ban on Growth-Promoting Antibiotics and Emerging Consequences for Human and Animal Health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Li, C.; Zhao, M.; Wang, H.; Tang, Y. Withdrawal of Antibiotic Growth Promoters in China and Its Impact on the Foodborne Pathogen Campylobacter coli of Swine Origin. Front. Microbiol. 2022, 13, 1004725. [Google Scholar] [CrossRef] [PubMed]
- Kyung-hyo, D.; Jae-won, B.; Wan-kyu, L. Antimicrobial Resistance Profiles of Escherichia coli from Diarrheic Weaned Piglets after the Ban on Antibiotic Growth Promoters in Feed. Antibiotics 2020, 9, 755. [Google Scholar] [CrossRef] [PubMed]
- Bednorz, C.; Oelgeschläger, K.; Kinnemann, B.; Hartmann, S.; Neumann, K.; Pieper, R.; Bethe, A.; Semmler, T.; Tedin, K.; Schierack, P.; et al. The broader context of antibiotic resistance: Zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med. Microbiol. 2013, 303, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Slifierz, M.J.; Friendship, R.; Weese, J.S. Zinc oxide therapy increases prevalence and persistence of methicillin-resistant Staphylococcus aureus in pigs: A randomized controlled trial. Zoonoses Public Health 2015, 62, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Pejsak, Z.; Kaźmierczak, P.; Butkiewicz, A.F.; Wojciechowski, J.; Woźniakowski, G. Alternatives to Zinc Oxide in Pig Production. Pol. J. Vet. Sci. 2023, 26, 319–330. [Google Scholar] [CrossRef]
- Delcenserie, V.; Martel, D.; Lamoureux, M.; Amiot, J.; Boutin, Y.; Roy, D. Immunomodulatory effects of probiotics in the intestinal tract. Curr. Issues Mol. Biol. 2008, 10, 37–54. [Google Scholar]
- Schlee, M.; Harder, J.; Köten, B.; Stange, E.F.; Wehkamp, J.; Fellermann, K. Probiotic lactobacilli and VSL#3 induce enterocyte beta-defensin 2. Clin. Exp. Immunol. 2008, 151, 528–535. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Hong, H.A.; Le, H.D.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef]
- Weichselbaum, E. Probiotics and health: A review of the evidence. Nutr. Bull. 2009, 34, 340–373. [Google Scholar] [CrossRef]
- Andreoletti, O.; Lau Baggesen, D.; Bolton, D.; Butaye, P.; Cook, P.; Griffin, J.; Davies, R.; Fernández Escámez, P.S.; Griffin, J.; Hald, T.; et al. Scientific Opinion on the Maintenance of the List of QPS Biological Agents Intentionally Added to Food and Feed (2013 Update). EFSA J. 2013, 11, 3449. [Google Scholar] [CrossRef]
- Sun, T.; Miao, H.; Zhang, C.; Wang, Y.; Liu, S.; Jiao, P.; Li, W.; Li, Y.; Huang, Z. Effect of Dietary Bacillus coagulans on the Performance and Intestinal Microbiota of Weaned Piglets. Animal 2022, 16, 100561. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; McCartney, E.; Knox, A.; Francesch, M.; Oka, K.; Wada, K.; Ideno, M.; Uno, K.; Kozłowski, K.; Jankowski, J.; et al. Effects of the Butyric Acid-Producing Strain Clostridium butyricum MIYAIRI 588 on Broiler and Piglet Zootechnical Performance and Prevention of Necrotic Enteritis. Anim. Sci. J. 2018, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, G.; Cao, G.; Xu, Y.; Wang, Y.; Yang, C. Effects of Clostridium butyricum and Enterococcus faecalis on Growth Performance, Intestinal Structure, and Inflammation in Lipopolysaccharide-Challenged Weaned Piglets. J. Anim. Sci. 2019, 97, 4140. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Dun, Y.; Li, S.; Zhang, D.; Peng, N.; Zhao, S.; Liang, Y. Dietary Enterococcus faecalis LAB31 Improves Growth Performance, Reduces Diarrhea, and Increases Fecal Lactobacillus Number of Weaned Piglets. PLoS ONE 2015, 10, e0116635. [Google Scholar] [CrossRef]
- Tsukahara, T.; Inatomi, T.; Otomaru, K.; Amatatsu, M.; Romero-Pérez, G.A.; Inoue, R. Probiotic Supplementation Improves Reproductive Performance of Unvaccinated Farmed Sows Infected with Porcine Epidemic Diarrhea Virus. Anim. Sci. J. 2018, 89, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.; Tran, H.; Kim, I. Effects of probiotic supplementation in different nutrient density diets on growth performance, nutrient digestibility, blood profiles, fecal microflora and noxious gas emission in weaning pig. J. Sci. Food Agric. 2017, 97, 1335–1341. [Google Scholar] [CrossRef]
- Narayanappa, D. Randomized double blinded controlled trial to evaluate the efficacy and safety of Bifilac in patients with acute viral diarrhea. Indian J. Pediatr. 2008, 75, 709–713. [Google Scholar] [CrossRef]
- Poulsen, H.D. Zinc oxide for weanling piglets. Acta Agric. Scand. A-Anim. Sci. 1995, 45, 159–167. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of Supplementing Different Ratios of Laminarin and Fucoidan in the Diet of the Weanling Piglet on Performance, Nutrient Digestibility, and Fecal Scoring. J. Anim. Sci. 2012, 90, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, J.; Mahfuz, S.; Piao, X. Effects of Hydrolysable Tannins as Zinc Oxide Substitutes on Antioxidant Status, Immune Function, Intestinal Morphology, and Digestive Enzyme Activities in Weaned Piglets. Animals 2020, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- McNamara, J.R.; Cohn, J.S.; Wilson, P.W.F.; Schaefer, E.J. Calculated Values for Low-Density Lipoprotein Cholesterol in the Assessment of Lipid Abnormalities and Coronary Disease Risk. Clin. Chem. 1990, 36, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Marbut, M.M.; Majeed, B.M.; Rahim, S.M.; Yuusif, M.N. Estimation of malondialdehyde as oxidative factor & glutathione as early detectors of hypertensive pregnant women. Tikrit. Med. J. 2009, 15, 63–69. [Google Scholar]
- Chakraborty, M.; Gouroju, S.; Singh, P.; Thatkar, P.V.; Bagchi, D. Tracking Lipid Profile and Atherogenic Indices in the Prediabetics of Andaman Nicobar Islands: A Retrospective Hospital-Based Study. J. Fam. Med. Prim. Care 2019, 8, 1117. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kuroki, Y.; Oka, K.; Takahashi, M.; Rao, S.; Sukegawa, S.; Fujimura, T. Effects of Dietary Supplementation with Enterococcus faecium and Clostridium butyricum, Either Alone or in Combination, on Growth and Fecal Microbiota Composition of Post-Weaning Pigs at a Commercial Farm. Front. Vet. Sci. 2019, 6, 432044. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Indrakumar, S.; Kiarie, E.; Kim, I.H. Effects of a multi-strain Bacillus species–based direct-fed microbial on growth performance, nutrient digestibility, blood profile, and gut health in nursery pigs fed corn–soybean meal–based diets. J. Anim. Sci. 2015, 93, 4336–4342. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, M.; Zhao, L.; Ge, K.; Wang, Z.; Jun, L.; Ren, F. Growth performance and post-weaning diarrhea in piglets fed a diet supplemented with probiotic complexes. J. Microbiol. Biotechnol. 2018, 28, 1791–1799. [Google Scholar] [CrossRef]
- Bajagai, Y.S.; Klieve, A.V.; Bryden, W.L. Probiotics in Animal Nutrition—Production, Impact and Regulation; FAO Animal Production and Health Paper No. 179; Makkar, H.P.S., Ed.; FAO: Rome, Italy, 2016. [Google Scholar]
- Yi, R.; Pan, Y.; Long, X.; Tan, F.; Zhao, X. Enzyme Producing Activity of Probiotics and Preparation of Compound Enzyme. J. Chem. 2020, 2020, 9140281. [Google Scholar] [CrossRef]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Effects of Supplementation of Probiotics on the Performance, Nutrient Digestibility and Faecal Microflora in Growing-Finishing Pigs. Asian-Australasian J. Anim. Sci. 2011, 24, 655–661. [Google Scholar] [CrossRef]
- Méndez-Palacios, N.; Méndez-Mendoza, M.; Vázquez-Flores, F.; Castro-Colombres, J.G.; Ramírez-Bribiesca, J.E. Productive and Economic Parameters of Pigs Supplemented from Weaning to Finishing with Prebiotic and Probiotic Feed Additives. Anim. Sci. J. 2018, 89, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, J.N.; Laguna, J.S.; Millán, C.; Casabuena, O.; Gracia, M.I. Effects of a Bacillus-based probiotic and dietary energy content on the performance and nutrient digestibility of wean to finish pigs. Anim. Feed Sci. Technol. 2016, 221, 54–61. [Google Scholar] [CrossRef]
- Dodson, M.V.; Hausman, G.J.; Guan, L.; Du, M.; Rasmussen, T.P.; Poulos, S.P.; Mir, P.; Bergen, W.G.; Fernyhough, M.E.; McFarland, D.C.; et al. Lipid metabolism, adipocyte depot physiology and utilization of meat animals as experimental models for metabolic research. Int. J. Biol. Sci. 2010, 6, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.A. Plasma triglyceride and coronary heart disease. Arterioscler. Thromb. 1991, 11, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Busnelli, M.; Manzini, S.; Froio, A.; Vargiolu, A.; Cerrito, M.G.; Smolenski, R.T.; Giunti, M.; Cinti, A.; Zannoni, A.; Leone, B.E.; et al. Diet Induced Mild Hypercholesterolemia in Pigs: Local and Systemic Inflammation, Effects on Vascular Injury—Rescue by High-Dose Statin Treatment. PLoS ONE 2013, 8, 80588. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.T.; Ju, C.C.; Lin, J.; Wu, H.L.; Yen, H.T. Effects of probiotics and selenium combination on the immune and blood cholesterol concentration of pigs. J. Anim. Feed Sci. 2004, 13, 625–634. [Google Scholar] [CrossRef]
- Kim, D.; Min, Y.; Yang, J.; Heo, Y.; Kim, M.; Hur, C.G.; Lee, S.C.; Lee, H.K.; Song, K.D.; Heo, J.; et al. Multi-Probiotic Lactobacillus Supplementation Improves Liver Function and Reduces Cholesterol Levels in Jeju Native Pigs. Animals 2021, 11, 2309. [Google Scholar] [CrossRef]
- Novais, A.K.; Martel-Kennes, Y.; Roy, C.; Deschêne, K.; Beaulieu, S.; Bergeron, N.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Tissue-Specific Profiling Reveals Modulation of Cellular and Mitochondrial Oxidative Stress in Normal- and Low-Birthweight Piglets throughout the Peri-Weaning Period. Animal 2020, 14, 1014–1024. [Google Scholar] [CrossRef]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- Carvajal Carvajal, C. Reactive oxygen species: Training, function and oxidative stress. Med. Leg. Costa Rica 2019, 36, 91–100. [Google Scholar]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.R.; Harrison, D.G.; Bhatnagar, A. Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Cai, X.; Guo, Q.; Chen, X.; Zhu, S.; Xu, J. Effect of N-acetyl cysteine on enterocyte apoptosis and intracellular signalling pathways’ response to oxidative stress in weaned piglets. Br. J. Nutr. 2013, 110, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Q.; Zhuo, Y.; Fang, Z.; Che, L.; Xu, S.; Feng, B.; Lin, Y.; Jiang, X.; Zhao, X.; et al. Effects of Multi-Strain Probiotics and Perilla Frutescens Seed Extract Supplementation Alone or Combined on Growth Performance, Antioxidant Indices, and Intestinal Health of Weaned Piglets. Animals 2022, 12, 2246. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, Z.; Heng, J.; Song, H.; Tian, M.; Chen, F.; Guan, W. Combined Yeast Culture and Organic Selenium Supplementation during Late Gestation and Lactation Improve Preweaning Piglet Performance by Enhancing the Antioxidant Capacity and Milk Content in Nutrient-Restricted Sows. Anim. Nutr. 2020, 6, 160. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide Dismutases: Role in Redox Signaling, Vascular Function, and Diseases. Antioxid. Redox Signal. 2011, 15, 1583. [Google Scholar] [CrossRef]
- Wang, A.N.; Yi, X.W.; Yu, H.F.; Dong, B.; Qiao, S.Y. Free radical scavenging activity of Lactobacillus fermentum in vitro and its antioxidative effect on growing-finishing pigs. J. Appl. Microbiol. 2009, 107, 1140–1148. [Google Scholar] [CrossRef]
- Knauf, H.J.; Vogel, R.F.; Hammes, W.P. Cloning, sequence, and phenotypic expression of katA, which encodes the catalase of Lactobacillus sake LTH677. Appl. Environ. Microbiol. 1992, 58, 832–839. [Google Scholar] [CrossRef]
- Stecchini, M.L.; Del Torre, M.; Munari, M. Determination of peroxy radical-scavenging of lactic acid bacteria. Int. J. Food Microbiol. 2001, 64, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Arvans, D.L.; Vavricka, S.R.; Ren, H.; Musch, M.W.; Kang, L.; Rocha, F.G.; Lucioni, A.; Turner, J.R.; Alverdy, J.; Chang, E.B. Luminal Bacterial Flora Determines Physiological Expression of Intestinal Epithelial Cytoprotective Heat Shock Proteins 25 and 72. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G696–G704. [Google Scholar] [CrossRef]
- Sepponen, K.; Pösö, A.R. The inducible form of heat shock protein 70 in the serum, colon and small intestine of the pig: Comparison to conventional stress markers. Vet. J. 2006, 171, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Yang, J.; Qi, Z.; Wu, H.; Wang, B.; Zou, F.; Mei, H.; Liu, J.; Wang, W.; Liu, Q. Heat Shock Proteins: Biological Functions, Pathological Roles, and Therapeutic Opportunities. MedComm 2022, 3, e161. [Google Scholar] [CrossRef] [PubMed]
- David, J.C.; Grongnet, J.F.; Lalles, J.P. Weaning Affects the Expression of Heat Shock Proteins in Different Regions of the Gastrointestinal Tract of Piglets. J. Nutr. 2002, 132, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Ren, F.; Chen, X.; Lv, C.; Pan, C.; Ye, G.; Shi, J.; Shi, X.; Zhou, H.; Shituleni, S.A.; et al. Effects of selenium-enriched probiotics on heat shock protein mRNA levels in piglet under heat stress conditions. J. Agric. Food Chem. 2013, 61, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Kick, A.R.; Tompkins, M.B.; Flowers, W.L.; Whisnant, C.S.; Almond, G.W. Effects of Stress Associated with Weaning on the Adaptive Immune System in Pigs. J. Anim. Sci. 2012, 90, 649–656. [Google Scholar] [CrossRef]
- Hinds, J.A.; Sanchez, E.R. The Role of the Hypothalamus–Pituitary–Adrenal (HPA) Axis in Test-Induced Anxiety: Assessments, Physiological Responses, and Molecular Details. Stress 2022, 2, 146–155. [Google Scholar] [CrossRef]
- Martínez-Miró, S.; Tecles, F.; Ramón, M.; Escribano, D.; Hernández, F.; Madrid, J.; Orengo, J.; Martínez-Subiela, S.; Manteca, X.; Cerón, J.J. Causes, Consequences and Biomarkers of Stress in Swine: An Update. BMC Vet. Res. 2016, 12, 171. [Google Scholar] [CrossRef] [PubMed]
- Van Erp-Van Der Kooij, E.; Kuijpers, A.H.; Van Eerdenburg, F.J.C.M.; Dieleman, S.J.; Blankenstein, D.M.; Tielen, M.J.M. Individual behavioural characteristics in pigs—Influences of group composition but no differences in cortisol responses. Physiol. Behav. 2003, 78, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Burdick Sanchez, N.C.; Carroll, J.A.; Broadway, P.R.; Bass, B.E.; Frank, J.W. Supplementation of a Lactobacillus acidophilus Fermentation Product Can Attenuate the Acute Phase Response Following a Lipopolysaccharide Challenge in Weaned Pigs. Animal 2019, 13, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.N.; Cai, C.J.; Zeng, X.F.; Zhang, F.R.; Zhang, G.L.; Thacker, P.A.; Wang, J.J.; Qiao, S.Y. Dietary supplementation with Lactobacillus fermentum I5007 improves the anti-oxidative activity of weanling piglets challenged with diquat. J. Appl. Microbiol. 2013, 114, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Simon, M.V.; Lee, J.H.; Ustunol, Z.; Pestka, J.J. Ingestion of yogurt containing Lactobacillus acidophilus and Bifidobacterium to potentiate immunoglobulin A responses to cholera toxin in mice. J. Dairy Sci. 1999, 82, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Travers, P.; Walport, M.; Shlomchik, M. The humoral immune response, immunobiology. In The Immune System in Health and Disease, 5th ed.; Austin, P., Lawrence, E., Eds.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Dong, X.; Zhang, N.; Zhou, M.; Tu, Y.; Deng, K.; Diao, Q. Effects of dietary probiotics on growth performance, Faecal microbiota and serum profiles in weaned piglets. Anim. Prod. Sci. 2014, 54, 616–621. [Google Scholar] [CrossRef]
- Dlamini, Z.C.; Langa, R.L.S.; Aiyegoro, O.A.; Okoh, A.I. Effects of probiotics on growth performance, blood parameters, and antibody stimulation in piglets. S. Afr. J. Anim. Sci. 2017, 47, 766–775. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
- Johnson, R.W. Inhibition of growth by pro-inflammatory cytokines: An integrated view. J. Anim. Sci. 1997, 75, 1244–1255. [Google Scholar] [CrossRef]
- De Groot, N.; Fariñas, F.; Cabrera-Gómez, C.G.; Pallares, F.J.; Ramis, G. Weaning Causes a Prolonged but Transient Change in Immune Gene Expression in the Intestine of Piglets. J. Anim. Sci. 2021, 99, skab065. [Google Scholar] [CrossRef] [PubMed]
- Bomba, L.; Minuti, A.; Moisá, S.J.; Trevisi, E.; Eufemi, E.; Lizier, M.; Chegdani, F.; Lucchini, F.; Rzepus, M.; Prandini, A.; et al. Gut Response Induced by Weaning in Piglet Features Marked Changes in Immune and Inflammatory Response. Funct. Integr. Genom. 2014, 14, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Tao, F.; Hu, Y.; Li, Z.; Zhang, Y.; Deng, B.; Zhan, X. Positive effects of a Clostridium butyricum-based compound probiotic on growth performance, immune responses, intestinal morphology, hypothalamic neurotransmitters, and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2926–2934. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, E.; Jarosz, Ł.; Grądzki, Z. Effect of multi-microbial probiotic formulation bokashi on pro- and anti-inflammatory cytokines profile in the serum, colostrum and milk of sows, and in a culture of polymorphonuclear cells isolated from colostrum. Probiotics Antimicrob. Proteins 2019, 11, 220–232. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Composition | Strain Number | GenBank Accession Details | Deposition Details | Content per Gram of Product |
|---|---|---|---|---|
| Enterococcus faecalis | T-110 | AB687552: 16S DNA CP006030: Complete genome CP006031: Complete plasmid | 8936 * | 3 × 107 CFU/g |
| Clostridium butyricum | TOA | AB687551: 16S DNA CP014704: Chromosome 1 CP014705: Chromosome 2 CP014706: Plasmid | 8935 * | 2 × 106 CFU/g |
| Bacillus mesentericus | TOA | AB687550:16S DNA CP005997: Complete genome | 8934 * | 1 × 106 CFU/g |
| Bacillus coagulans | SNZ 1969 | KC146407: 16S DNA | MTCC 5724 | 5 × 107 CFU/g |
| Ingredients | Percentage |
|---|---|
| Maize | 50.00 |
| Wheat bran | 15.00 |
| Soybean meal | 29.00 |
| Vitamin and trace min. mix | 2.50 |
| MCP | 1.00 |
| Salt | 1.00 |
| CaCO3 | 1.28 |
| DL-Methionine | 0.22 |
| Chemical formula of basal diet (on dry matter basis) | |
| Dry matter (%) | 91.46 |
| Crude Protein (CP) (%) | 20.72 |
| Crude fiber (%) | 3.490 |
| Ether extract (%) | 4.827 |
| Calcium (%) | 0.80 |
| Total phosphorus (%) | 0.63 |
| Lysine (%) | 0.80 |
| Methionine and cystine (%) | 0.70 |
| Metabolizable energy (ME) (kcal/kg) | 3382.31 |
| Parameters | WC | WB | PC | p-Value |
|---|---|---|---|---|
| Body Weight/kg | ||||
| 0 days | 8.84 ± 0.03 | 8.79 ± 0.04 | 8.75 ± 0.04 | 0.204 |
| 7 days | 9.87 b ± 0.09 | 10.11 a ± 0.07 | 10.38 a ± 0.05 | <0.001 |
| 14 days | 11.25 b ± 0.04 | 11.79 a ± 0.03 | 11.78 a ± 0.04 | <0.001 |
| 21 days | 12.58 b ± 0.05 | 13.63 a ± 0.10 | 13.71 a ± 0.11 | <0.001 |
| 28 days | 13.80 b ± 0.06 | 14.88 a ± 0.12 | 14.97 a ± 0.13 | <0.001 |
| ADG/g | ||||
| 0–14 days | 171.90 b ± 3.09 | 214.17 a ± 4.08 | 216.19 a ± 4.61 | <0.001 |
| 14–28 days | 182.26 b ± 5.11 | 220.48 a ± 8.22 | 228.10 a± 9.15 | <0.001 |
| 0–28 days | 177.08 b ± 2.08 | 217.32 a ± 4.49 | 222.14 a ± 4.84 | <0.001 |
| ADFI/g | ||||
| 0–14 days | 417.73 b ± 7.51 | 454.03 ab ± 8.66 | 459.20 a ± 9.83 | 0.003 |
| 14–28 days | 466.59 ± 13.08 | 485.05 ± 18.08 | 499.20 ± 20.04 | 0.413 |
| 0–28 days | 442.16 ± 5.33 | 469.54 ± 9.81 | 479.20 ± 10.62 | 0.116 |
| F:G | ||||
| 0–14 days | 2.43 a ± 0.01 | 2.11 b ± 0.01 | 2.12 b ± 0.004 | <0.001 |
| 14–28 days | 2.57 a ± 0.01 | 2.20 b ± 0.02 | 2.18 b ± 0.003 | <0.001 |
| 0–28 days | 2.49 a ± 0.02 | 2.16 b ± 0.04 | 2.16 b ± 0.003 | <0.001 |
| Diarrhea rate (%) | ||||
| 0–28 days | 11.67 a ± 0.38 | 4.67 b ± 0.31 | 4.58 b ± 0.26 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, G.; Mondal, S.; Bhattacharya, D.; Ponraj, P.; Sawhney, S.; Bala, P.; Chakraborty, D.; Sunder, J.; De, A.K. Effect of a Multi-Strain Probiotic on Growth Performance, Lipid Panel, Antioxidant Profile, and Immune Response in Andaman Local Piglets at Weaning. Fermentation 2023, 9, 970. https://doi.org/10.3390/fermentation9110970
Sarkar G, Mondal S, Bhattacharya D, Ponraj P, Sawhney S, Bala P, Chakraborty D, Sunder J, De AK. Effect of a Multi-Strain Probiotic on Growth Performance, Lipid Panel, Antioxidant Profile, and Immune Response in Andaman Local Piglets at Weaning. Fermentation. 2023; 9(11):970. https://doi.org/10.3390/fermentation9110970
Chicago/Turabian StyleSarkar, Gopal, Samiran Mondal, Debasis Bhattacharya, Perumal Ponraj, Sneha Sawhney, Prokasananda Bala, Dibyendu Chakraborty, Jai Sunder, and Arun Kumar De. 2023. "Effect of a Multi-Strain Probiotic on Growth Performance, Lipid Panel, Antioxidant Profile, and Immune Response in Andaman Local Piglets at Weaning" Fermentation 9, no. 11: 970. https://doi.org/10.3390/fermentation9110970
APA StyleSarkar, G., Mondal, S., Bhattacharya, D., Ponraj, P., Sawhney, S., Bala, P., Chakraborty, D., Sunder, J., & De, A. K. (2023). Effect of a Multi-Strain Probiotic on Growth Performance, Lipid Panel, Antioxidant Profile, and Immune Response in Andaman Local Piglets at Weaning. Fermentation, 9(11), 970. https://doi.org/10.3390/fermentation9110970