
Monascus Yellow Pigment Production by Coupled Immobilized-Cell Fermentation and Extractive Fermentation in Nonionic Surfactant Micelle Aqueous Solution

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
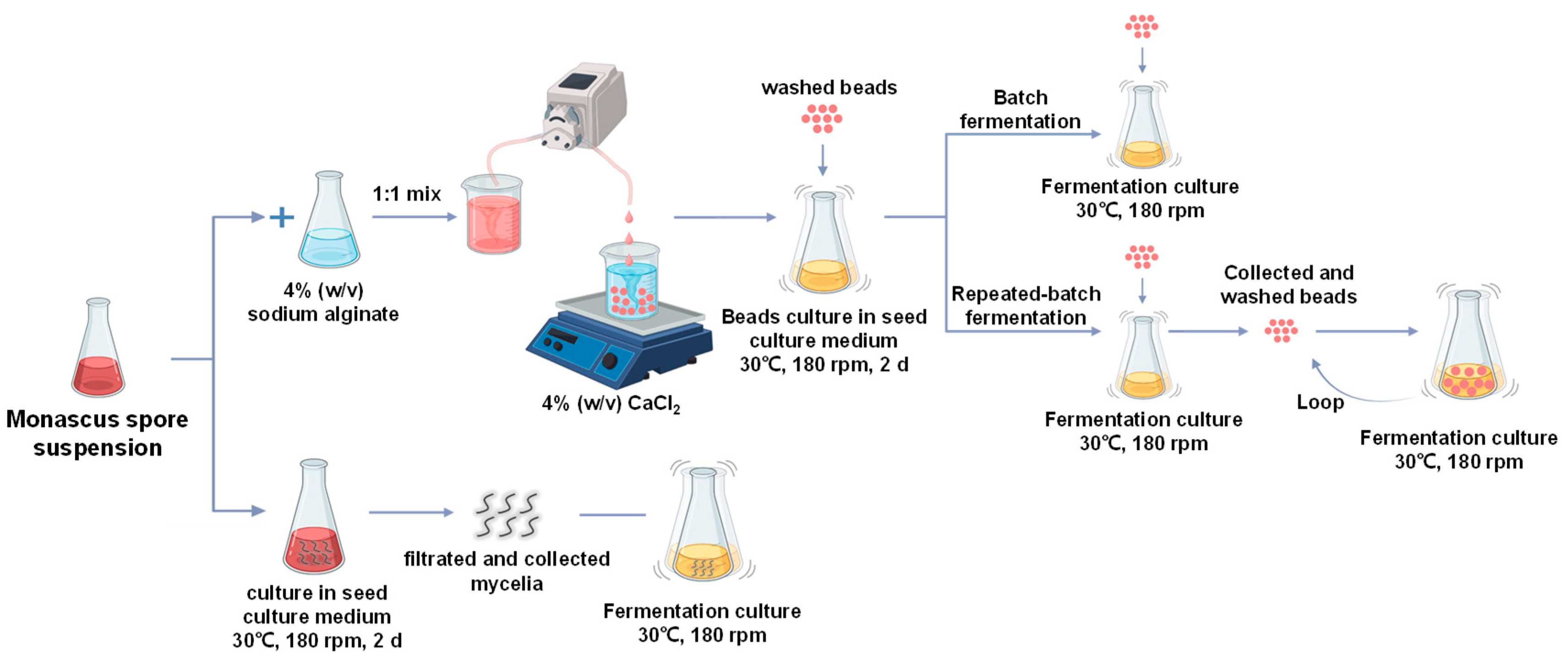
2.1. Microorganism and Cultivation Media
2.2. Fermentation
2.2.1. Free-Cell Fermentation
2.2.2. Immobilized-Cell Fermentation
2.2.3. Repeated-Batch Extractive Fermentation with Immobilized Cells
2.3. Analysis Methods
2.4. Statistical Analysis
3. Results and Discussion
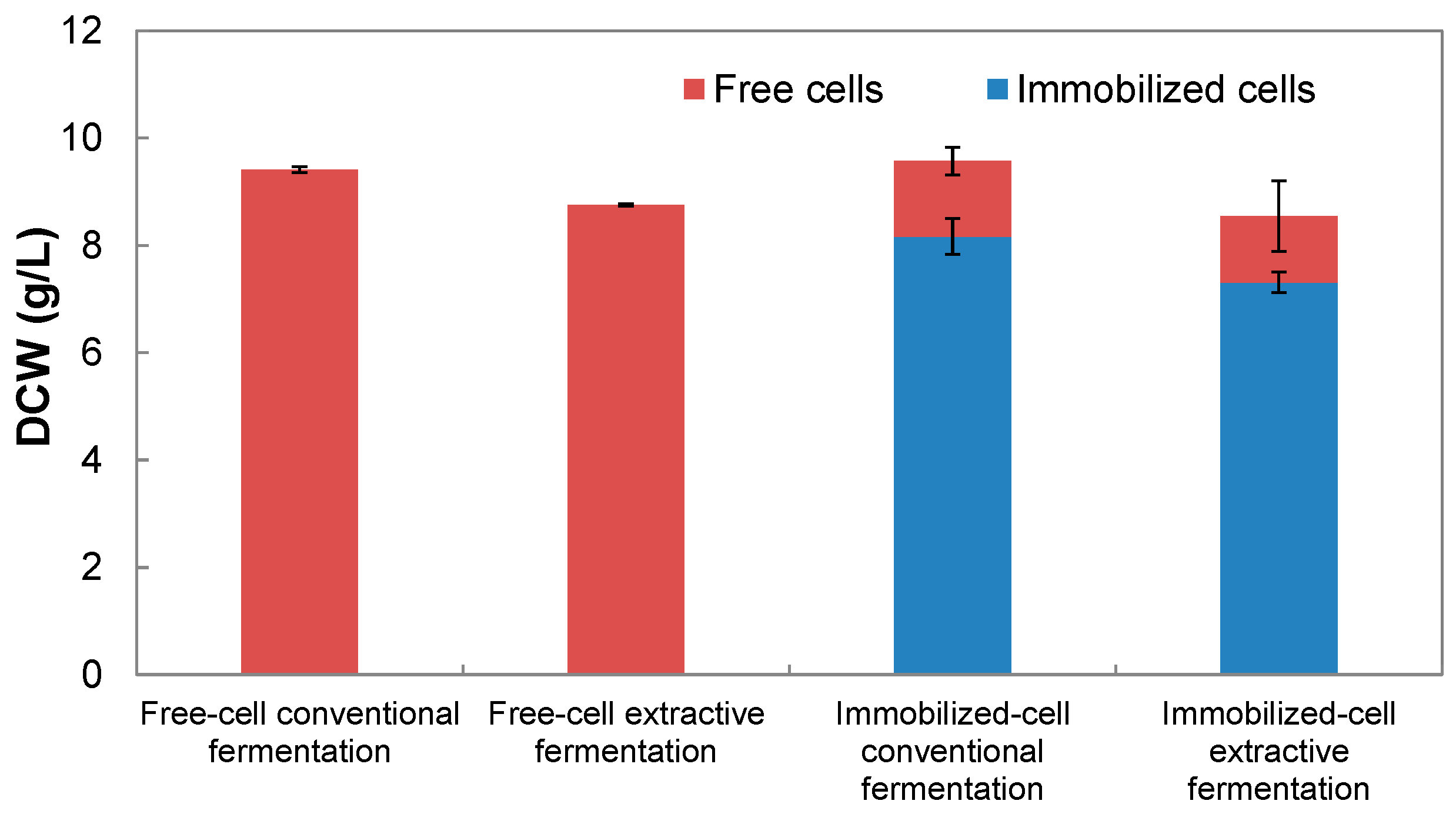
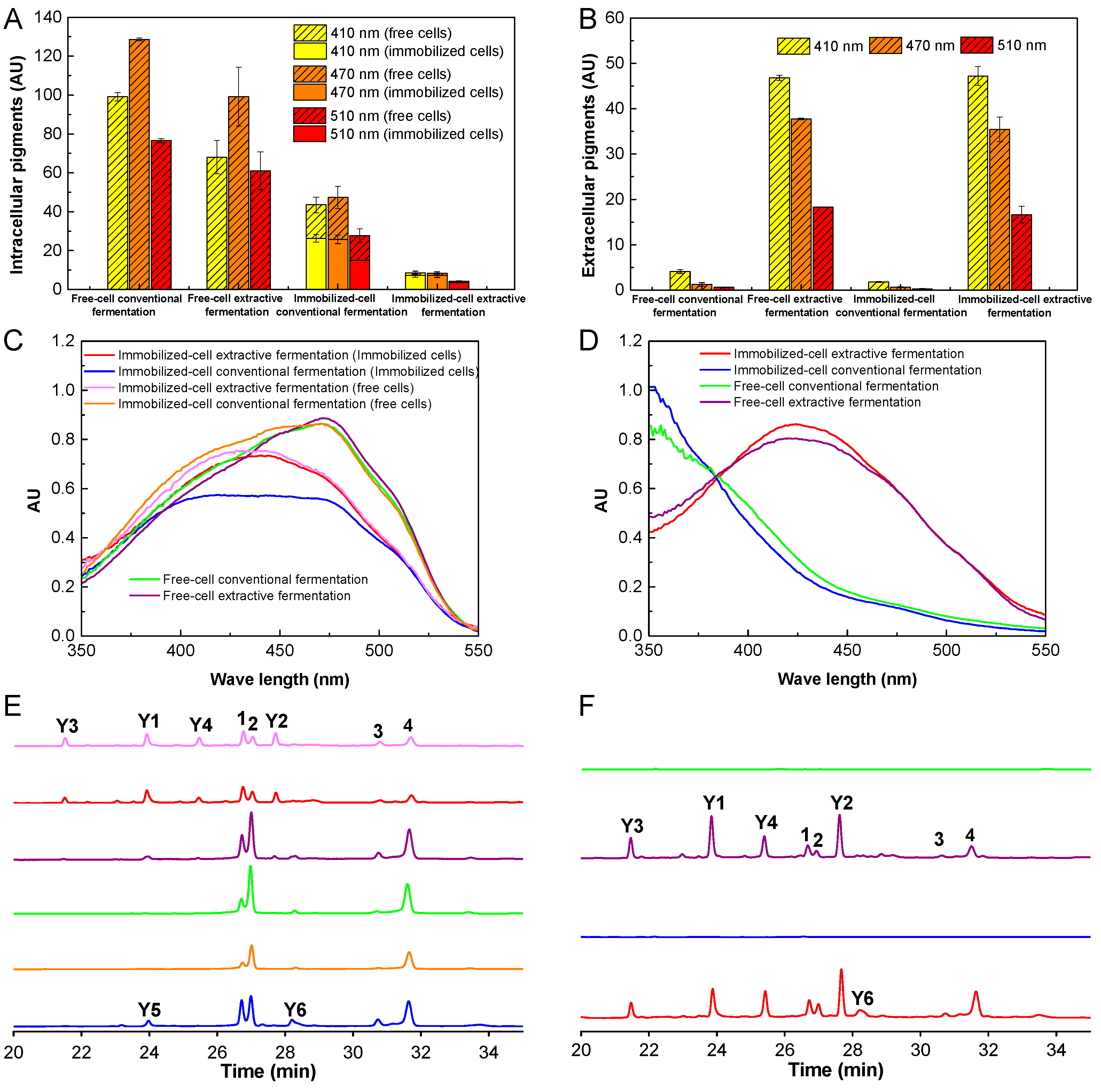
3.1. Effects of Cell Immobilization and Triton X-100 on Cell Growth, Pigment Production and Pigment Composition
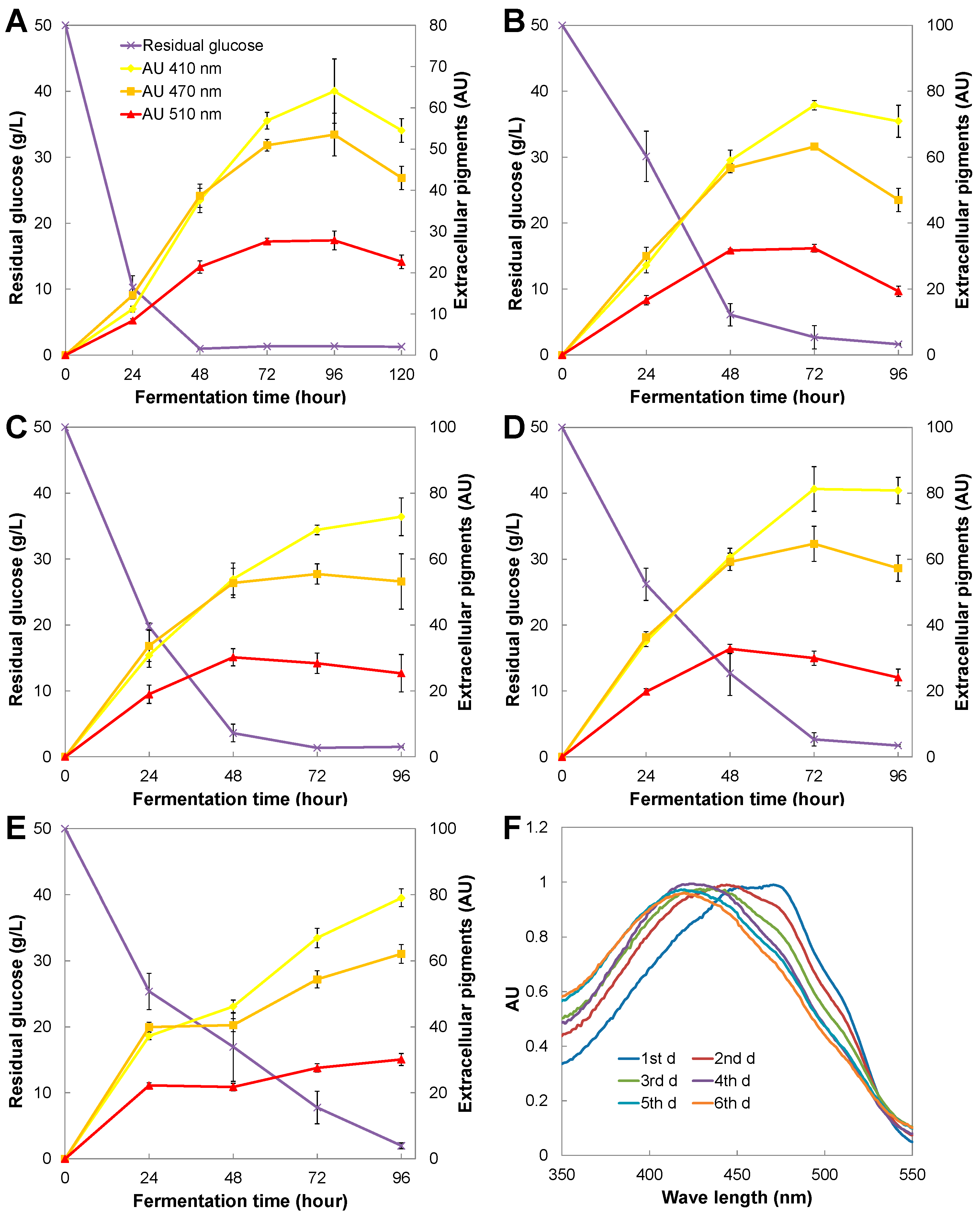
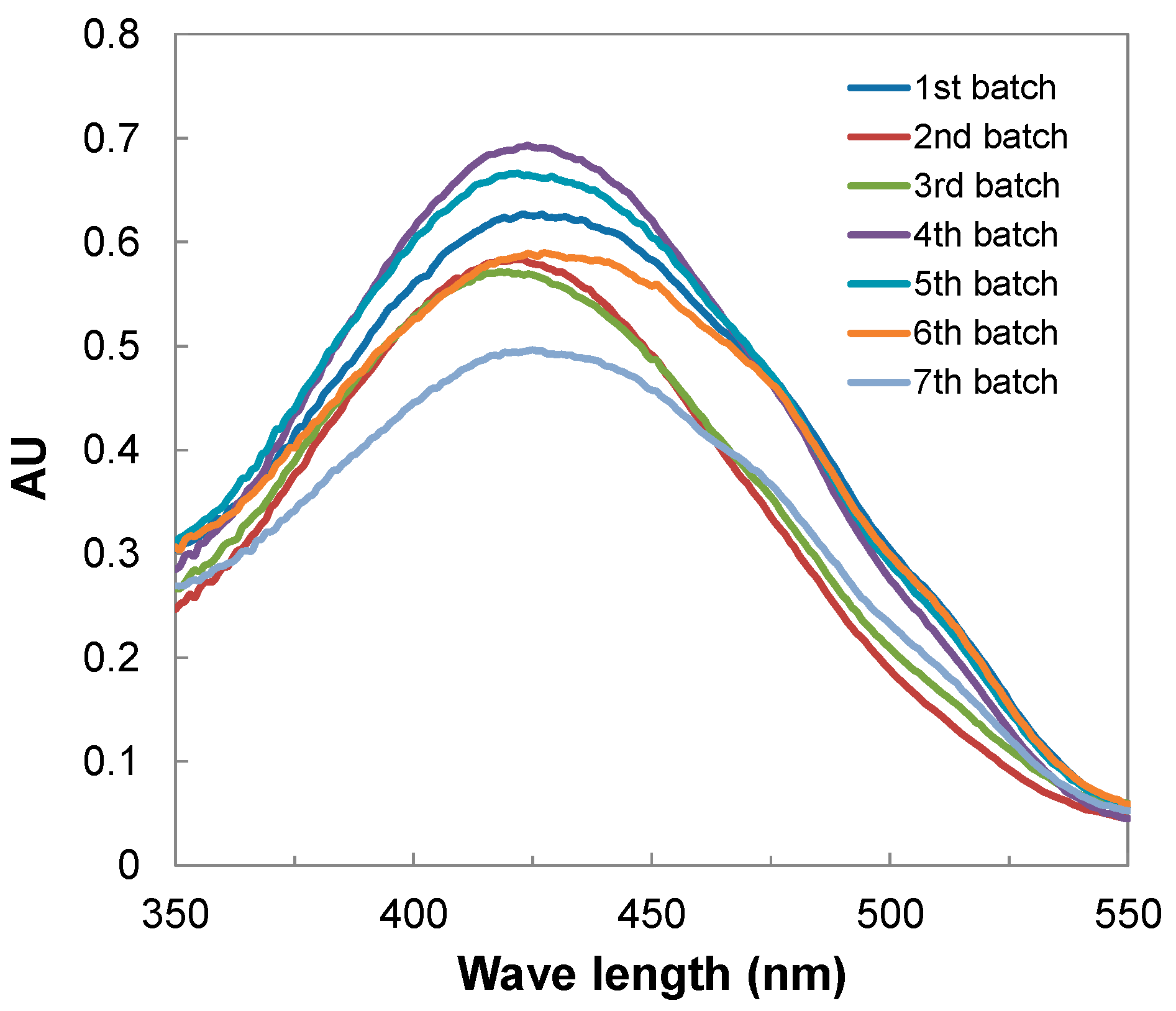
3.2. Yellow Pigment Production in Repeated-Batch Extractive Fermentation with Immobilized Cells
3.3. Cell Growth and Intracellular Pigments Metabolism in Repeated-Batch Extractive Fermentation with Immobilized Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, W.; He, Y.; Zhou, Y.; Shao, Y.; Feng, Y.; Li, M.; Chen, F. Edible filamentous fungi from the species Monascus: Early traditional fermentations, modern molecular biology, and future genomics. Compr. Rev. Food. Sci. Food Saf. 2015, 14, 555–567. [Google Scholar] [CrossRef]
- Feng, Y.; Shao, Y.; Chen, F. Monascus pigments. Appl. Microbiol. Biotechnol. 2012, 96, 1421–1440. [Google Scholar] [CrossRef] [PubMed]
- Adin, S.N.; Gupta, I.; Panda, B.P.; Mujeeb, M. Monascin and ankaflavin—Biosynthesis from Monascus purpureus, production methods, pharmacological properties: A review. Biotechnol. Appl. Biochem. 2022, 1–11. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jia, M.; Li, W.; Deng, J.; Ren, J.; Luo, F.; Bai, J.; Liu, J. Toward improvements for enhancement the productivity and color value of Monascus pigments: A critical review with recent updates. Crit. Rev. Food Sci. Nutr. 2021, 62, 7139–7153. [Google Scholar] [CrossRef] [PubMed]
- Fabre, C.E.; Santerre, A.L.; Loret, M.O.; Baberian, R.; Pareilleux, A.; Goma, G.; Blanc, P.J. Production and food applications of the red pigments of Monascus ruber. J. Food Sci. 1993, 58, 1099–1102. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Metabolic regulation and overproduction of primary metabolites. Microb. Biotechnol. 2008, 1, 283–319. [Google Scholar] [CrossRef]
- Stark, D.; von Stockar, U. In situ product removal (ISPR) in whole cell biotechnology during the last twenty years. Adv. Biochem. Eng. Biotechnol. 2003, 80, 149–175. [Google Scholar]
- Santos, A.G.; de Albuquerque, T.L.; Ribeiro, B.D.; Coelho, M.A.Z. In situ product recovery techniques aiming to obtain biotechnological products: A glance to current knowledge. Biotechnol. Appl. Biochem. 2021, 68, 1044–1057. [Google Scholar] [CrossRef]
- Van Hecke, W.; Kaur, G.; De Wever, H. Advances in in-situ product recovery (ISPR) in whole cell biotechnology during the last decade. Biotechnol. Adv. 2014, 32, 1245–1255. [Google Scholar] [CrossRef]
- Lam, K.S.; Gustavson, D.R.; Veitch, J.A.; Forenza, S. The effect of cerulenin on the production of esperamicin A1 by Actinomadura verrucosospora. J. Ind. Microbiol. 1993, 12, 99–102. [Google Scholar] [CrossRef]
- Lee, H.B.; Kim, C.J.; Lee, J.C.; Kim, Y.B.; Park, H.R.; Park, D.J. Improved production of teicoplanin using adsorbent resin in fermentations. Lett. Appl. Microbiol. 2003, 37, 196–200. [Google Scholar] [CrossRef]
- Bing, L.; Hui, J.; Cheng, Y.Q.; Zhang, X. Extractive fermentation for enhanced production of thailandepsin A from Burkholderia thailandensis E264 using polyaromatic adsorbent resin Diaion HP-20. J. Ind. Microbiol. Biotechnol. 2012, 39, 767–776. [Google Scholar]
- Hua, D.; Lin, S.; Li, Y.; Chen, H.; Zhang, Z.; Du, Y.; Zhang, X.; Xu, P. Enhanced 2-phenylethanol production from L-phenylalanine via in situ product adsorption. Biocatal. Biotransform. 2010, 28, 259–266. [Google Scholar] [CrossRef]
- Outram, V.; Lalander, C.A.; Lee, J.G.; Davies, E.T.; Harvey, A.P. Applied in situ product recovery in ABE fermentation. Biotechnol. Prog. 2017, 33, 563–579. [Google Scholar] [CrossRef]
- Kleinegris, D.M.; Janssen, M.; Brandenburg, W.A.; Wijffels, R.H. Two-phase systems: Potential for in situ extraction of microalgal products. Biotechnol. Adv. 2011, 29, 502–507. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, B.; Lu, L.; Huang, Y.; Xu, G. Enhanced production of pigments by addition of surfactants in submerged fermentation of Monascus purpureus H1102. J. Sci. Food Agric. 2013, 93, 3339–3344. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Perstraction of intracellular pigments by submerged cultivation of Monascus in nonionic surfactant micelle aqueous solution. Appl. Microbiol. Biotechnol. 2012, 94, 81–89. [Google Scholar] [CrossRef]
- Kang, B.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Solubilization capacity of nonionic surfactant micelles exhibiting strong influence on export of intracellular pigments in Monascus fermentation. Microb. Biotechnol. 2013, 6, 540–550. [Google Scholar] [CrossRef]
- Xiong, X.; Zhang, X.; Wu, Z.; Wang, Z. Accumulation of yellow Monascus pigments by extractive fermentation in nonionic surfactant micelle aqueous solution. Appl. Microbiol. Biotechnol. 2014, 99, 1173–1180. [Google Scholar] [CrossRef]
- Shi, K.; Tang, R.; Huang, T.; Wang, L.; Wu, Z. Pigment fingerprint profile during extractive fermentation with Monascus anka GIM 3.592. Bmc Biotechnol. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Gao, H.; Lu, J.; Jiang, Y.; Fang, Y.; Tang, Y.; Yu, Z.; Zhang, W.; Xin, F.; Jiang, M. Material-mediated cell immobilization technology in the biological fermentation proces. Biofuels Bioprod. Biorefining 2021, 15, 1160–1173. [Google Scholar] [CrossRef]
- Li, C.; Wang, C.; Zhang, H.; Li, T. Study on water-soluble pigment from fermentation of immobilized Monuscus. China Food Addit. 2008, 2, 106–108. [Google Scholar]
- Liu, J.; Ren, Y.; Yao, S. Repeated-batch Cultivation of Encapsulated Monascus purpureus by Polyelectrolyte Complex for Natural Pigment Production. Chin. J. Chem. Eng. 2010, 18, 1013–1017. [Google Scholar] [CrossRef]
- Yi, X.; Hou, H.; Li, T.; Shao, W. Optimization of fermentation condition for the immobilization of Monascus purpureus cell with RSM. China Brew. 2008, 9, 80–83. [Google Scholar]
- Evans, P.J.; Wang, H.Y. Pigment production from immobilized Monascus sp. utilizing polymeric resin adsorption. Appl. Environ. Microbiol. 1984, 47, 1323–1326. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S. Study of production of Monascus pigment by PVA immobilized Monascus cell. J. Yantai Univ. (Nat. Sci. Eng.) 1998, 11, 306–309. [Google Scholar]
- Fenice, M.; Federici, F.; Selbmann, L.; Petruccioli, M. Repeated-batch production of pigments by immobilised Monascus purpureus. J. Biotechnol. 2000, 80, 271–276. [Google Scholar] [CrossRef]
- Babitha, S.; Soccol, C.R.; Pandey, A. Effect of stress on growth, pigment production and morphology of Monascus sp. in solid cultures. J. Basic Microbiol. 2007, 47, 118–126. [Google Scholar] [CrossRef]
- Shi, K.; Chen, G.; Pistolozzi, M.; Xia, F.; Wu, Z. Improved analysis of Monascus pigments based on their pH-sensitive Uv-Vis absorption and reactivity properties. Food Addit. Contam. Part A 2016, 33, 1396–1401. [Google Scholar] [CrossRef]
- Pashova, S.; Slokoska, L.; Sheremetska, P.; Krumova, E.; Vasileva, L.; Angelova, M. Physiological aspects of immobilised Aspergillus niger cells producing polymethylgalacturonase. Process Biochem. 1999, 35, 15–19. [Google Scholar] [CrossRef]
- Zhu, Y. Immobilized cell fermentation for production of chemicals and fuels. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.-T., Ed.; Elsevier: Amsterdam, Netherlands, 2007; pp. 373–396. [Google Scholar]
- Bucke, C.; Brown, D.E. Immobilized cells [and discussion]. Philos. Trans. R. Soc. Lond. Ser. B 1983, 300, 369–389. [Google Scholar]
- Kang, B.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Effect of pH and nonionic surfactant on profile of intracellular and extracellular Monascus pigments. Process Biochem. 2013, 48, 759. [Google Scholar] [CrossRef]
- Shi, K.; Song, D.; Chen, G.; Pistolozzi, M.; Wu, Z.; Quan, L. Controlling composition and color characteristics of Monascus pigments by pH and nitrogen sources in submerged fermentation. J. Biosci. Bioeng. 2015, 120, 145–154. [Google Scholar] [CrossRef]
- Chen, G.; Bei, Q.; Huang, T.; Wu, Z. Tracking of pigment accumulation and secretion in extractive fermentation of Monascus anka GIM 3.592. Microb. Cell. Fact. 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Export of intracellular Monascus pigments by two-stage microbial fermentation in nonionic surfactant micelle aqueous solution. J. Biotechnol. 2012, 162, 202–209. [Google Scholar] [CrossRef]
- Chen, G.; Bei, Q.; Shi, K.; Tian, X.; Wu, Z. Saturation effect and transmembrane conversion of Monascus pigment in nonionic surfactant aqueous solution. Amb Express 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.Q.; Xin, Y.W.; Guo, Y.H. Study on the fingerprint profile of Monascus products with HPLC–FD, PAD and MS. Food Chem. 2009, 113, 705–711. [Google Scholar] [CrossRef]
- Teng, S.S.; Feldheim, W. Analysis of anka pigments by liquid chromatography with diode array detection and tandem mass spectrometry. Chromatographia 1998, 47, 529–536. [Google Scholar] [CrossRef]
- de Alteriis, E.; Zueco, J.; Rico, H.; Sentandreu, R.; Parascandola, P. Electrophoretic mobility of external invertase from free and gel-immobilized yeast cells. Res. Microbiol. 1995, 146, 217–225. [Google Scholar] [CrossRef]
- Chen, W.; Feng, Y.; Molnár, I.; Chen, F. Nature and nurture: Confluence of pathway determinism with metabolic and chemical serendipity diversifies Monascus azaphilone pigments. Nat. Prod. Rep. 2019, 36, 561–572. [Google Scholar] [CrossRef]
- Junter, G.; Coquet, L.; Vilain, S.; Jouenne, T. Immobilized-cell physiology: Current data and the potentialities of proteomics. Enzyme Microb. Technol. 2002, 31, 201–212. [Google Scholar] [CrossRef]
- Liu, J.; Guo, T.; Luo, Y.; Chai, X.; Wu, J.; Zhao, W.; Jiao, P.; Luo, F.; Lin, Q. Enhancement of Monascus pigment productivity via a simultaneous fermentation process and separation system using immobilized-cell fermentation. Bioresour. Technol. 2019, 272, 552–560. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Pigments | HPLC Retention Time (min) | Characteristic Absorption Peaks (nm) | Molecular Weight a |
|---|---|---|---|---|
| 1 | Monascin | 26.68 | 230, 291, 388 | 358 |
| 2 | Rubropunctatin | 26.94 | 214, 247, 286, 471 | 354 |
| 3 | Ankaflavin | 30.62 | 230, 293, 388 | 386 |
| 4 | Monascorubrin | 31.51 | 214, 274, 285, 471 | 382 |
| Y1 | Not reported | 23.84 | 219, 310, 430 | 480 |
| Y2 | Not reported | 27.62 | 219, 310, 430 | 508 |
| Y3 | Not reported | 21.64 | 218, 310, 430 | 467 |
| Y4 | Not reported | 25.40 | 218, 310, 430 | 495 |
| Y5 | Not reported | 23.98 | 278, 420 | |
| Y6 | Not reported | 28.20 | 278, 420 |
| Batch | Cultivation Time (day) | Color Characteristic | Batch Yield (AU410 nm) | Batch Productivity (AU410 nm/Day) | Cumulative Yield (AU410 nm) | Mean Productivity (AU410 nm/Day) | |
|---|---|---|---|---|---|---|---|
| AU410 nm/AU510 nm | AU410 nm/AU470 nm | ||||||
| 1 | 4 | 2.30 | 1.20 | 64.06 | 16.01 | 64.06 | 16.02 |
| 2 | 3 | 2.34 | 1.20 | 75.77 | 25.26 | 139.82 | 19.97 |
| 3 | 3 | 2.43 | 1.24 | 68.84 | 22.95 | 208.67 | 20.87 |
| 4 | 3 | 2.71 | 1.26 | 81.30 | 27.10 | 289.97 | 22.31 |
| 5 | 4 | 2.63 | 1.27 | 79.08 | 19.77 | 369.05 | 21.71 |
| 6 | 4 | 2.26 | 1.16 | 66.85 | 16.71 | 435.90 | 20.76 |
| 7 | 4 | 2.46 | 1.23 | 56.63 | 14.16 | 492.53 | 19.70 |
| Monascus Stain | Immobilization Carrier | Number of Batch and Time | Major Pigments | Mean Productivity |
|---|---|---|---|---|
| M. anka SCUT1123 | Ca-alginate | 5 batches, 20 days | Yellow | 15.38 AU410/day (this study) |
| M. purpureus C322 | Ca-alginate | 6 batches, 37 days | Orange | 4.7 AU470/day [27] |
| Monascus F4018 | Ca-alginate | 12 batches, 36 days | Red (uncertainty) a | 15~15.33 (AU420 + AU520)/day [22] |
| M. purpureus AS3.972 | Ca-alginate | 5 batches, 20 days | Red (uncertainty) a | 14.5~15 (AU420 + AU520)/day [24] |
| Monascus purpureus M183 | Ca-alginate | 7 batches, 44 days | uncertainty a | 4.76 (AU420 + AU470470 + AU505) [43] |
| M. purpureus | Polyelectrolyte complex | 14 batches, 10.33 days | Red | 3.79 AU500/day [23] |
| Monascus M101 | Polyvinyl alcohol | 12 batches, 60 days | Red | 9.25 AU520/day (without active carbon)36.34 AU520/day (with active carbon) [26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, K.; Zhao, Y.; Song, D.; Chen, G.; Wang, C.; Wu, Z.; Gu, H. Monascus Yellow Pigment Production by Coupled Immobilized-Cell Fermentation and Extractive Fermentation in Nonionic Surfactant Micelle Aqueous Solution. Fermentation 2023, 9, 168. https://doi.org/10.3390/fermentation9020168
Shi K, Zhao Y, Song D, Chen G, Wang C, Wu Z, Gu H. Monascus Yellow Pigment Production by Coupled Immobilized-Cell Fermentation and Extractive Fermentation in Nonionic Surfactant Micelle Aqueous Solution. Fermentation. 2023; 9(2):168. https://doi.org/10.3390/fermentation9020168
Chicago/Turabian StyleShi, Kan, Yuzhu Zhao, Da Song, Gong Chen, Chengtao Wang, Zhenqiang Wu, and Huawei Gu. 2023. "Monascus Yellow Pigment Production by Coupled Immobilized-Cell Fermentation and Extractive Fermentation in Nonionic Surfactant Micelle Aqueous Solution" Fermentation 9, no. 2: 168. https://doi.org/10.3390/fermentation9020168
APA StyleShi, K., Zhao, Y., Song, D., Chen, G., Wang, C., Wu, Z., & Gu, H. (2023). Monascus Yellow Pigment Production by Coupled Immobilized-Cell Fermentation and Extractive Fermentation in Nonionic Surfactant Micelle Aqueous Solution. Fermentation, 9(2), 168. https://doi.org/10.3390/fermentation9020168