

Overproduction of Laccase by Trametes versicolor and Pycnoporus sanguineus in Farnesol-Pineapple Waste Solid Fermentation

 , , , , ,
, , , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganisms
2.3. Evaluation of Mycelium Growth and Laccase Activity on PDA Cultures
2.4. Production of Laccase in Solid-State Fermentation (SSF) in the Presence and Absence of Farnesol
2.5. Laccase Enzyme Assay
2.6. Scanning Electron Microscopy
2.7. Determination of Catalase, Superoxide Dismutase, and Reactive Oxygen Species (ROS)
2.8. Physico-Chemical Properties of Laccases Produced in the Absence and Presence of Farnesol
2.9. Statistical Analysis
3. Results and Discussion
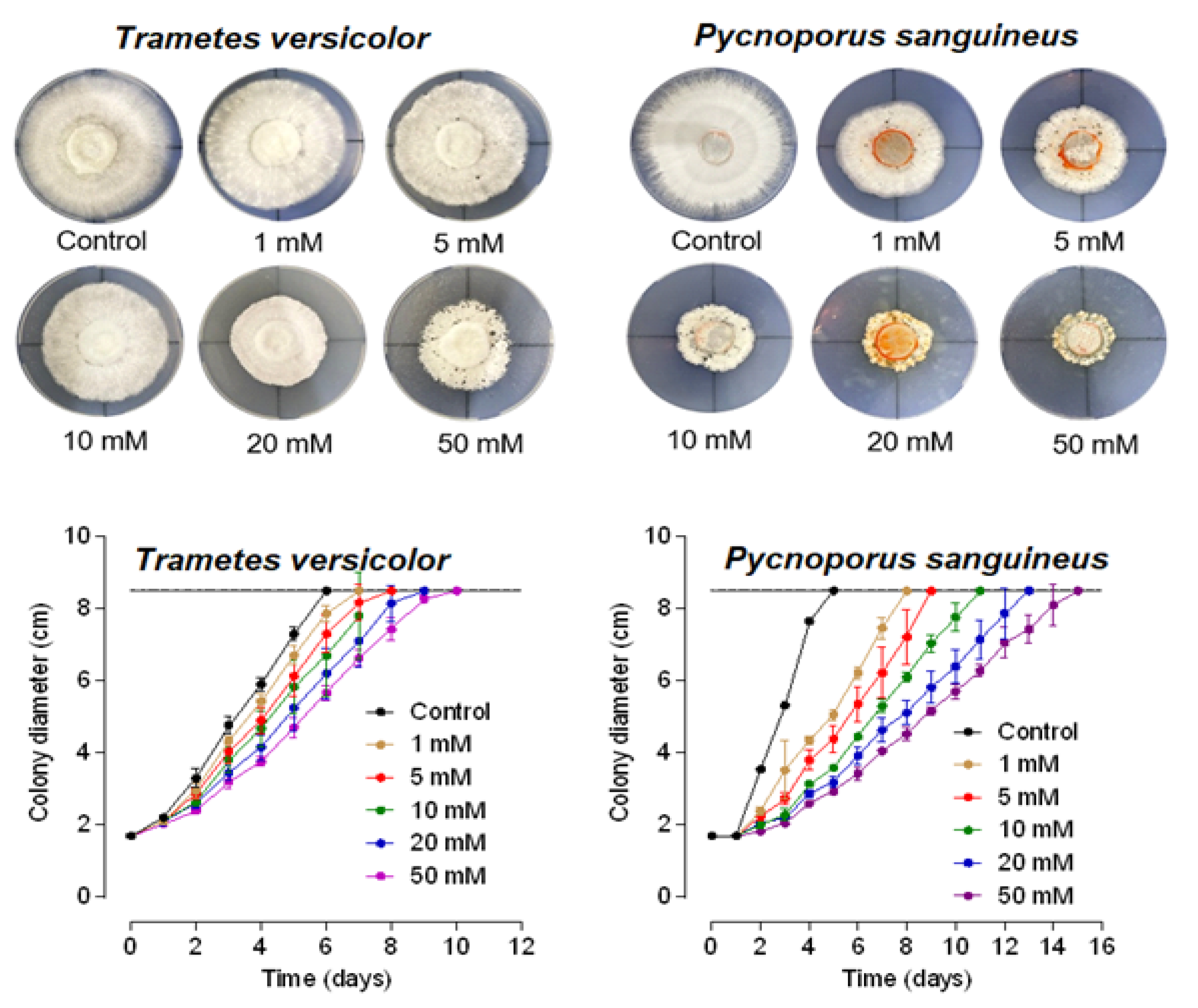
3.1. Inhibitory Effect of Farnesol on Mycelial Growth
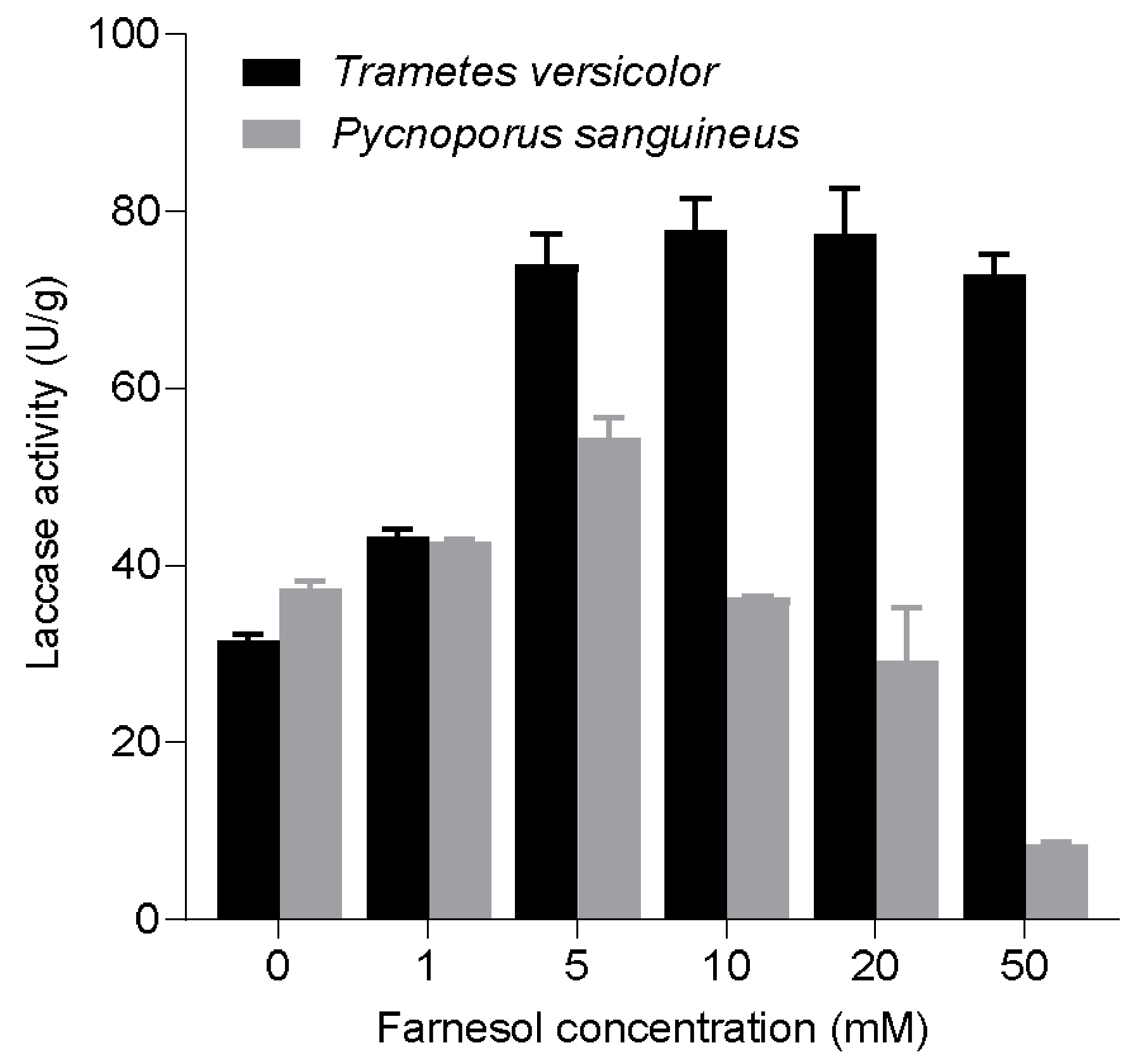
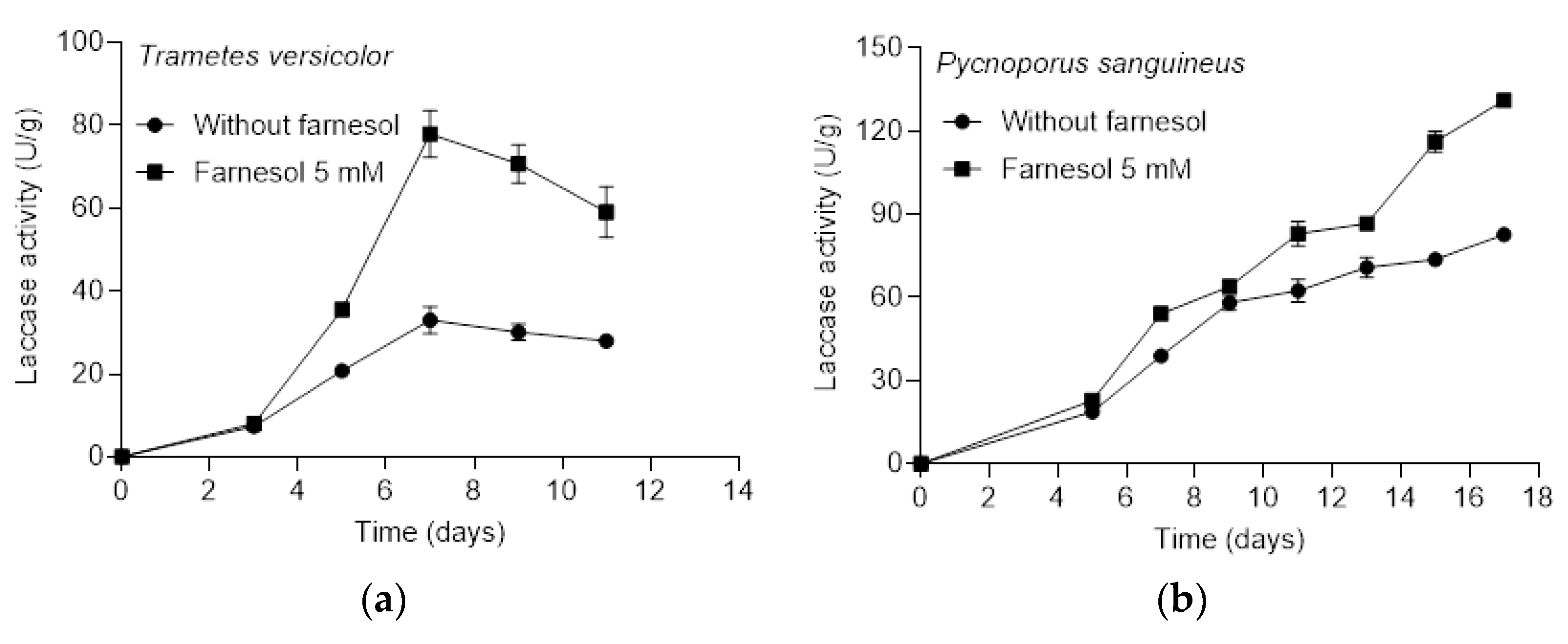
3.2. Effect of Farnesol on Laccase Production by T. versicolor and P. sanguineus in Solid-State Conditions
3.3. Effect of Farnesol on Hyphae Morphology and on Oxidative Stress
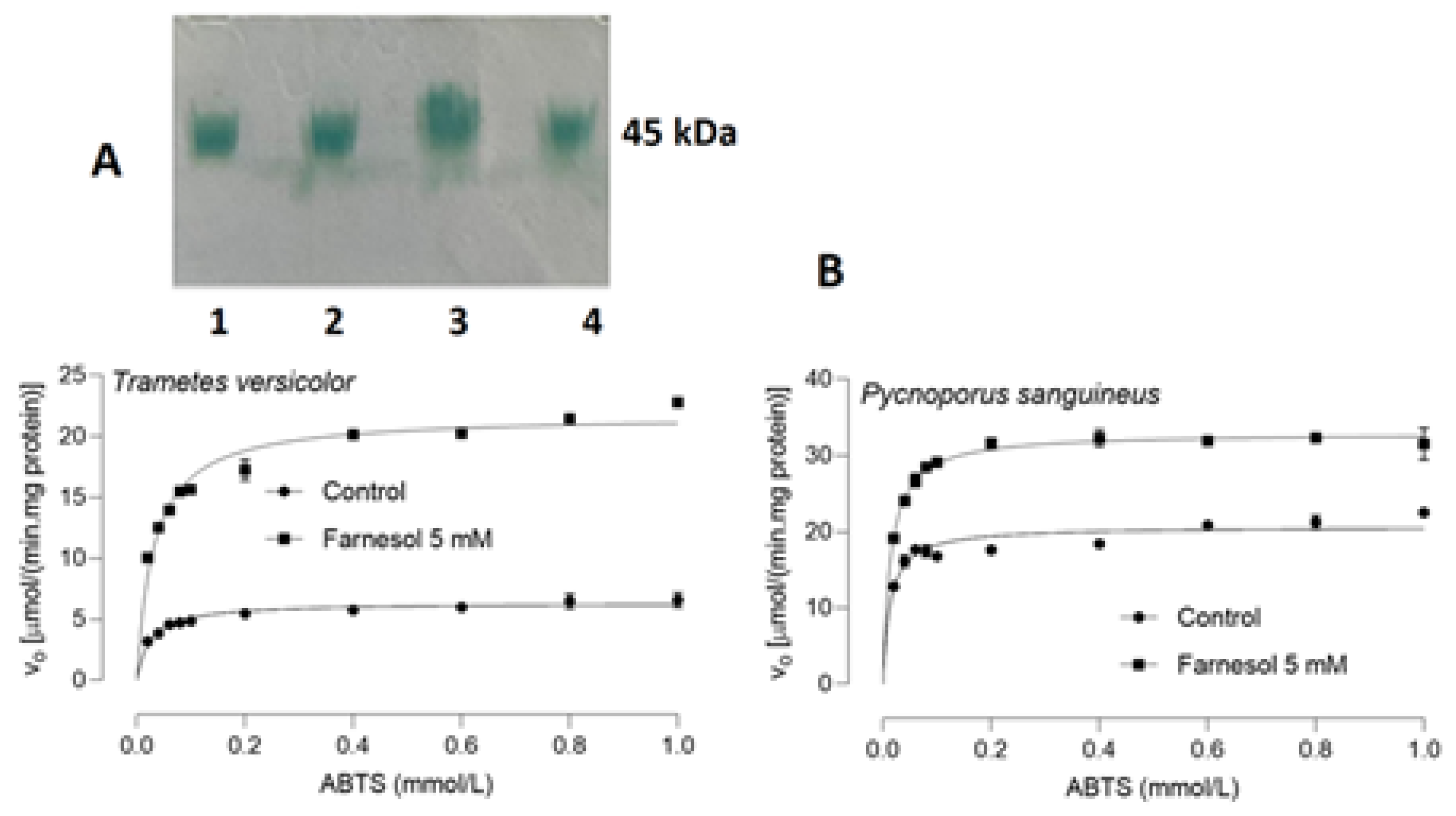
3.4. Physico-Chemical Properties of Laccases Produced in the Absence and Presence of Farnesol
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Backes, E.; Kato, C.G.; Corrêa, R.C.G.; Moreira, R.F.P.M.; Peralta, R.A.; Barros, L.; Ferreira, I.C.F.R.; Zanin, G.M.; Bracht, A.; Peralta, R.M. Laccases in food processing: Current status, bottlenecks and perspectives. Trends Food Sci. Technol. 2021, 115, 445–460. [Google Scholar] [CrossRef]
- Cruz-Vázquez, A.; Tomasini, A.; Armas-Tizapantzi, A.; Marcial-Quino, J. Montiel-González, A Extracellular proteases and laccases produced by Pleurotus ostreatus PoB: The effects of proteases on laccase activity. Int. Microbiol. 2022, 25, 495–502. [Google Scholar] [CrossRef]
- Aruwa, C.E.; Amoo, S.O.; Koorbanally, N.; Kudanga, Y. Laccase-mediated modification of isorhamnetin improves antioxidant and antibacterial activities. Proc. Biochem. 2022, 112, 53–61. [Google Scholar] [CrossRef]
- Backes, E.; Kato, C.G.; Silva, T.B.V.; Uber, T.M.; Pasquarelli, D.L.; Bracht, A.; Peralta, R.M. Production of fungal laccase on pineapple waste and application in detoxification of malachite green. J. Environ. Environ. Sci. Health Part B 2022, 57, 90–101. [Google Scholar] [CrossRef]
- Curran, L.M.C.L.K.; Pham, L.T.M.; Sale, K.L.; Simmons, B.A. Review of advances in the development of laccases for the valorization of lignin to enable the production of lignocellulosic biofuels and bioproduct. Biotechnol. Adv. 2022, 54, 107809. [Google Scholar] [CrossRef]
- Zofair, S.F.F.; Ahmad, S.; Hashmi, A.; Khan, S.H.; Khan, M.A.; Younus, H. Catalytic roles, immobilization, and management of recalcitrant environmental pollutants by laccases: Significance in sustainable green chemistry. J. Environ. Manage. 2022, 39, 114676. [Google Scholar] [CrossRef]
- Chmelová, D.; Legerská, B.; Kunstová, J.; Ondrejovič, M.; Miertuš, S. The production of laccases by white-rot fungi under solid-state fermentation conditions. World J. Microbiol. Biotechnol. 2022, 38, 21. [Google Scholar] [CrossRef]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, N.A. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid-state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microb. Biotechnol. 2017, 101, 5213–5222. [Google Scholar] [CrossRef] [PubMed]
- Soccol, C.R.; Costa, E.S.F.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; Vandenberghe, L.P.S. Recent developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- Velásquez-Quintero, C.; Merino-Restrepo, A.; Hormaza-Anaguano, A. Production, extraction, and quantification of laccase obtained from an optimized solid-state fermentation of corncob with white-rot fungi. J. Clean Prod. 2022, 370, 133598. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Mimi Sakinah, A.M.; Zularisam, A.W.; Sirohi, R.; Khilji, I.A.; Ahmad, N.; Pandey, A. Advances in solid-state fermentation for bioconversion of agricultural wastes to value-added products: Opportunities and challenges. Bioresour. Technol. 2022, 343, 12065. [Google Scholar] [CrossRef]
- Delmondes., G.A.; Bezerra, D.S.; Dias, D.Q.; Borges, A.S.; Araújo, I.M.; Cunha, G.L.; Bandeira, P.F.R.; Barbosa, R.; Coutinho, H.D.M.; Felipe, F.B.; et al. Toxicological and pharmacologic effects of farnesol (C15H26O): A descriptive systematic review. Food Chem. Toxicol. 2019, 129, 169–200. [Google Scholar] [CrossRef]
- Hu, J.; Wang, F.; Ma, A.; Zhuang, G.; Liu, Y.; Lu, J.; Guo, C.; Liu, C. Farnesol stimulates laccase production in Trametes versicolor. Eng. Life Sci. 2016, 16, 364–370. [Google Scholar] [CrossRef]
- Wang, K.F.; Guo, C.; Ju, F.; Samak, N.A.; Zhuang, G.Q.; Liu, C. Z Farnesol-induced hyperbranched morphology with short hyphae and bulbous tips of Coriolus versicolor. Sci. Rep. 2018, 8, 15213. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.J. A Convenient Growth Medium for Neurospora (Medium N). Microb. Genet. Bull 1956, 13, 42–43. [Google Scholar]
- Maciel, G.M.; Inácio, F.D.; Sá-Nakanishi, A.B.; Haminiuk, C.W.I.; Castoldi, R.; Comar, J.F.; Bracht, A.; Peralta, R.M. Response of Ganoderma lucidum and Trametes sp. to the herbicide picloram: Tolerance, antioxidants and production of ligninolytic enzymes. Pestic. Biochem. Physiol. 2013, 105, 84–92. [Google Scholar] [CrossRef]
- Freitas, E.N.; Bubna, G.A.; Brugnari, T.; Kato, C.G.; Nolli, M.; Rauen, T.G.; Moreira, R.F.P.M.; Peralta, R.A.; Bracht, A.; de Souza, C.G.M.; et al. Removal of bisphenol A by laccases from Pleurotus ostreatus and Pleurotus pulmonarius and evaluation of ecotoxicity of degradation products. Chem. Eng. J. 2017, 330, 1361–1369. [Google Scholar] [CrossRef]
- Rossignol, T.; Logue, M.E.; Reynolds, K.; Grenon, M.; Lowndes, N.F.; Butler, G. Transcriptional response of Candida parapsilosis following exposure to farnesol. Antimicrob. Agents Chemother. 2007, 51, 2304–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, L.; Chen, N.; Li, J.; Han, C.F.; Wang, S.; Hao, L.; Jia, S.; Han, P. The effects of quorum sensing molecule farnesol on the yield and activity of extracellular polysaccharide from Grifola frondosa in liquid fermentation. Int. J. Biol. Macrom. 2021, 191, 377–384. [Google Scholar] [CrossRef]
- Henriques, M.; Martins, M.; Azeredo, J.; Oliveira, J.R. Effect of farnesol on Candida dubliniensis morphogenesis. Lett. Appl. Microbiol. 2007, 44, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Prado, K.S.; Spinacé, M.A.S. Isolation and characterization of cellulose nanocrystals from pineapple crown waste and their potential uses. Int. J. Biol. Macromol. 2019, 122, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Salas, F.; Martínez, M.J.; Barriuso, J. Quorum-sensing mechanisms mediated by farnesol in Ophiostoma piceae: Effect on secretion of sterol esterase. Appl. Environ. Microbiol. 2015, 81, 4351–4357. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ma, A.; Zhao, G.; Yun, J.; Liu, X.; Zhang, H.; Zhuang, G. Effects of farnesol on Penicillium decumbens’s morphology and cellulase production. BioResources 2011, 6, 3252–3259. [Google Scholar]
- Wang, Z.; Liu, J.; Ning, Y.; Liao, X.; Jia, Y. Eichhornia crassipes: Agro-waster for a novel thermostable laccase production by Pycnoporus sanguineus SYBC-L1. J. Biosci. Bioeng. 2017, 123, 163–169. [Google Scholar] [CrossRef]
- Harris, S.D. Hyphal branching in filamentous fungi. Dev. Biol. 2019, 451, 35–39. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Q.; Wang, Q.; Qin, Y.; Zhong, Y.; Gao, L.; Liu, G.; Qu, Y. Disruption of the Trichoderma reesei gul1 gene stimulates hyphal branching and reduces broth viscosity in cellulase production. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab012. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Li, C.; Ma, L.; Zhang, D.; Chen, S. Effect of highly branched hyphal morphology on the enhanced production of cellulase in Trichoderma reesei DES-15. Biotech 2016, 6, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.F.; Sui, K.Y.; Guo, C.; Liu, C.Z. Quorum sensing molecule-farnesol increased the production and biological activities of extracellular polysaccharide from Trametes versicolor. Int. J. Biol. Macrom. 2017, 104, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Jaszek, M.; Grzywnowicz, K.; Malarczyk, E.; Leonowicz, A. Enhanced extracellular laccase activity as a part of the response system of white rot fungi: Trametes versicolor and Abortiporus biennis to paraquat-caused oxidative stress conditions. Pestic. Biochem. Physiol. 2006, 85, 147–154. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, K.K.; Kumar, P.; Ramchiary, N. Laccase isozymes from Ganoderma lucidum MDU-7: Isolation, characterization, catalytic properties and differential role during oxidative stress. J. Mol. Catal. B Enzym. 2015, 113, 68–75. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, F.; Ma, F.; Gong, Y.; Wan, X.; Jiang, M.; Zhang, X. Expression of the laccase gene from a white rot fungus in Pichia pastoris can enhance the resistance of this yeast to H2O2-mediated oxidative stress by stimulating the glutathione-based antioxidative system. Appl. Environ. Microbiol. 2012, 78, 5845–5854. [Google Scholar] [CrossRef] [Green Version]
- Pompeu, G.B.; Pietrobon, V.C.; Andreote, C.C.F.; Ferreira, L.F.R.; Aguiar, M.; Sartori, S.B.; Cruz, S.H.; Monteiro, R.T.R. Role of the antioxidant defense system during the production of lignocellulolytic enzymes by fungi. Int. Microbiol. 2019, 22, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Peng, C.; Han, Q.; Wang, M.; Zhou, B.Y.; Xiao, Y.; Fang, Z.; Kues, U. Coprinopsis cinerea uses laccase Lcc9 as a defense strategy to eliminate oxidative stress during fungal-fungal interactions. Appl. Environ. Microbiol. 2022, 88, e0176021. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Moreira, J.S.; Bracht, A.; Souza, A.C.S.; Oliveira, R.F.; Sá-Nakanishi, A.B.; Souza, C.G.M.; Peralta, R.M. Degradation of Diuron by Phanerochaete chrysosporium: Role of Ligninolytic Enzymes and Cytochrome P450. J. Biomed. Biotechnol. 2013, 2013, 251354. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.; Geng, A. Utilization of horticultural waste for laccase production by Trametes versicolor under solid-state fermentation. Appl. Biochem. Biotechnol. 2011, 163, 235–246. [Google Scholar] [CrossRef]
- Xu, L.; Sun, K.; Wang, F.; Zhao, L.; Hu, J.; Ma, H.; Ding, Z. Laccase production by Trametes versicolor in solid-state fermentation using tea residues as substrate and its application in dye decolorization. J. Environ. Manage. 2020, 270, 110904. [Google Scholar] [CrossRef] [PubMed]
- Lonappan, L.; Rouissi, T.; Laadila, M.A.; Brar, S.K.; Galan, L.H.; Verma, M.; Surampalli, R.Y. Agro-industrial-produced laccase for degradation of diclofenac and identification of transformation products. ACS Sustain. Chem. Eng. 2017, 5, 5772–5781. [Google Scholar] [CrossRef]
- Montoya, S.; Patiño, A.; Sánchez, O.J. Production of lignocellulolytic enzymes and biomass of Trametes versicolor from agro-industrial residues in a novel fixed-bed bioreactor with natural convection and forced aeration at pilot scale. Processes 2021, 9, 397. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K.; Jana, M.M.; Gupta, A. Enhancement in multiple lignolytic enzymes production for optimized lignin degradation and selectivity in fungal pretreatment of sweet sorghum bagasse. Bioresour. Technol. 2017, 236, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Vikineswary, S.N.; Abdullah, M.; Renuvathani, M.; Sekaran, M.; Pandey, A.; Jones, E.B.G. Productivity of laccase in solid substrate fermentation of selected agro-residues by Pycnoporus sanguineus. Bioresour. Technol. 2006, 97, 171–177. [Google Scholar] [CrossRef]
- Zimbardi, A.L.R.L.; Camargo, P.F.; Sibeli, C.; Aquino Neto, S.; Meleiro, L.P.; Rosa, J.C.; de Andrade, A.R.; Jorge, J.A.; Furriel, R.P.M. A high redox potential laccase from Pycnoporus sanguineus RP15: Potential application for dye decolorization. Int. J. Mol. Sci. 2016, 17, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Laccase Activity U/mL | |||
|---|---|---|---|
| Control | 1 mM Farnesol | 5 mM Farnesol | |
| T. versicolor | 0.61 ± 0.09 a | 1.10 ± 0.08 b | 2.07 ± 0.12 c |
| P. sanguineus | 0.71 ± 0.06 a | 1.56 ± 0.12 b | 0.88 ± 0.07 c |
| Fungus/Condition | Time Course | ||
|---|---|---|---|
| 4 Days | 7 Days | 10 Days | |
| ROS (nmols/mg Protein) | |||
| T. versicolor | |||
| Control | 20.00 ± 3.00 a,* | 34.00 ± 6.00 a,* | 28.00 ± 5.00 a,* |
| 5 mM farnesol | 42.00 ± 7.00 a,* | 68.00 ± 8.00 a,* | 93.00 ± 10.00 b,* |
| P. sanguineus | |||
| Control | 38.00 ± 4.00 a | 32.00 ± 5.00 a,* | 32.00 ± 5.00 a,* |
| 5 mM farnesol | 45.00 ± 6.00 a | 75.00 ± 5.00 b,* | 103.00 ± 8.00 c,* |
| Catalase (U per mg of protein) | |||
| T. versicolor | |||
| Control | 180 ± 24 a | 390 ± 45 b,* | 330 ± 40 b,* |
| 5 mM farnesol | 255 ± 32 a | 880 ± 100 b,* | 560 ± 50 c,* |
| P. sanguineus | |||
| Control | 140 ± 18 a,* | 270 ± 32 b,* | 510 ± 22 c,* |
| 5 mM farnesol | 315 ± 45 a,* | 460 ± 35 a,* | 730 ± 58 b,* |
| Superoxide dismutase (U per mg protein) | |||
| T. versicolor | |||
| Control | 9.00 ± 0.50 a,* | 7.10 ± 0.90 a | 5.15 ± 0.80 b,* |
| 5 mM farnesol | 19.63 ± 0.72 a,* | 12.42 ± 2.50 a | 12.64 ± 1.41 a,* |
| P. sanguineus | |||
| Control | 13.00 ± 0.83 a,* | 8.52 ± 0.60 b,* | 7.00 ± 0.50 b,* |
| 5 mM farnesol | 27.24 ± 1.00 a,* | 22.30 ± 1.40 a,* | 14.80 ± 2.10 b,* |
| White-Rot Fungi | Lignocellulosic Material (Substrate) | Inducer | Laccase Activity (U/g Substrate) | Reference |
|---|---|---|---|---|
| T. versicolor ATCC 20869 | Horticultural waste | Veratryl alcohol | 8.60 | [35] |
| T. versicolor CICC 14001 | Tea residues | Copper sulfate | 25.70 | [36] |
| T. versicolor ATCC 20869 | Apple pomace | Copper sulfate | 49.16 ± 4.50 | [37] |
| Pulp and paper solid waste | 52.40 ± 2.20 | |||
| T. versicolor PSUWC 430 | Oak sawdust, coffee husk, and corn bran | Copper and manganese sulfate | 6.37 | [38] |
| Coriolus versicolor MTCC 138 | Sweet sorghum bagasse | Copper sulfate | 58.20 ± 4.30 | [39] |
| Gallic acid | 42.10 ± 3.60 | |||
| Syringic acid | 67.40 ± 7.70 | |||
| T. versicolor | Pineapple crown leaves | Control Farnesol | 32.97 ± 1.99 77.88 ± 5.62 | This study |
| P. sanguineus SYBC-L1 | Flower stems | Copper sulfate and Gallic acid | 32.02 | [24] |
| P. sanguineus | Sago ‘hampas’ | Urea | 46.50 | [40] |
| P. sanguineus | Wheat bran and corncob | NH4Cl and CuSO4 | 138.60 | [41] |
| P. sanguineus | Pineapple crown leaves | Control Farnesol | 82.64 ± 1.99 130.95 ± 2.19 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Backes, E.; Kato, C.G.; de Oliveira Junior, V.A.; Uber, T.M.; dos Santos, L.F.O.; Corrêa, R.C.G.; Bracht, A.; Peralta, R.M. Overproduction of Laccase by Trametes versicolor and Pycnoporus sanguineus in Farnesol-Pineapple Waste Solid Fermentation. Fermentation 2023, 9, 188. https://doi.org/10.3390/fermentation9020188
Backes E, Kato CG, de Oliveira Junior VA, Uber TM, dos Santos LFO, Corrêa RCG, Bracht A, Peralta RM. Overproduction of Laccase by Trametes versicolor and Pycnoporus sanguineus in Farnesol-Pineapple Waste Solid Fermentation. Fermentation. 2023; 9(2):188. https://doi.org/10.3390/fermentation9020188
Chicago/Turabian StyleBackes, Emanueli, Camila Gabriel Kato, Verci Alves de Oliveira Junior, Thaís Marques Uber, Luís Felipe Oliva dos Santos, Rúbia Carvalho Gomes Corrêa, Adelar Bracht, and Rosane Marina Peralta. 2023. "Overproduction of Laccase by Trametes versicolor and Pycnoporus sanguineus in Farnesol-Pineapple Waste Solid Fermentation" Fermentation 9, no. 2: 188. https://doi.org/10.3390/fermentation9020188
APA StyleBackes, E., Kato, C. G., de Oliveira Junior, V. A., Uber, T. M., dos Santos, L. F. O., Corrêa, R. C. G., Bracht, A., & Peralta, R. M. (2023). Overproduction of Laccase by Trametes versicolor and Pycnoporus sanguineus in Farnesol-Pineapple Waste Solid Fermentation. Fermentation, 9(2), 188. https://doi.org/10.3390/fermentation9020188