
Mutagenesis of Novel Clostridial fusants for Enhanced Green Biobutanol Production from Agriculture Waste


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Experimental Procedure and Method
2.2.1. Culture Conditions and Medium Preparation
2.2.2. Mutagenesis Study
UV-Mutagenesis
Chemical Mutagenesis
2.2.3. Hydrolysis of WS
2.2.4. Biobutanol Production in Batch SSF
2.2.5. Statistical Analysis
3. Results and Analysis
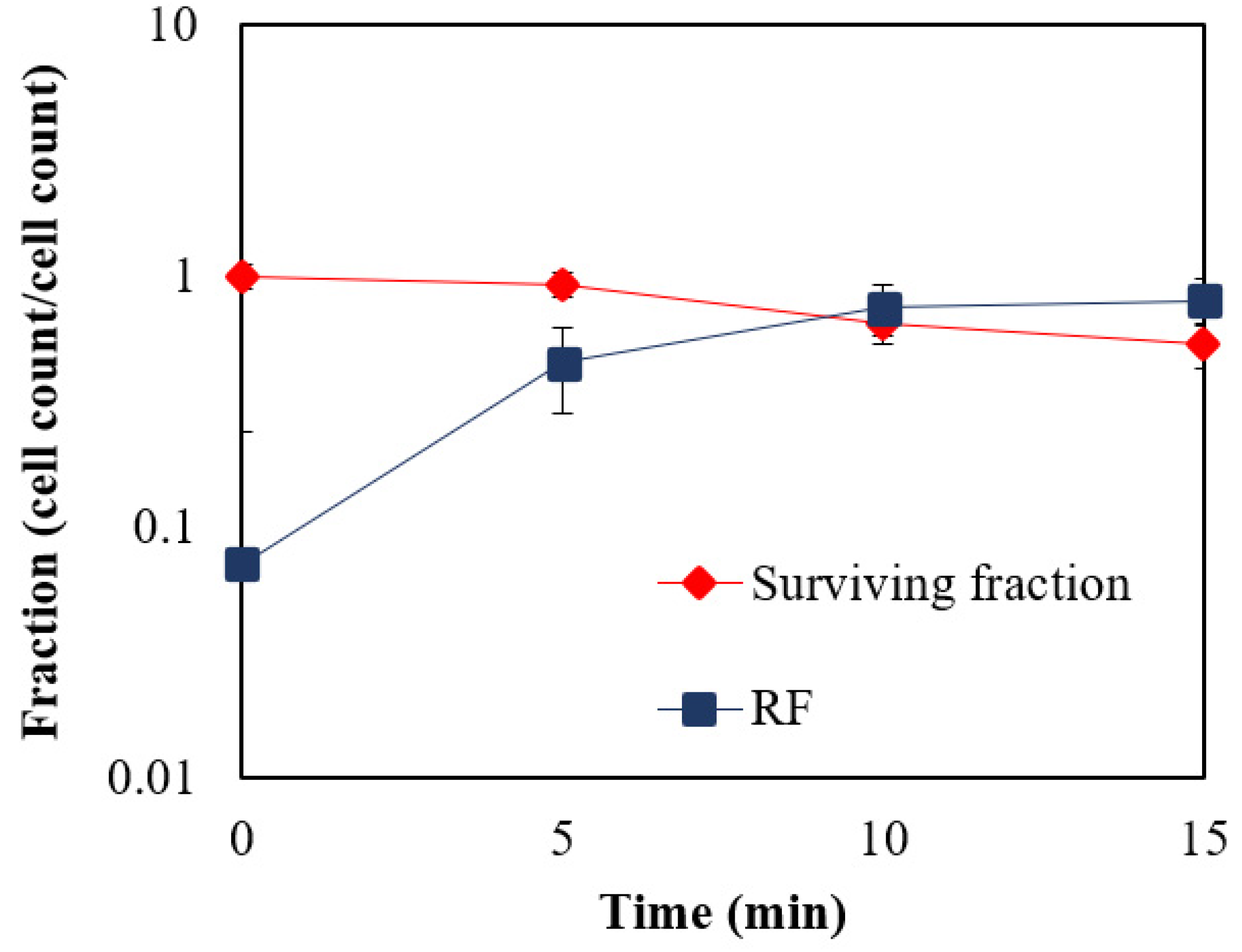
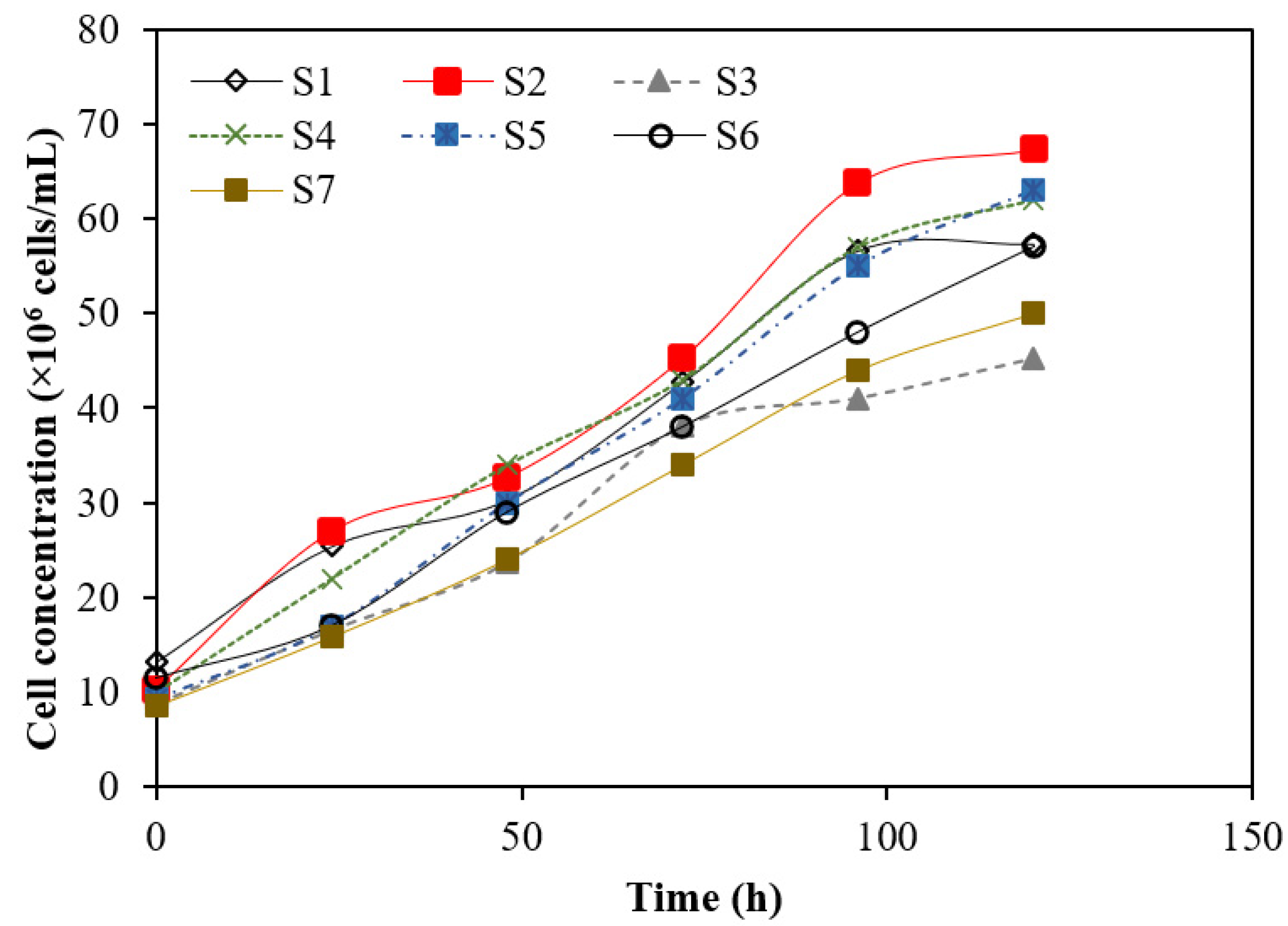
3.1. Mutagenesis of Bacterial Strains and Production of Biobutanol
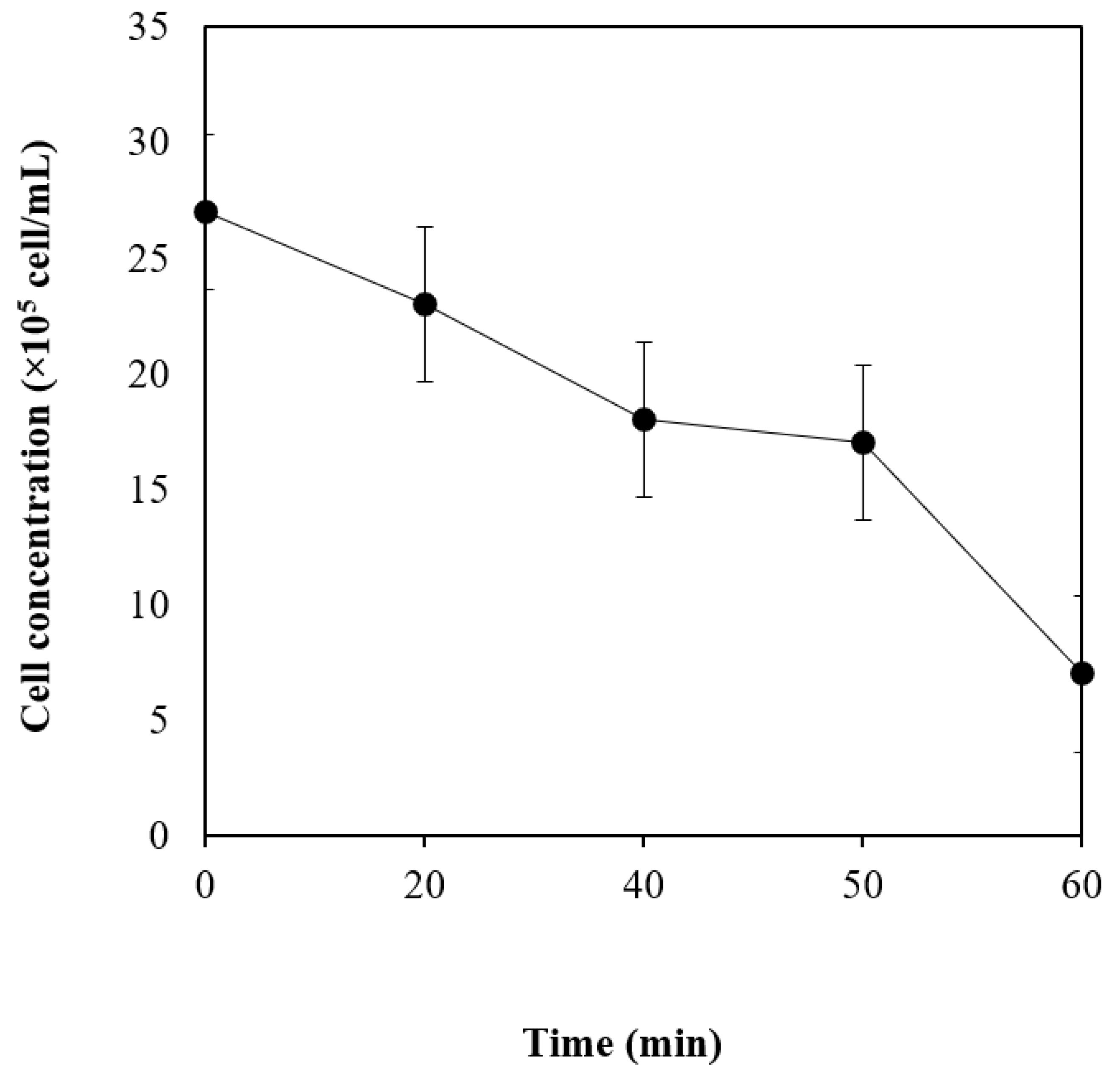
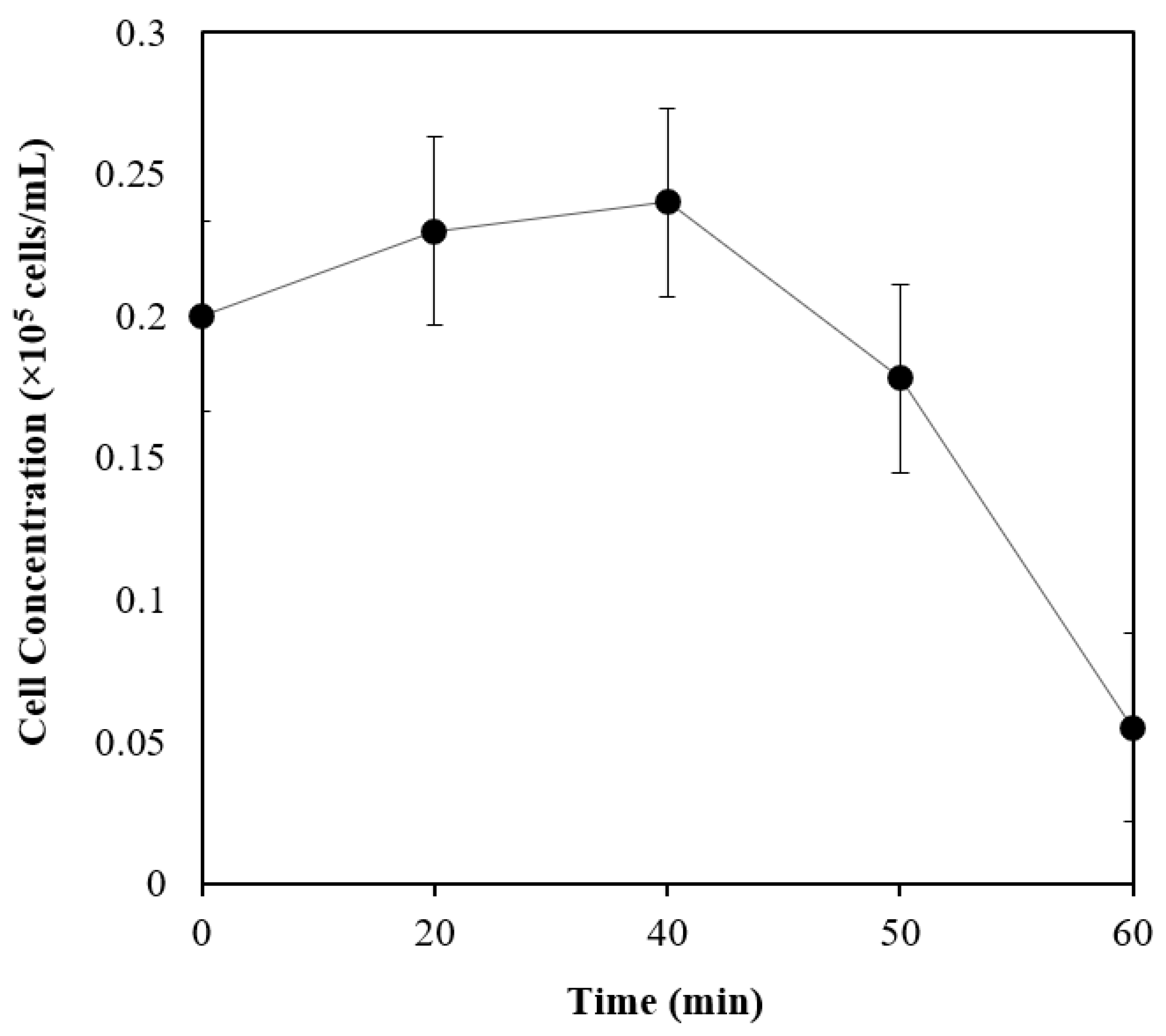
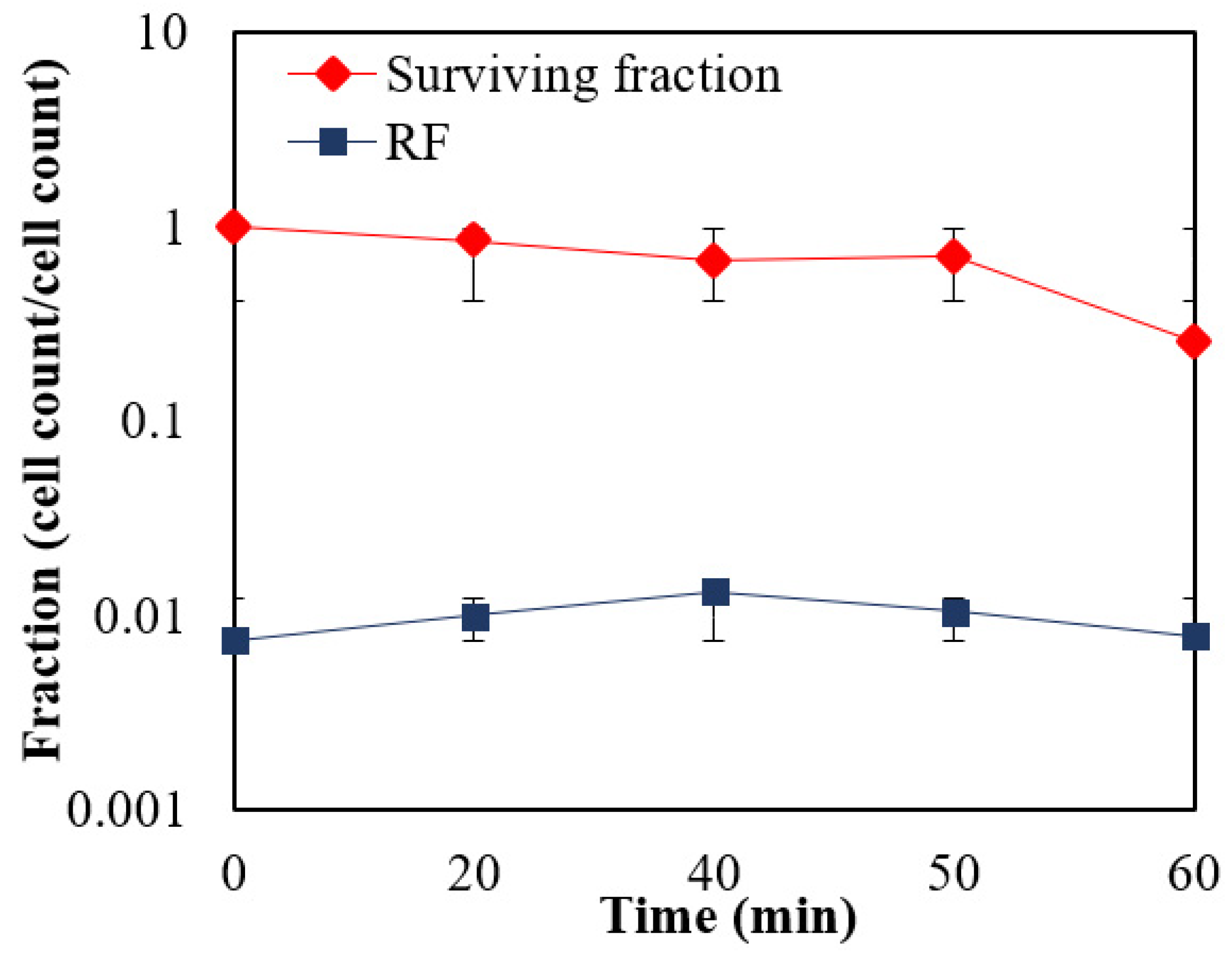
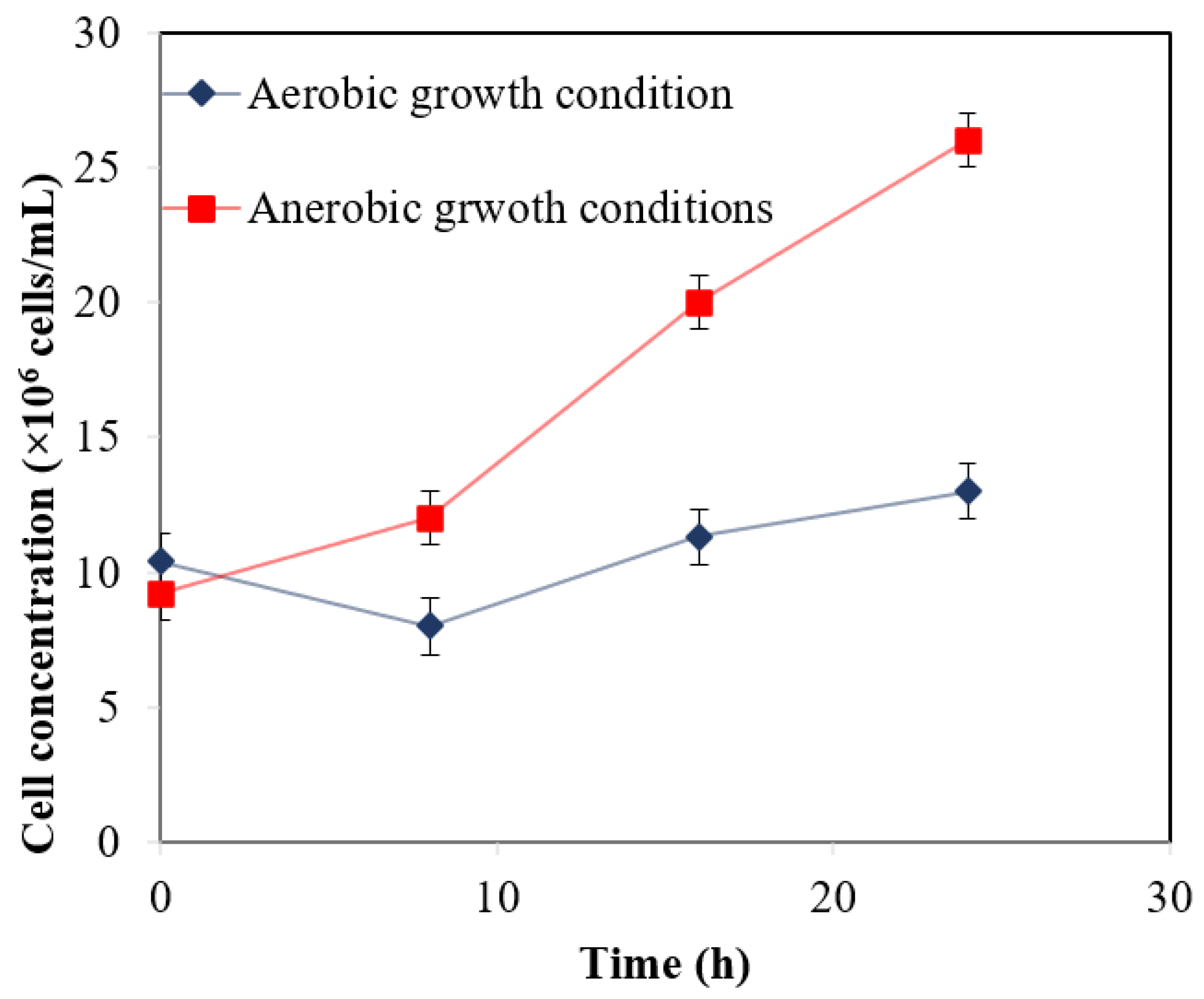
3.2. Oxygen Tolerance Studies
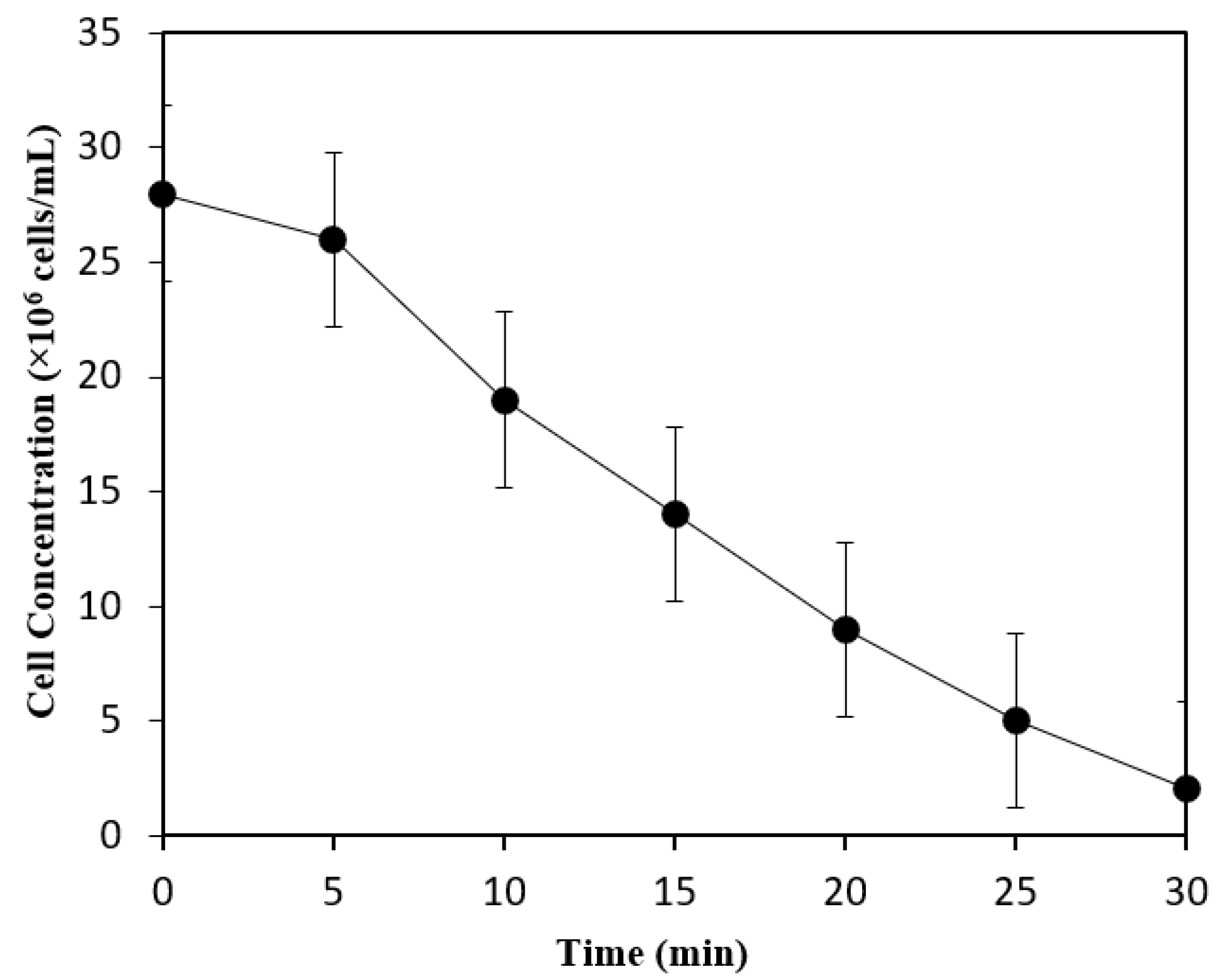
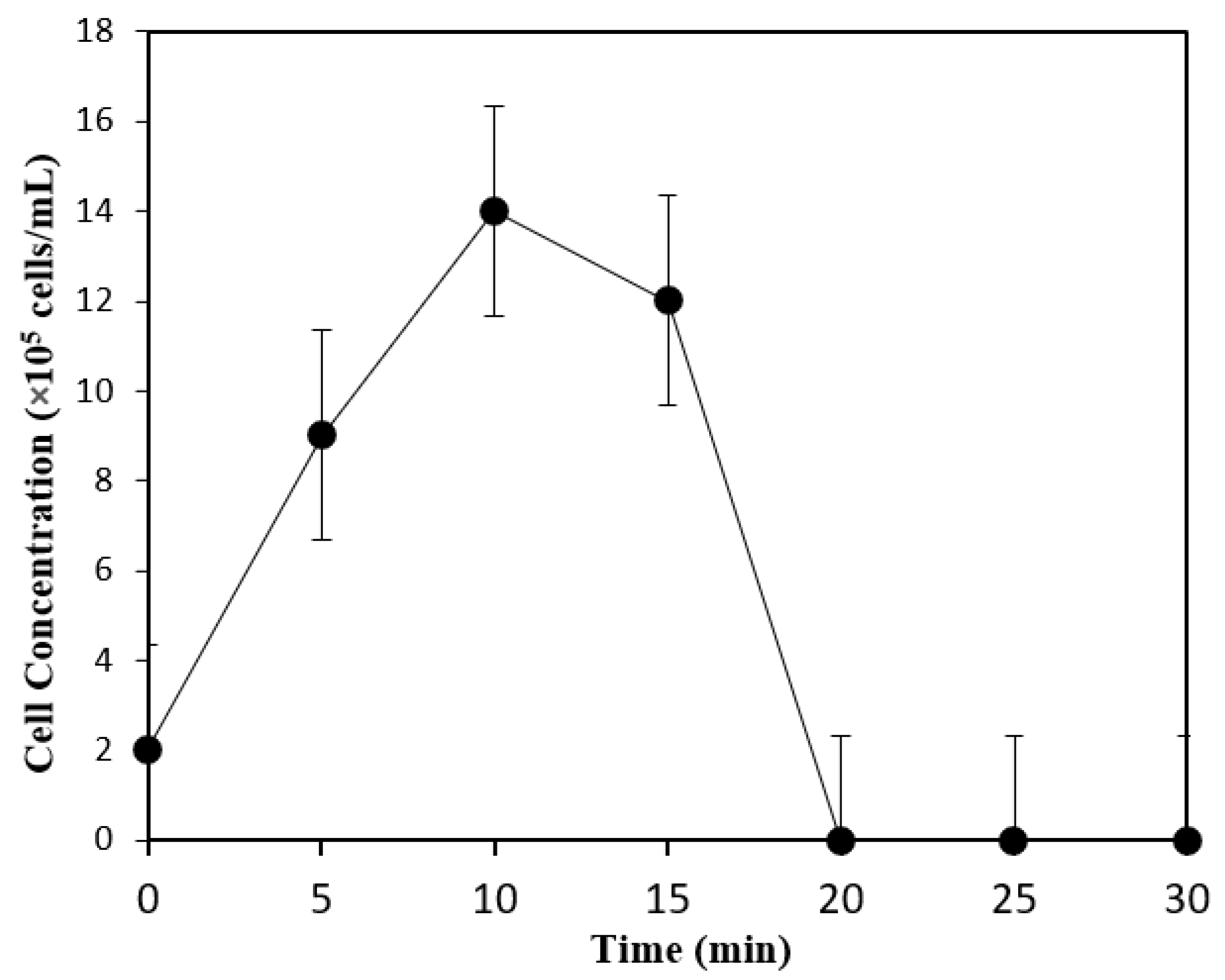
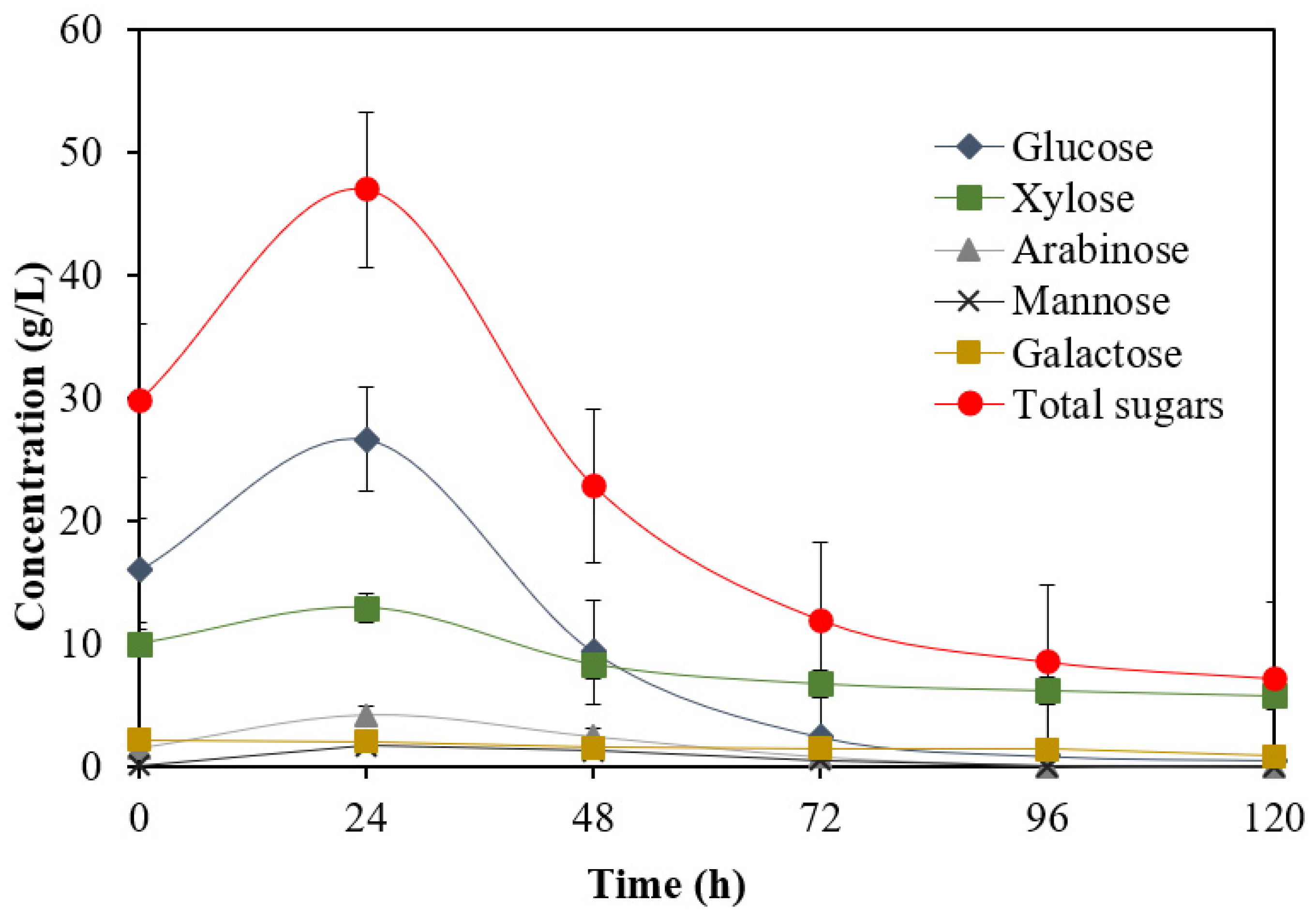
3.3. Biobutanol Production in Batch SSF
4. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| %RSD | Percent relative standard deviation |
| ABE | Acetone-biobutanol-ethanol |
| Cb | Clostridium beijerinckii |
| CbCt | Fused clostrial strain: Cb and Ct |
| CBM | Clostridium basal medium |
| CBP | Consolidated bioprocessing |
| Ct | Clostridium thermocellum |
| EMS | Ethyl methane sulphonate |
| GHG | Green house emissions |
| HPLC | High performance liquid chromatography |
| PEG | Polyethylene glycol |
| SD | Standard deviation |
| SSF | Simultaneous saccharification and fermentation |
| UV | Ultraviolet |
| WS | Wheat straw |
References
- Walsh, M.E.; Daniel, G.; Shapouri, H.; Slinsky, S.P. Bioenergy crop production in the United States: Potential quantities, land use changes and economic impacts on the agricultural sector. Environ. Resour. Econ. 2003, 24, 313–333. [Google Scholar] [CrossRef]
- Sorda, G.; Banse, M.; Kemfert, C. An overview of biofuel policies across the world. Energy Policy 2010, 38, 6977–6988. [Google Scholar] [CrossRef]
- Trejo, M.; Bhuyar, P.; Unpaprom, Y.; Dussadee, N.; Ramaraj, R. Advancement of fermentable sugars from fresh elephant ear plant weed for efficient bioethanol production. Environ. Dev. Sustain. 2022, 24, 7377–7387. [Google Scholar] [CrossRef] [PubMed]
- Ramaraj, R.; Bhuyar, P.; Intarod, K.; Sameechaem, N.; Unpaprom, Y. Stimulation of natural enzymes for germination of mimosa weed seeds to enhanced bioethanol production. Biotech 2021, 11, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Esfandabadi, Z.S.; Ranjbari, M.; Scagnelli, S. The imbalance of food and biofuel markets amid ukraine-russia crisis: A system thinking perspective. Biofuel Res. J. 2022, 9, 1640–1647. [Google Scholar] [CrossRef]
- Nigam, P.S.; Singh, A. Production of liquid biofuels from renewable resources. Prog. Energy Combust. Sci. 2011, 37, 52–68. [Google Scholar] [CrossRef]
- Dürre, P. Biobutanol: An attractive biofuel. Biotechnol. J. 2007, 2, 1525–1534. [Google Scholar] [CrossRef]
- Savage, N. Fuel options: The ideal biofuel. Nature 2011, 474, S9–S11. [Google Scholar] [CrossRef] [Green Version]
- Schoutens, G.H.; Nieuwenhuizen, M.C.H.; Kossen, N.W.F. Continuous biobutanol production from whey permeate with immobilized Clostridium beyerinckii LMD 27.6. Appl. Microbiol. Biotechnol. 1985, 21, 282–286. [Google Scholar] [CrossRef]
- Formanek, J.; Mackie, R.; Blaschek, H.P. Enhanced Biobutanol Production by Clostridium beijerinckii BA101 Grown in Semidefined P2 Medium Containing 6 Percent Maltodextrin or Glucose. Appl. Environ. Microbiol. 1997, 63, 2306–2310. [Google Scholar] [CrossRef]
- Dürre, P. New insights and novel developments in clostridial acetone/ biobutanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol. 1998, 49, 639–648. [Google Scholar] [CrossRef]
- Hahn-Hägerdal, B.; Galbe, M.; Gorwa-Grauslund, M.F.; Liden, G.; Zacchi, G. Bio-ethanol–the fuel of tomorrow from the residues of today. Trends Biotechnol. 2006, 24, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Syed, K.; Dahman, Y. Novel CLOSTRIDIAL FUSANTS in Comparison with Co-Cultured Counterparts Species for Enhanced Production of Biobutanol using Green Renewable and Sustainable Feedstock. Bioprocess Biosyst. Eng. 2015, 38, 2249–2262. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Dahman, Y. Enhanced Biobutanol production using novel Clostridial Fusants in simultaneous saccharification and fermentation of green renewable agriculture residues. Biofuels Bioprod. Biorefining 2015, 9, 529–544. [Google Scholar] [CrossRef]
- Mohtasebi, B.; Maki, M.; Qin, W.; Dahman, Y. Novel fusants of two and three Clostridia for enhanced green production of biobutanol. Biofuels 2019, 12, 1017–1027. [Google Scholar] [CrossRef]
- Helgeson, J.P.; Hunt, G.J.; Haberlach, G.T.; Austin, S. Somatic hybrids between Solanum brevidens and Solanum tuberosum: Expression of a late blight resistance gene and potato leaf roll resistance. Plant Cell Rep. 1986, 5, 212–214. [Google Scholar] [CrossRef]
- Gokhale, D.V.; Puntambekar, U.S.; Deobagkar, D.N. Protoplast fusion: A tool for intergeneric gene transfer in bacteria. Biotechnol. Adv. 1993, 11, 199–217. [Google Scholar] [CrossRef]
- Sarkar, D.; Schilde-Rentschler, L.; Löffelhardt-Ruoss, B.; Ninnemann, H. An electrofusion-mediated improved method for donor-recipient somatic hybridization in potato (Solanum tuberosum L.). In Recent Trends in Horticultural Biotechnology; New India Publishing Agency: New Delhi, India, 2007; pp. 129–147. [Google Scholar]
- Jurgens, G.; Survase, S.; Berezina, O.; Sklavounos, E.; Linnekoski, J.; Kurkijärvi, A.; Väkevä, M.; van Heiningen, A.; Granström, T. Butanol production from lignocellulosics. Biotechnol. Lett. 2012, 34, 1415–1434. [Google Scholar] [CrossRef]
- Liu, S.; Qureshi, N. How microbes tolerate ethanol and biobutanol. New Biotechnol. 2009, 26, 117–121. [Google Scholar] [CrossRef]
- Qurat-ul-Ain Syed Nadeem, M.; Nelofer, R. Enhanced biobutanol production by mutant strains of Clostridium acetobutylicum in molasses medium. Turk. J. Biochem. 2008, 33, 25–30. [Google Scholar]
- Lütke-Eversloh, T.; Bahl, H. Metabolic engineering of Clostridium acetobutylicum: Recent advances to improve biobutanol production. Curr. Opin. Biotechnol. 2011, 22, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shen, H.; Zhang, X.; Yu, X.; Wang, H.; Xiao, S.; Wang, J.; Zhao, Z.K. Combined mutagenesis of Rhodosporidium toruloides for improved production of carotenoids and lipids. Biotechnol. Lett. 2016, 38, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Nochur, S.V.; Roberts, M.F.; Demain, A.L. Mutation of Clostridium thermocellum in the presence of certain carbon sources. FEMS Microbiol. Lett. 1990, 71, 199–204. [Google Scholar] [CrossRef]
- Thompson, W.; Meyer, S.; Westhoff, P. How does petroleum price and corn yield volatility affect ethanol markets with and without an ethanol use mandate? Energy Policy 2009, 37, 745–749. [Google Scholar] [CrossRef]
- Kenyon, C.J. The bacterial response to DNA damage. Trends Biochem. Sci. 1983, 8, 84–87. [Google Scholar] [CrossRef]
- Bowring, S.N.; Morris, J.G. Mutagenesis of Clostridium acetobutylicum. J. Appl. Microbiol. 1985, 58, 577–584. [Google Scholar] [CrossRef]
- Lemmel, S.A. Mutagenesis in Clostridium acetobutylicum. Biotechnol. Lett. 1985, 7, 711–716. [Google Scholar] [CrossRef]
- Qureshi, N.; Blaschek, H. Evaluation of recent advances in biobutanol fermentation, upstream, and downstream processing. Bioprocess Biosyst. Eng. 2001, 24, 219–226. [Google Scholar] [CrossRef]
- Qureshi, N.; Ezeji, T.C. Biobutanol: A superior biofuel production from agricultural residues (renewable biomass): Recent progress in technology. Biofuels Bioprod. Biorefining 2008, 2, 319–330. [Google Scholar] [CrossRef]
- Talebnia, F.; Karakashev, D.; Angelidaki, I. Production of bioethanol from wheat straw: An overview on pretreatment, hydrolysis and fermentation. Bioresour. Technol. 2010, 101, 4744–4753. [Google Scholar] [CrossRef]
- Qureshi, N.; Saha, B.C.; Hector, R.E.; Hughes, S.R.; Cotta, M.A. Biobutanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part I—Batch fermentation. Biomass Bioenergy 2008, 32, 168–175. [Google Scholar] [CrossRef]
- Rogers, P.; Palosaari, N. Clostridium acetobutylicum mutants that produce butyraldehyde and altered quantities of solvents. Appl. Environ. Microbiol. 1987, 53, 2761–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikehata, H.; Ono, T. The mechanisms of UV mutagenesis. J. Radiat. Res. 2011, 52, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillmann, F.; Fischer, R.J.; Saint-Prix, F.; Girbal, L.; Bahl, H. PerR acts as a switch for oxygen tolerance in the strict anaerobe Clostridium acetobutylicum. Mol. Microbiol. 2008, 68, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.; Saha, B.; Cotta, M.A. Biobutanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng. 2007, 30, 419–427. [Google Scholar] [CrossRef]
- Ezeji, T.; Qureshi, N.; Blaschek, H.P. Production of acetone–biobutanol–ethanol (ABE) in a continuous flow bioreactor using degermed corn and Clostridium beijerinckii. Process Biochem. 2007, 42, 34–39. [Google Scholar] [CrossRef]
- Fond, O.; Engasser, J.M.; Matta-El-Amouri, G.; Petitdemange, H. The acetone biobutanol fermentation on glucose and xylose. I. Regulation and kinetics in batch cultures. Biotechnol. Bioeng. 1986, 28, 160–166. [Google Scholar] [CrossRef]
- El Kanouni, A.; Zerdani, I.; Zaafa, S.; Znassni, M.; Loutfi, M.; Boudouma, M. The improvement of glucose/xylose fermentation by Clostridium acetobutylicum using calcium carbonate. World J. Microbiol. Biotechnol. 1998, 14, 431–435. [Google Scholar] [CrossRef]
- Ounine, K.; Petitdemange, H.; Raval, G.; Gay, R. Regulation and biobutanol inhibition of D-xylose and D-glucose uptake in Clostridium acetobutylicum. Appl. Environ. Microbiol. 1985, 49, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Mielenz, J.R.; Xiao, X.; Ge, Y.; Hamilton, C.Y.; Rodriguez, M.; Wang, Z.Y. Genetic manipulation of lignin reduces recalcitrance and improves ethanol production from switchgrass. Proc. Natl. Acad. Sci. USA 2011, 108, 3803–3808. [Google Scholar] [CrossRef] [Green Version]
- Baronofsky, J.J.; Schreurs, W.J.; Kashket, E.R. Uncoupling by acetic acid limits growth of and acetogenesis by Clostridium thermoaceticum. Appl. Environ. Microbiol. 1984, 48, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, S.P.; Pierson, M.D. “Phoenix phenomenon” in the growth of Clostridium perfringens. Appl. Environ. Microbiol. 1976, 32, 803–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.M.; Moreira, A.R. Growth inhibition kinetics for the acetone-biobutanol fermentation. In ACS Symposium Series; ACS Publication: Washinton, DC, USA, 1983. [Google Scholar]
- Leung, J.C.Y.; Wang, D.I.C. Production of acetone and biobutanol by Clostridium acetobutylicum in continuous culture using free cells and immobilized cells. Appl. Microbiol. Biotechnol. 1981, 1, 348–352. [Google Scholar] [CrossRef]
- Friedberg, E.C.; Walker, G.C.; Siede, W. DNA Repair and Mutagenesis; American Society for Microbiology: Washington, DC, USA, 1995; ISBN 1-55581-319-4. [Google Scholar]
- Aghbashlo, M.; Khounani, Z.; Hosseinzadeh-Bandbafha, H.; Gupta, V.K.; Amiri, H.; Lam, S.S.; Morosuk, T.; Tabatabaei, M. Exergoenvironmental analysis of bioenergy systems: A comprehensive review. Renew. Sustain. Energy Rev. 2021, 149, 111399. [Google Scholar] [CrossRef]
- Soltanian, S.; Aghbashlo, M.; Almasi, F.; Hosseinzadeh-Bandbafha, H.; Nizami, A.S.; Ok, Y.S.; Lam, S.S.; Tabatabaei, M. A critical review of the effects of pretreatment methods on the exergetic aspects of lignocellulosic biofuels. Energy Convers. Manag. 2020, 212, 112792. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Mutation Parameters | Glucose | Xylose | Arabinose | Mannose | Galactose | Total Sugar (g/L) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Maximum * (g/L) | Final (g/L) | Maximum (g/L) | Final (g/L) | Maximum (g/L) | Final (g/L) | Maximum (g/L) | Final (g/L) | Maximum (g/L) | Final (g/L) | |||
| S1 | Control | 25.6 | 0.52 | 12.9 | 5.76 | 4.2 | 0 | 1.65 | 0 | 2.2 | 0.87 | 39.77 |
| S2 | 10 min UV | 27.1 | 0.50 | 13 | 5.71 | 4.2 | 0 | 1.67 | 0 | 2.2 | 0.88 | 41.26 |
| S3 | 15 min UV | 25.9 | 0.6 | 12.6 | 5.74 | 4.5 | 0.01 | 1.5 | 0 | 2.2 | 0.89 | 39.88 |
| S4 | 20 min EMS | 26.4 | 0.56 | 14.2 | 5.74 | 4.39 | 0.02 | 1.26 | 0 | 2.2 | 1.02 | 39.53 |
| S5 | 40 min EMS | 26.9 | 0.52 | 14.3 | 5.73 | 4.3 | 0.03 | 1.35 | 0.01 | 2.2 | 1.05 | 40.11 |
| S6 | 50 min EMS | 26.7 | 0.51 | 13 | 5.77 | 4.21 | 0.01 | 1.64 | 0.01 | 2.2 | 0.9 | 39.77 |
| S7 | 60 min EMS | 25.9 | 0.53 | 12.9 | 5.91 | 4.19 | 0.02 | 1.4 | 0.01 | 2.2 | 1.04 | 38.47 |
| Strain | Total Sugar Consumed (g/L) | ABE Yield (g/gsugars) * | Total ABE Yield | ||
|---|---|---|---|---|---|
| Acetone Yield | Butanol Yield | Ethanol Yield | |||
| S1 | 39.77 | 0.15 | 0.342 | 0.057 | 0.55 |
| S2 | 41.26 | 0.174 | 0.358 | 0.067 | 0.60 |
| S3 | 39.88 | 0.17 | 0.356 | 0.065 | 0.59 |
| S4 | 39.53 | 0.172 | 0.346 | 0.060 | 0.57 |
| S5 | 40.11 | 0.172 | 0.361 | 0.062 | 0.59 |
| S6 | 39.77 | 0.173 | 0.357 | 0.060 | 0.59 |
| S7 | 38.47 | 0.171 | 0.356 | 0.062 | 0.59 |
| Reference/Notes | Feedstock | Microbial | Biobutanol (g/L) |
|---|---|---|---|
| Qureshi and Ezeji, 2008 (Ref. [30]) | Glucose | Cb | 13 |
| Qureshi et al., 2007 (Ref. [36]) | WS | Cb | 7.4 |
| Dahman et al., 2019 (Ref. [15]) | WS | CbCt | 13.82 |
| Dahman et al., 2015 (Ref. [13]) | WS | CbCt | 13.81 |
| Current Study (Sample S2-UV) | WS | CbCt mutated | 15.0 |
| Current Study (Sample S4-EMS) | WS | CbCt mutated | 14.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, P.; Dahman, Y. Mutagenesis of Novel Clostridial fusants for Enhanced Green Biobutanol Production from Agriculture Waste. Fermentation 2023, 9, 92. https://doi.org/10.3390/fermentation9020092
Roy P, Dahman Y. Mutagenesis of Novel Clostridial fusants for Enhanced Green Biobutanol Production from Agriculture Waste. Fermentation. 2023; 9(2):92. https://doi.org/10.3390/fermentation9020092
Chicago/Turabian StyleRoy, Pallavi, and Yaser Dahman. 2023. "Mutagenesis of Novel Clostridial fusants for Enhanced Green Biobutanol Production from Agriculture Waste" Fermentation 9, no. 2: 92. https://doi.org/10.3390/fermentation9020092
APA StyleRoy, P., & Dahman, Y. (2023). Mutagenesis of Novel Clostridial fusants for Enhanced Green Biobutanol Production from Agriculture Waste. Fermentation, 9(2), 92. https://doi.org/10.3390/fermentation9020092