
Investigation of Geraniol Biotransformation by Commercial Saccharomyces Yeast Strains by Two Headspace Techniques: Solid-Phase Microextraction Gas Chromatography/Mass Spectrometry (SPME-GC/MS) and Proton Transfer Reaction-Time of Flight-Mass Spectrometry (PTR-ToF-MS)
 ,
,  , ,
, ,  ,
, 
Abstract
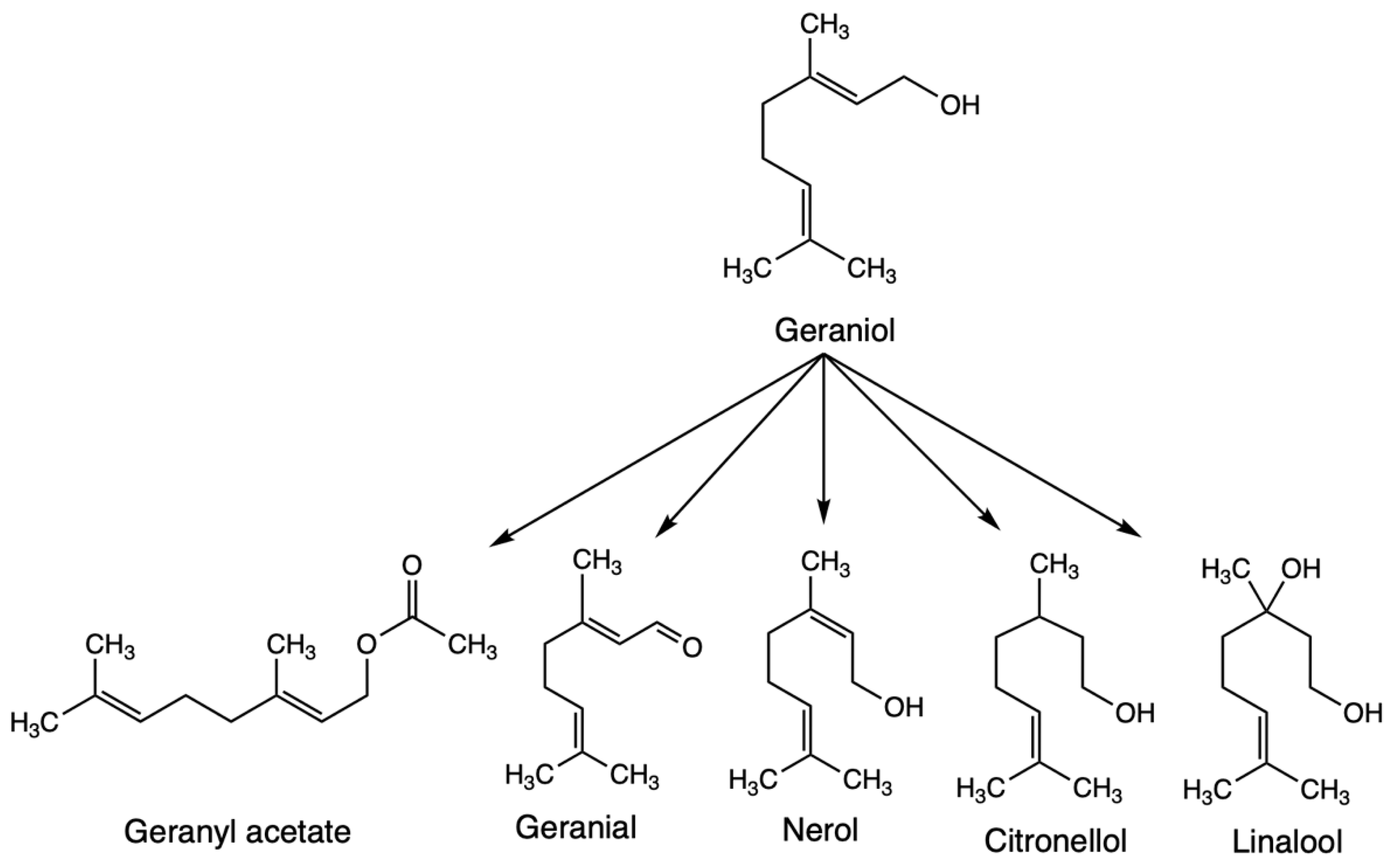
:1. Introduction
2. Materials and Methods
2.1. Yeast Hydration and Model Wort Preparation
2.2. Micro-Fermentations
2.3. HS- SPME-GC/MS Analytical Conditions
2.4. PTR-ToF-MS Measurement
Fragmentation Pattern Measurement
2.5. Data Analysis
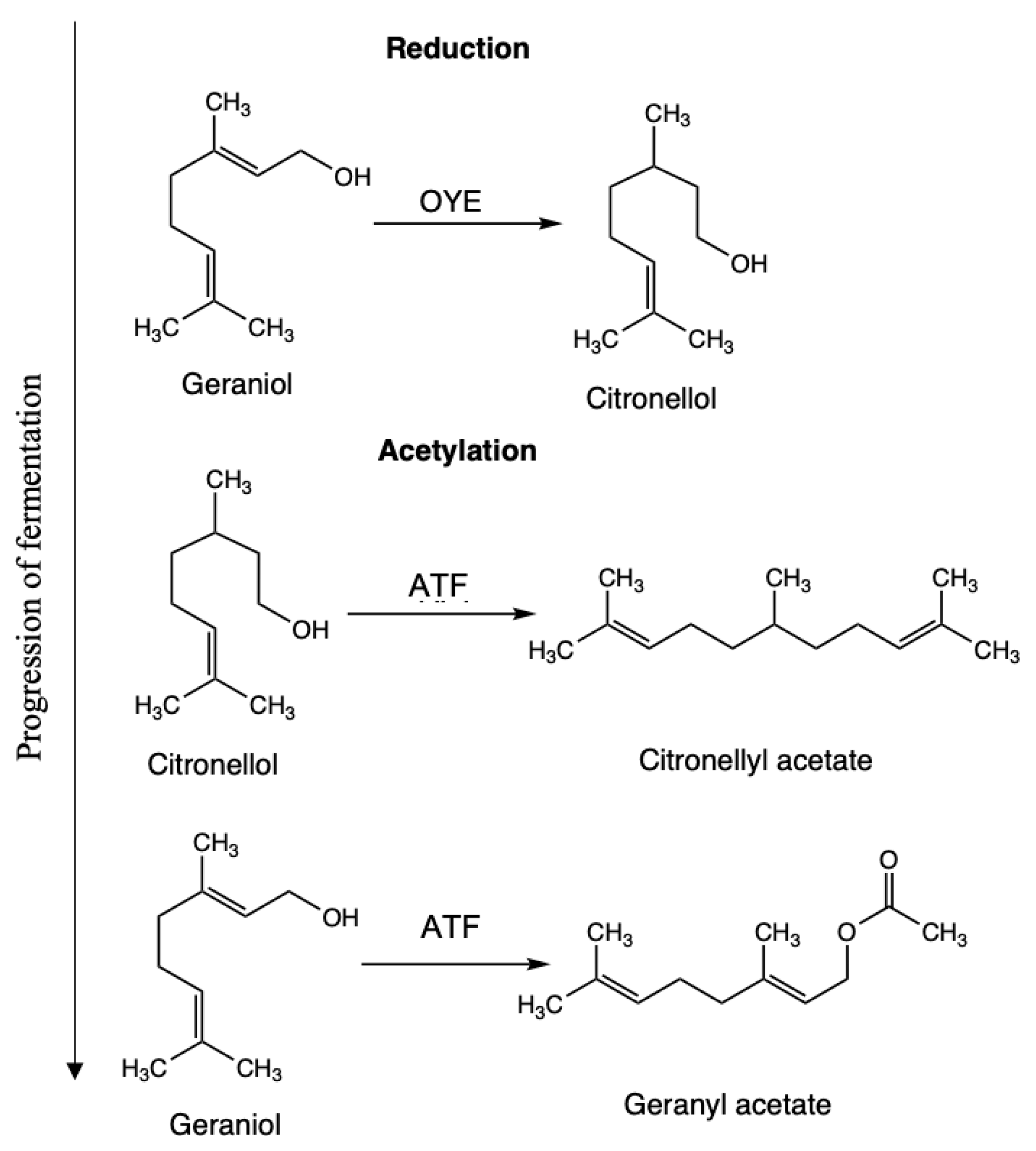
3. Results and Discussion
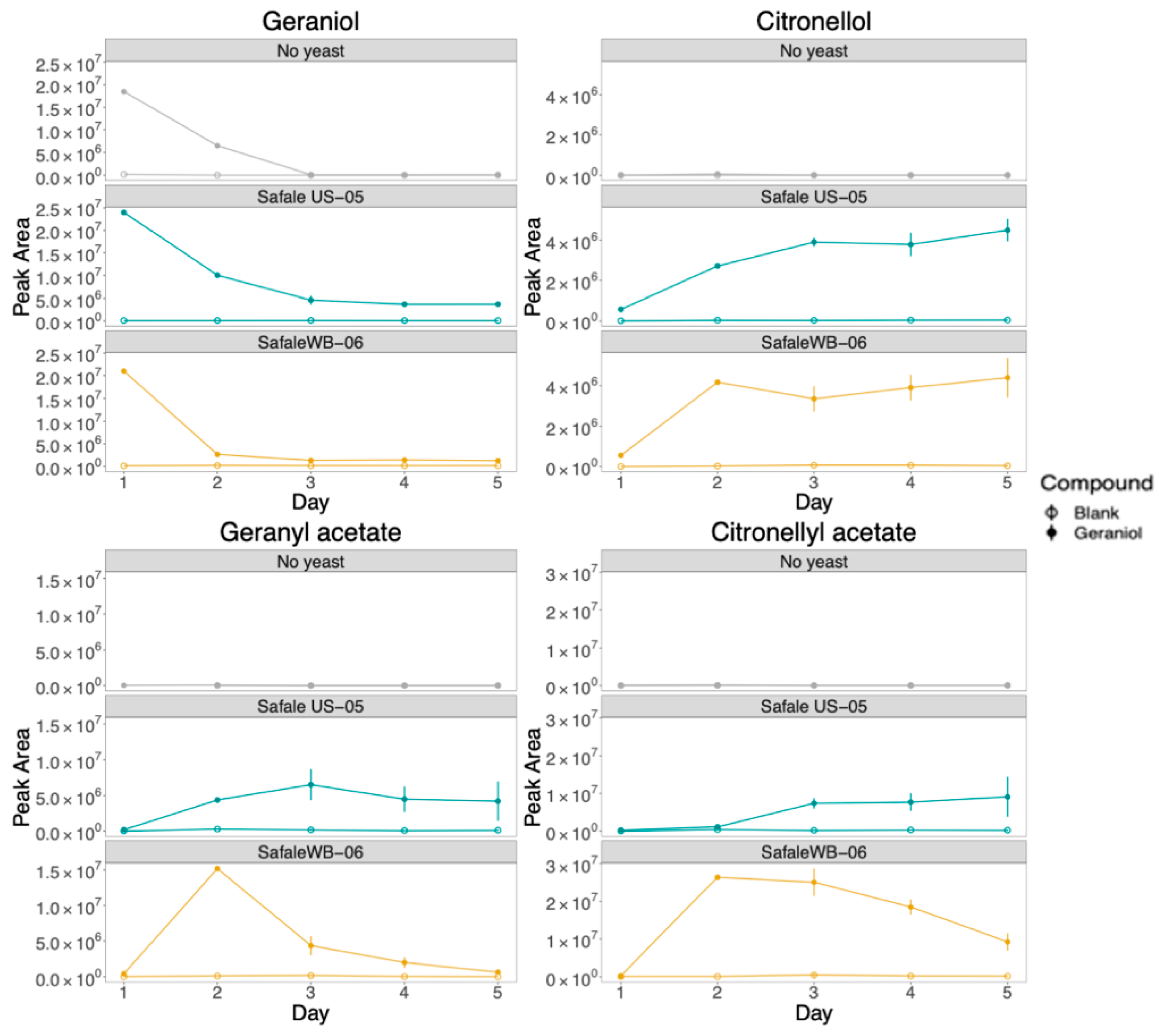
3.1. SPME-GC/MS Results
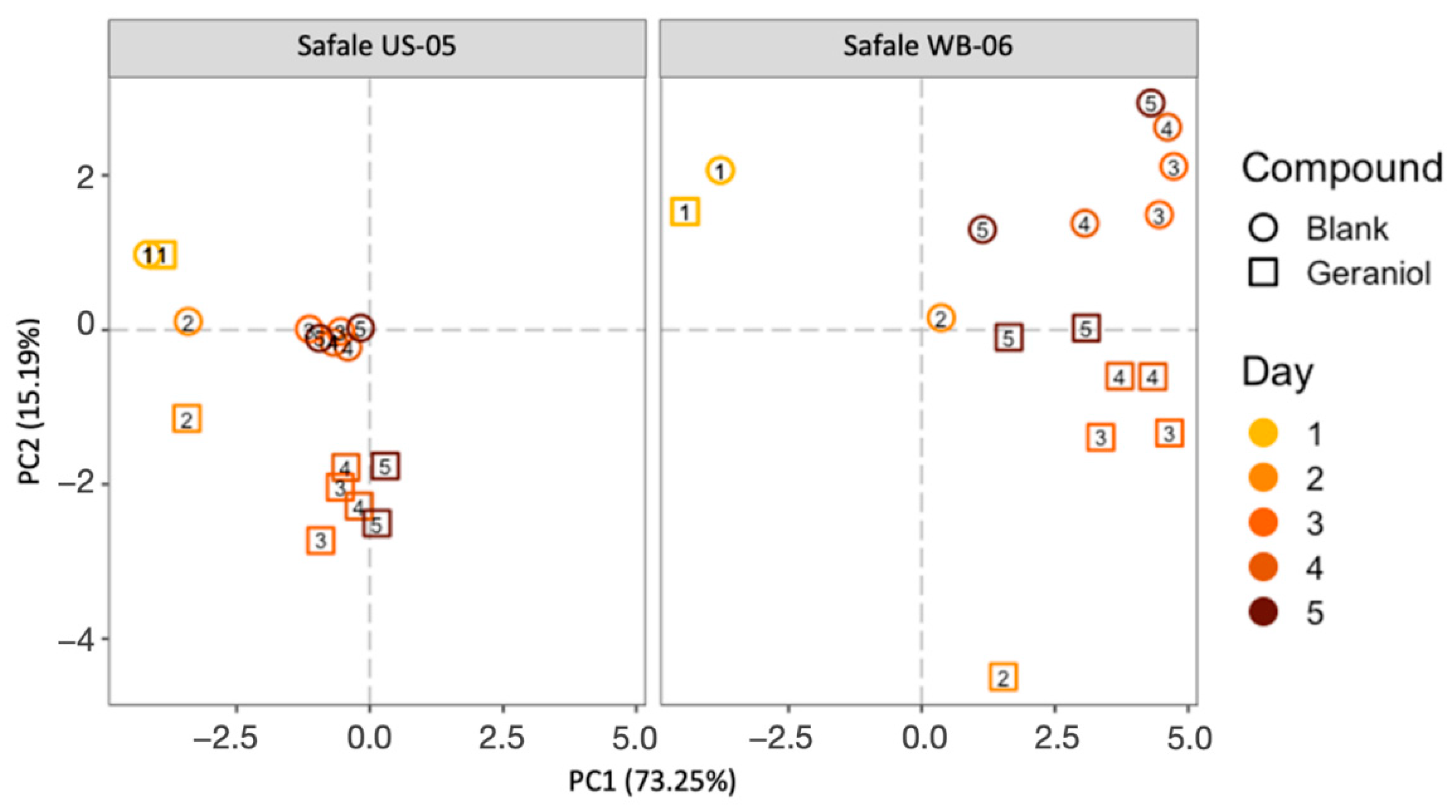
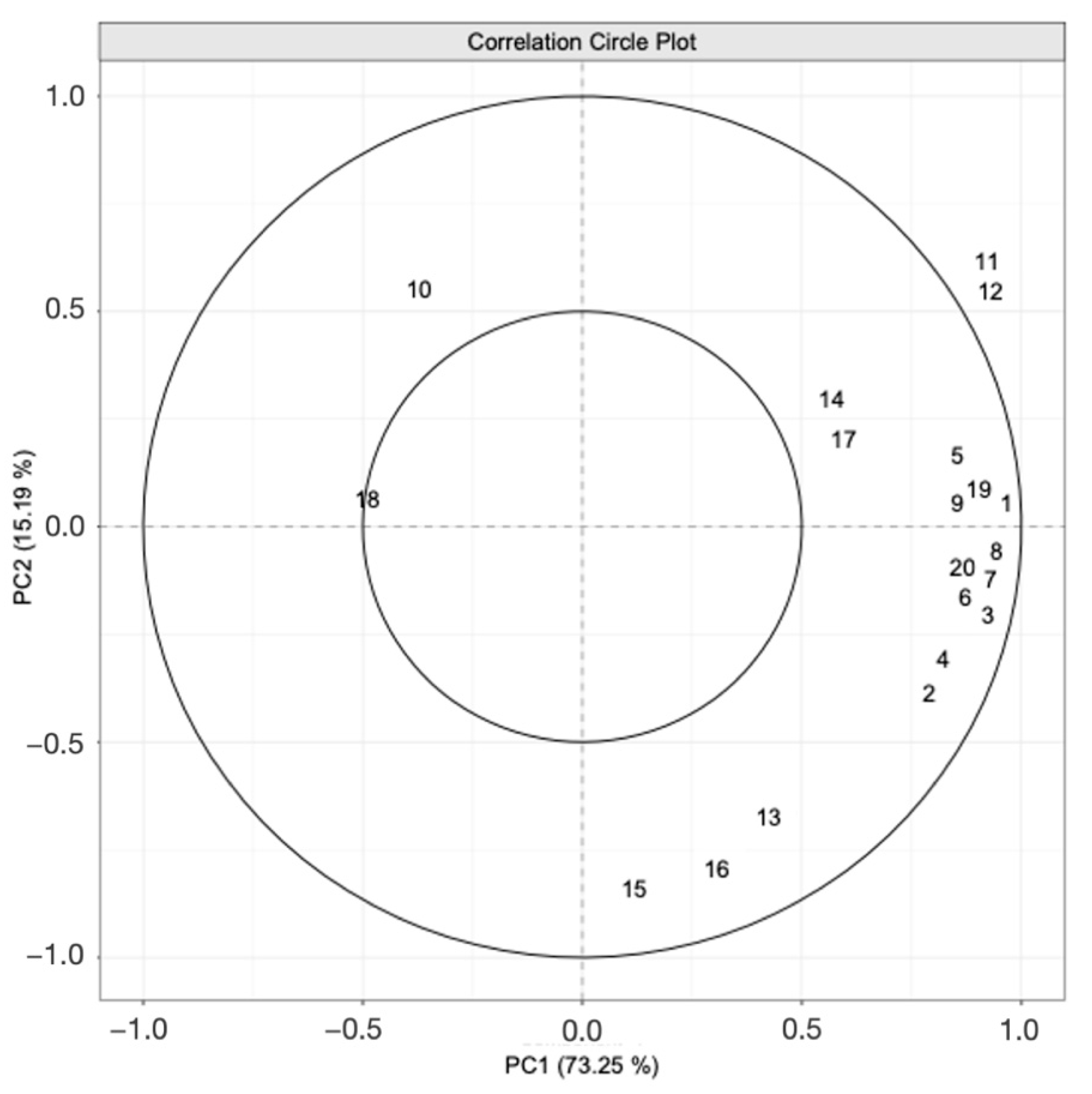
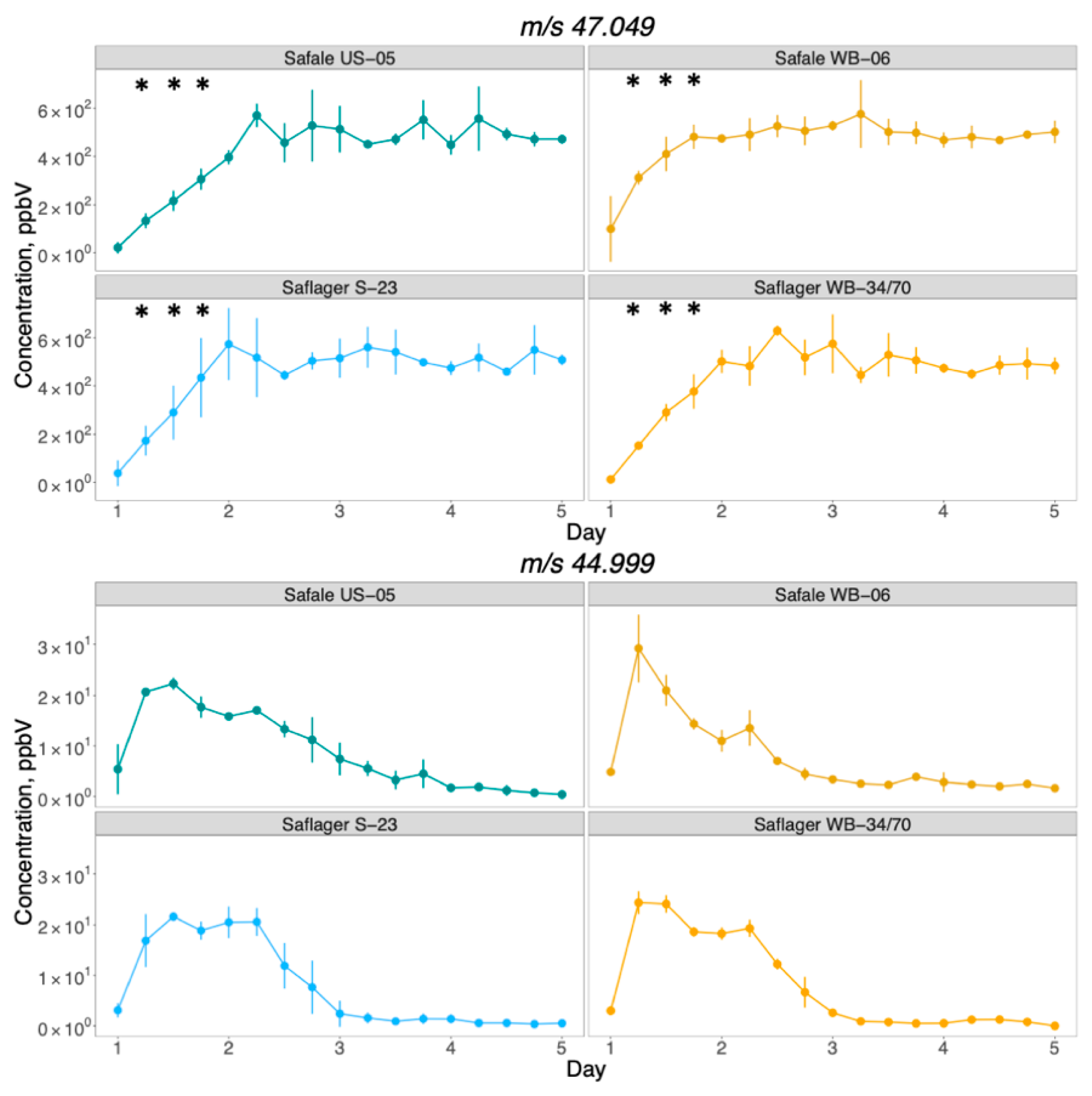
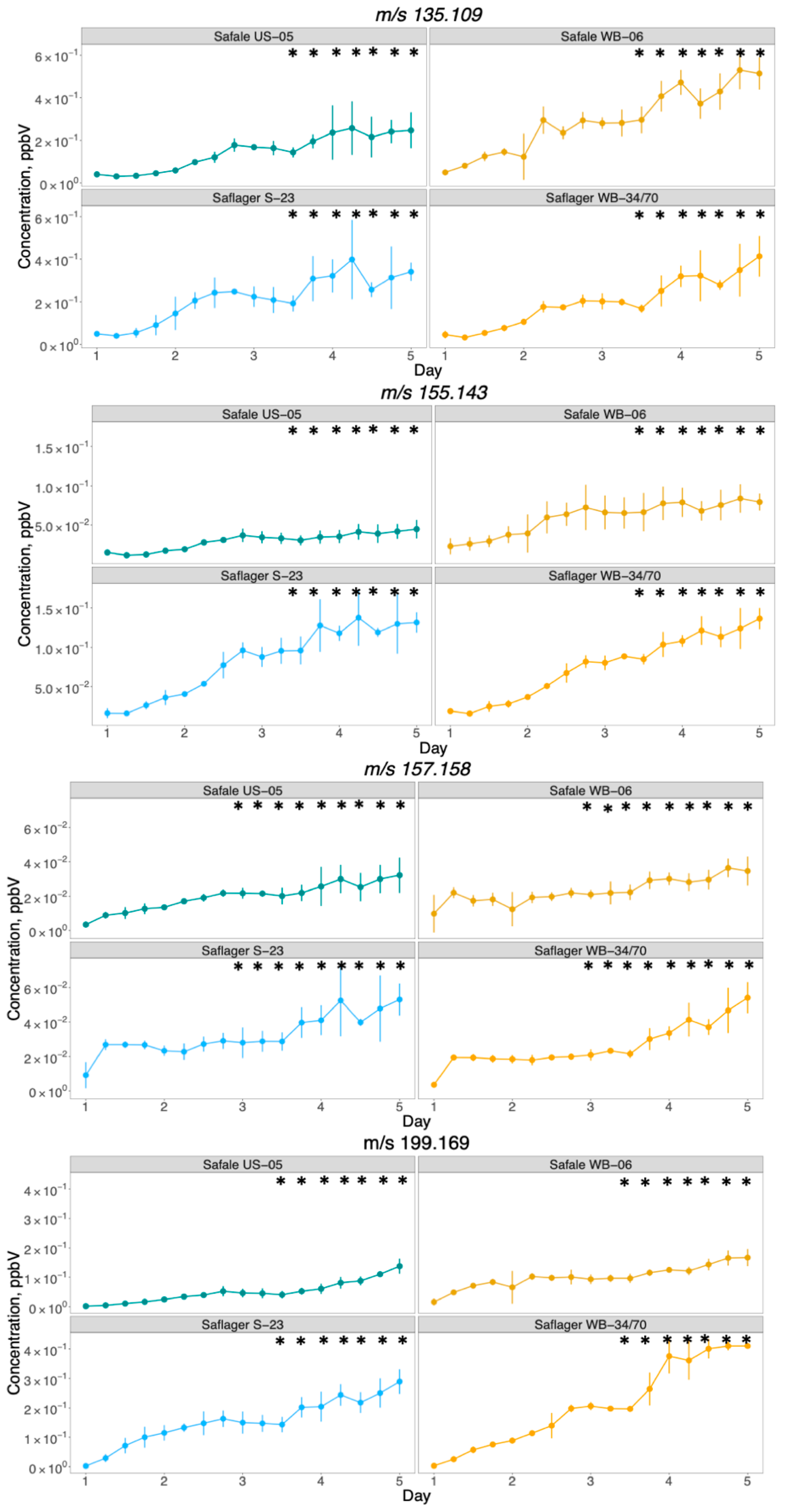
3.2. PTR-ToF-MS Results
Development of Volatiles during Fermentation
3.3. Comparison between PTR-ToF-MS and GC/MS to Monitor the Formation of Compounds throughout Beer Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tisserand, R.; Young, R. (Eds.) 2-Essential oil composition. In Essential Oil Safety, 2nd ed.; Churchill Livingstone: St. Louis, MO, USA, 2014; pp. 5–22. [Google Scholar]
- Buttery, R.; Ling, L. The chemical composition of the volatile oil of hops. Brew. Dig. 1966, 41, 71–77. [Google Scholar]
- Eyres, G.; Dufour, J.-P. 22-Hop Essential Oil: Analysis, Chemical Composition and Odor Characteristics. In Beer in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 239–254. [Google Scholar]
- Takoi, K.; Itoga, Y.; Koie, K.; Kosugi, T.; Shimase, M.; Katayama, Y.; Nakayama, Y.; Watari, J. The Contribution of Geraniol Metabolism to the Citrus Flavour of Beer: Synergy of Geraniol and β-Citronellol Under Coexistence with Excess Linalool. J. Inst. Brew. 2010, 116, 251–260. [Google Scholar] [CrossRef]
- King, A.; Dickinson, R. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- King, A.; Dickinson, R. Biotransformation of hop aroma terpenoids by ale and lager yeasts. FEMS Yeast Res. 2003, 3, 53–62. [Google Scholar] [CrossRef]
- Richter, T.M.; Silcock, P.; Algarra, A.; Eyres, G.T.; Capozzi, V.; Bremer, P.J.; Biasioli, F. Evaluation of PTR-ToF-MS as a tool to track the behavior of hop-derived compounds during the fermentation of beer. Food Res. Int. 2018, 111, 582–589. [Google Scholar] [CrossRef]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Gamero, A.; Manzanares, P.; Querol, A.; Belloch, C. Monoterpene alcohols release and bioconversion by Saccharomyces species and hybrids. Int. J. Food Microbiol. 2011, 145, 92–97. [Google Scholar] [CrossRef]
- Ugliano, M.; Genovese, A.; Moio, L. Hydrolysis of wine aroma precursors during malolactic fermentation with four commercial starter cultures of Oenococcus oeni. J. Agric. Food Chem. 2003, 51, 5073–5078. [Google Scholar] [CrossRef]
- Takoi, K.; Itoga, Y.; Takayanagi, J.; Kosugi, T.; Shioi, T.; Nakamura, T.; Watari, J. Screening of Geraniol-Rich Flavor Hop and Interesting Behavior of β-Citronellol during Fermentation under Various Hop-Addition Timings. J. Am. Soc. Brew. Chem. 2014, 72, 22–29. [Google Scholar] [CrossRef]
- Sharp, D.; Qian, Y.; Shellhammer, G.; Shellhammer, T.H. Contributions of Select Hopping Regimes to the Terpenoid Content and Hop Aroma Profile of Ale and Lager Beers. J. Am. Soc. Brew. Chem. 2017, 75, 93–100. [Google Scholar] [CrossRef]
- Sharp, D.C.; Vollmer, D.M.; Qian, Y.; Shellhammer, T.H. Examination of Glycoside Hydrolysis Methods for the Determination of Terpenyl Glycoside Contents of Different Hop Cultivars. J. Am. Soc. Brew. Chem. 2017, 75, 101–108. [Google Scholar] [CrossRef]
- Steyer, D.; Erny, C.; Claudel, P.; Riveill, G.; Karst, F.; Legras, J.-L. Genetic analysis of geraniol metabolism during fermentation. Food Microbiol. 2013, 33, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Richter, T.; Algarra Alarcon, A.; Silcock, P.; Eyres, G.; Bremer, P.; Capozzi, V.; Biasioli, F. Tracking of Hop-Derived Compounds in Beer During Fermentation with PTR-TOF-MS. In Proceedings of the 15th Weurman Flavour Research Symposium, Graz, Austria, 18–22 September 2017; Technische Universität: Graz, Austria, 2017; pp. 171–174. [Google Scholar]
- Berbegal, C.; Khomenko, I.; Russo, P.; Spano, G.; Fragasso, M.; Biasioli, F.; Capozzi, V. PTR-ToF-MS for the Online Monitoring of Alcoholic Fermentation in Wine: Assessment of VOCs Variability Associated with Different Combinations of Saccharomyces/Non-Saccharomyces as a Case-Study. Fermentation 2020, 6, 55. [Google Scholar] [CrossRef]
- Noma, Y.; Asakawa, Y. Biotransformation of monoterpenoids by microorganisms, insects, and mammals. In Handbook of Essential Oils; CRC Press: Boca Raton, FL, USA, 2020; pp. 613–767. [Google Scholar]
- Demyttenaere, J.C.R.; del Carmen Herrera, M.; De Kimpe, N. Biotransformation of geraniol, nerol and citral by sporulated surface cultures of Aspergillus niger and Penicillium sp. Phytochemistry 2000, 55, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miks, M.H.; de Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2018, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, D.C.; Steensels, J.; Shellhammer, T.H. The effect of hopping regime, cultivar and β-glucosidase activity on monoterpene alcohol concentrations in wort and beer. J. Inst. Brew. 2017, 123, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Sharp, D.C. Factors that Influence the Aroma and Monoterpene Alcohol Profile of Hopped Beer. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2016. [Google Scholar]
- Slaghenaufi, D.; Indorato, C.; Troiano, E.; Luzzini, G.; Felis, G.E.; Ugliano, M. Fate of Grape-Derived Terpenoids in Model Systems Containing Active Yeast Cells. J. Agric. Food Chem. 2020, 68, 13294–13301. [Google Scholar] [CrossRef]
- da Silva, G.C.; da Silva, A.A.S.; da Silva, L.S.N.; Godoy, R.L.d.O.; Nogueira, L.C.; Quitério, S.L.; Raices, R.S.L. Method development by GC–ECD and HS-SPME–GC–MS for beer volatile analysis. Food Chem. 2015, 167, 71–77. [Google Scholar] [CrossRef]
- Jeleń, H.H.; Majcher, M.; Dziadas, M. Microextraction techniques in the analysis of food flavor compounds: A review. Anal. Chim. Acta 2012, 738, 13–26. [Google Scholar] [CrossRef]
- Cappellin, L.; Karl, T.; Probst, M.; Ismailova, O.; Winkler, P.M.; Soukoulis, C.; Aprea, E.; Märk, T.D.; Gasperi, F.; Biasioli, F. On quantitative determination of volatile organic compound concentrations using proton transfer reaction time-of-flight mass spectrometry. Environ. Sci. Technol. 2012, 46, 2283–2290. [Google Scholar] [CrossRef]
- Farneti, B.; Khomenko, I.; Grisenti, M.; Ajelli, M.; Betta, E.; Algarra, A.A.; Cappellin, L.; Aprea, E.; Gasperi, F.; Biasioli, F. Exploring blueberry aroma complexity by chromatographic and direct-injection spectrometric techniques. Front. Plant Sci. 2017, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak, T.; Wojnowski, W.; Lubinska-Szczygeł, M.; Różańska, A.; Namieśnik, J.; Dymerski, T. PTR-MS and GC/MS as complementary techniques for analysis of volatiles: A tutorial review. Anal. Chim. Acta 2018, 1035, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Fischer, L.; Herbig, J.; Campbell-Sills, H.; Coulon, J.; Lucas, P.; Cappellin, L.; Biasioli, F. Wine analysis by FastGC proton-transfer reaction-time-of-flight-mass spectrometry. Int. J. Mass Spectrom. 2014, 369, 81–86. [Google Scholar] [CrossRef]
- Boscaini, E.; Van Ruth, S.; Biasioli, F.; Gasperi, F.; Märk, T.D. Gas chromatography-olfactometry (GC-O) and proton transfer reaction-mass spectrometry (PTR-MS) analysis of the flavor profile of grana padano, parmigiano reggiano, and grana trentino cheeses. J. Agric. Food Chem. 2003, 51, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Blasioli, S.; Biondi, E.; Samudrala, D.; Spinelli, F.; Cellini, A.; Bertaccini, A.; Cristescu, S.M.; Braschi, I. Identification of volatile markers in potato brown rot and ring rot by combined GC/MS and PTR-MS techniques: Study on in vitro and in vivo samples. J. Agric. Food Chem. 2014, 62, 337–347. [Google Scholar] [CrossRef]
- van Ruth, S.M.; Floris, V.; Fayoux, S. Characterisation of the volatile profiles of infant formulas by proton transfer reaction-mass spectrometry and gas chromatography–mass spectrometry. Food Chem. 2006, 98, 343–350. [Google Scholar] [CrossRef]
- Pollien, P.; Lindinger, C.; Yeretzian, C.; Blank, I. Proton transfer reaction mass spectrometry, a tool for on-line monitoring of acrylamide formation in the headspace of Maillard reaction systems and processed food. Anal. Chem. 2003, 75, 5488–5494. [Google Scholar] [CrossRef]
- Marconi, O.; Rossi, S.; Galgano, F.; Sileoni, V.; Perretti, G. Influence of yeast strain, priming solution and temperature on beer bottle conditioning. J. Sci. Food Agric. 2016, 96, 4106–4115. [Google Scholar] [CrossRef]
- Capozzi, V.; Yener, S.; Khomenko, I.; Farneti, B.; Cappellin, L.; Gasperi, F.; Scampicchio, M.; Biasioli, F. PTR-ToF-MS coupled with an automated sampling system and tailored data analysis for food studies: Bioprocess monitoring, screening and nose-space analysis. JoVE 2017, 123, e54075. [Google Scholar]
- Khomenko, I.; Stefanini, I.; Cappellin, L.; Cappelletti, V.; Franceschi, P.; Cavalieri, D.; Märk, T.D.; Biasioli, F. Non-invasive real time monitoring of yeast volatilome by PTR-ToF-MS. Metabolomics 2017, 13, 118. [Google Scholar] [CrossRef] [Green Version]
- Cappellin, L.; Biasioli, F.; Granitto, P.M.; Schuhfried, E.; Soukoulis, C.; Costa, F.; Märk, T.D.; Gasperi, F. On data analysis in PTR-TOF-MS: From raw spectra to data mining. Sens. Actuators B Chem. 2011, 155, 183–190. [Google Scholar] [CrossRef]
- Cappellin, L.; Biasioli, F.; Fabris, A.; Schuhfried, E.; Soukoulis, C.; Märk, T.D.; Gasperi, F. Improved mass accuracy in PTR-TOF-MS: Another step towards better compound identification in PTR-MS. Int. J. Mass Spectrom. 2010, 290, 60–63. [Google Scholar] [CrossRef]
- Lindinger, W.; Hansel, A.; Jordan, A. On-line monitoring of volatile organic compounds at pptv levels by means of proton-transfer-reaction mass spectrometry (PTR-MS) medical applications, food control and environmental research. Int. J. Mass Spectrom. Ion Process. 1998, 173, 191–241. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.M.P.L.V.O.; Jorge, K.; Nogueira, L.C.; Silva, F.; Trugo, L.C. Effects of the Combination of Hydrophobic Polypeptides, Iso-α Acids, and Malto-oligosaccharides on Beer Foam Stability. J. Agric. Food Chem. 2005, 53, 4976–4981. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. Whole-genome comparison reveals novel genetic elements that characterize the genome of industrial strains of Saccharomyces cerevisiae. PLoS Genet. 2011, 7, e1001287. [Google Scholar] [CrossRef] [Green Version]
- Holzinger, R.; Sandoval-Soto, L.; Rottenberger, S.; Crutzen, P.; Kesselmeier, J. Emissions of volatile organic compounds from Quercus ilex L. measured by Proton Transfer Reaction Mass Spectrometry under different environmental conditions. J. Geophys. Res. Atmos. 2000, 105, 20573–20579. [Google Scholar] [CrossRef]
- Warneke, C.; Karl, T.; Judmaier, H.; Hansel, A.; Jordan, A.; Lindinger, W.; Crutzen, P.J. Acetone, methanol, and other partially oxidized volatile organic emissions from dead plant matter by abiological processes: Significance for atmospheric HOx chemistry. Glob. Biogeochem. Cycles 1999, 13, 9–17. [Google Scholar] [CrossRef]
- Tani, A.; Hayward, S.; Hewitt, C. Measurement of monoterpenes and related compounds by proton transfer reaction-mass spectrometry (PTR-MS). Int. J. Mass Spectrom. 2003, 223, 561–578. [Google Scholar] [CrossRef]
- Soukoulis, C.; Cappellin, L.; Aprea, E.; Costa, F.; Viola, R.; Märk, T.D.; Gasperi, F.; Biasioli, F. PTR-ToF-MS, a novel, rapid, high sensitivity and non-invasive tool to monitor volatile compound release during fruit post-harvest storage: The case study of apple ripening. Food Bioprocess Technol. 2013, 6, 2831–2843. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amelynck, C.; Mees, B.; Schoon, N.; Bultinck, P. FA-SIFT study of the reactions of H3O+·(H2O)n (n = 0, 1, 2), NO+ and O2+ with the terpenoid aldehydes citral, citronellal and myrtenal and their alcohol analogues. Int. J. Mass Spectrom. 2015, 379, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Haslbeck, K.; Bub, S.; von Kamp, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Coelhan, M. The influence of brewing yeast strains on monoterpene alcohols and esters contributing to the citrus flavour of beer. J. Inst. Brew. 2018, 124, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Ito, M. Bioconversion of essential oil components of Perilla frutescens by Saccharomyces cerevisiae. J. Nat. Med. 2020, 74, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression Levels of the Yeast Alcohol Acetyltransferase Genes ATF1, Lg-ATF1, and ATF2 Control the Formation of a Broad Range of Volatile Esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.B.; Dufour, J.P. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Tuszyński, T. The effect of wort aeration on fermentation, maturation and volatile components of beer produced on an industrial scale. J. Inst. Brew. 2017, 123, 31–38. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Species | Yeast Strain | SPME-GC/MS | Measurement Frequency (h) | PTR-ToF-MS | Measurement Frequency (h) |
|---|---|---|---|---|---|
| S. cerevisiae | SafAle US-05 | ✓ | 24 | ✓ | 6 |
| S. cerevisiae var. Diastaticus | SafAleWB-06 | ✓ | 24 | ✓ | 6 |
| S. pastorianus | SafLager W-34/70 | - | - | ✓ | 6 |
| S. pastorianus | SafLager S-23 | - | - | ✓ | 6 |
| Theoretical m/z | Measured m/z | Sum Formula | Chemical Class | Tentative Identification |
|---|---|---|---|---|
| 28.0062 | 28.006 | C2H5+ | Alcohols | Ethanol Fragment |
| 33.0339 | 33.034 | CH4OH+ | Alcohols | Methanol |
| 48.0529 | 48.053 | C2H5OH+ | Alcohols | Ethanol (isotopologue) |
| 59.0491 | 59.049 | C3H6OH+ | Aldehydes/ketones | Propanol/acetone |
| 62.0317 | 62.031 | C2H4O2H+ | Esters and acids | Acetic acid |
| 64.0292 | 64.029 | C2H6SH+ | Sulphur compounds | Dimethylsulfide |
| 69.0697 | 69.069 | C5H8H+ | Terpene | Terpene fragment |
| 76.047 | 75.043 | C3H6O2H+ | Esters and acids | Propionic acid |
| 81.0699 | 81.07 | C6H8H+ | Terpene | Terpene fragment |
| 83.0783 | 83.084 | C6H10H+ | Terpene | Terpene fragment |
| 85.0654 | 85.064 | C5H8OH+ | Aldehydes/Ketones | Pentanal/pentenone |
| 87.0439 | 87.043 | C4H6O2H+ | Ketones | Butanedione |
| 87.0803 | 87.08 | C5H10OH+ | Alcohols | Pentanol |
| 94.0952 | 93.068 | C7H7+ | Terpene | Terpene fragment |
| 95.0492 | 95.046 | C6H6OH+ | Phenols | Phenol |
| 95.096 | 95.09 | C7H10H+ | Terpenes | Terpene fragment |
| 97.0284 | 97.027 | C5H4O2H+ | Aldehydes | Furfural |
| 97.0642 | 97.057 | C6H8OH+ | Aldehydes/Furans | Hexadienal/ethylfuran |
| 99.0802 | 99.079 | C6H10OH+ | Aldehydes | Hexenal/methylpentenone |
| 101.0951 | 101.091 | C6H12OH+ | Alcohols | Hexanol |
| 103.0749 | 103.074 | C5H10O2H+ | Esters and acids | Methylbutanoic acid |
| 107.0705 | 107.07 | C7H6OH+ | Aldehydes | Benzaldehyde |
| 107.1071 | 107.102 | C8H10H+ | Aromatic hydrocarbons | Xylene/ethylbenzene |
| 109.0712 | 109.059 | C7H8OH+ | Phenols | Benzyl alcohol (cresol) |
| 111.0463 | 111.042 | C6H6O2H+ | Furans | Acetyl furan |
| 111.0804 | 111.076 | C7H10OH+ | Aldehydes | Heptadienal |
| 113.0965 | 113.096 | C7H12OH+ | Aldehydes | Heptanal |
| 115.1109 | 115.111 | C7H14OH+ | Ketones | Heptanone |
| 121.0691 | 121.067 | C8H8OH+ | Aldehydes | Methylbenzaldehyde-coumaran |
| 127.1117 | 127.112 | C8H14OH+ | Ketones | Octenone/methylheptenone |
| 129.0911 | 129.091 | C7H12O2H+ | Esters and acids | Hexenyl formate |
| 129.1272 | 129.125 | C8H16OH+ | Ketones | Octanone/Dimethylcyclohexanol |
| 131.1062 | 131.107 | C7H14O2H+ | Esters and acids | Heptanoic acid/hexyl formate |
| 135.1032 | 135.109 | C10H14H+ | Aromatic hydrocarbons | Methylpropylbenzene |
| 136.1073 | 136.112 | C9H13NH+ | Heterocyclic compounds | Butyl-pyridine/ethyl-propylpyridine |
| 137.132 | 137.133 | C10H16H+ | Terpenes | Various monoterpenes |
| 141.1357 | 141.127 | C9H16OH+ | Aldehydes | Nonanal |
| 143.1443 | 143.148 | C9H18OH+ | Ketones/Aldehydes | Nonanone/nonanal |
| 151.1108 | 151.112 | C10H14OH+ | Terpenes | Carvacrol/safranal |
| 153.0615 | 153.063 | C8H8O3H+ | Aldehydes | Vanillin, methyl salicylate |
| 153.1234 | 153.126 | C10H16OH+ | Aldehydes | Citral |
| 155.1424 | 155.143 | C10H18OH+ | Alcohols | Linalool/geraniol/a-terpineol/nerol |
| 157.1576 | 157.158 | C10H20OH+ | Alcohols | Citronellol/dihydrolinalool |
| 171.1373 | 171.137 | C10H18O2H+ | Terpenes | Linalool oxide/Citronellic acid |
| 199.1677 | 199.169 | C12H23O2H+ | Terpenes | Citronellyl acetate |
| 201.1819 | 201.184 | C12H24O2H+ | Terpenes | Dihydrocitronellyl acetate |
| 205.1878 | 205.200 | C12H23O2H+ | Terpenes | Humulene |
| MW (g/mol) | m/z | 81 | 83 | 93 | 95 | 135 | 137 | 139 | 155 | 157 | 199 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Geraniol | 154.25 | C10H18O | 82.03 | 100 | 51.25 | 46.06 | 0.27 | |||||
| Citronellol | 156.27 | C10H20O | 78.52 | 100 | 64.75 | 26.60 | 20.32 | |||||
| Geranyl acetate | 196.29 | C12H22O2 | 100 | 10.45 | 1.6 | 42.53 | ||||||
| Citronellyl acetate | 198.30 | C12H22O2 | 40.47 | 100 | 11.10 | 41.32 | 25.51 |
| Number | Compound | Formula | CAS |
|---|---|---|---|
| 1 | Ethyl Acetate | C4H8O2 | 141-78-6 |
| 2 | Ethanol | C2H6O | 200-578-6 |
| 3 | Ethyl propanoate | C5H10O2 | 105-37-3 |
| 4 | Ethyl butanoate | C6H12O2 | 105-54-4 |
| 5 | Isobutyl alcohol | C4H10O | 78-83-1 |
| 6 | Isoamyl acetate | C7H14O2 | 123-92-2 |
| 7 | Isoamyl alcohol | C5H12O | 123-51-3 |
| 8 | Ethyl hexanoate | C8H16O2 | 123-66-0 |
| 9 | Ethyl octanoate | C10H20O2 | 106-32-1 |
| 10 | Acetic acid | CH3COOH | 64-19-7 |
| 11 | Ethyl decanoate | C12H24O2 | 110-38-3 |
| 12 | Isoamyl octanoate | C13H26O2 | 2035-99-6 |
| 13 | Citronellyl acetate | C12H22O2 | 150-84-5 |
| 14 | Ethyl 9-decenoate | C12H22O2 | 67233-91-4 |
| 15 | Geranyl acetate | C12H20O2 | 105-87-3 |
| 16 | Citronellol | C10H20O | 106-22-9 |
| 17 | Ethyl dodecanoate | C14H28O2 | 106-33-2 |
| 18 | Geraniol | C10H18O | 106-24-1 |
| 19 | Phenylethyl alcohol | C8H10O | 60-12-8 |
| 20 | Octanoic acid | C8H16O2 | 124-07-2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, R.; Khomenko, I.; Eyres, G.T.; Bremer, P.; Silcock, P.; Betta, E.; Biasioli, F. Investigation of Geraniol Biotransformation by Commercial Saccharomyces Yeast Strains by Two Headspace Techniques: Solid-Phase Microextraction Gas Chromatography/Mass Spectrometry (SPME-GC/MS) and Proton Transfer Reaction-Time of Flight-Mass Spectrometry (PTR-ToF-MS). Fermentation 2023, 9, 294. https://doi.org/10.3390/fermentation9030294
Roberts R, Khomenko I, Eyres GT, Bremer P, Silcock P, Betta E, Biasioli F. Investigation of Geraniol Biotransformation by Commercial Saccharomyces Yeast Strains by Two Headspace Techniques: Solid-Phase Microextraction Gas Chromatography/Mass Spectrometry (SPME-GC/MS) and Proton Transfer Reaction-Time of Flight-Mass Spectrometry (PTR-ToF-MS). Fermentation. 2023; 9(3):294. https://doi.org/10.3390/fermentation9030294
Chicago/Turabian StyleRoberts, Rebecca, Iuliia Khomenko, Graham T. Eyres, Phil Bremer, Patrick Silcock, Emanuela Betta, and Franco Biasioli. 2023. "Investigation of Geraniol Biotransformation by Commercial Saccharomyces Yeast Strains by Two Headspace Techniques: Solid-Phase Microextraction Gas Chromatography/Mass Spectrometry (SPME-GC/MS) and Proton Transfer Reaction-Time of Flight-Mass Spectrometry (PTR-ToF-MS)" Fermentation 9, no. 3: 294. https://doi.org/10.3390/fermentation9030294
APA StyleRoberts, R., Khomenko, I., Eyres, G. T., Bremer, P., Silcock, P., Betta, E., & Biasioli, F. (2023). Investigation of Geraniol Biotransformation by Commercial Saccharomyces Yeast Strains by Two Headspace Techniques: Solid-Phase Microextraction Gas Chromatography/Mass Spectrometry (SPME-GC/MS) and Proton Transfer Reaction-Time of Flight-Mass Spectrometry (PTR-ToF-MS). Fermentation, 9(3), 294. https://doi.org/10.3390/fermentation9030294