

Fermented Wheat Bran Polysaccharides Improved Intestinal Health of Zebrafish in Terms of Intestinal Motility and Barrier Function
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Wheat Bran Polysaccharides
2.2. Experiment 1: Effect of FWBP on Promoting Intestinal Peristalsis and Alleviating Constipation in Zebrafish Larvae
2.2.1. Intestinal Peristalsis-Promoting Effect in Zebrafish Larvae
2.2.2. Alleviating the Constipating Effect of FWBP in Zebrafish Larvae
2.3. Experiment 2: Effects of FWBP on the Immune Activity, Intestinal Morphology and Gut MicroBiota of Zebrafish
2.3.1. Animals and Experimental Diets
2.3.2. Histological of the Intestines
2.3.3. RT-PCR Analysis
2.3.4. 16S rRNA Gene Sequencing Analyses of Zebrafish Gut Microbiota
2.4. Statistical Analysis
3. Results
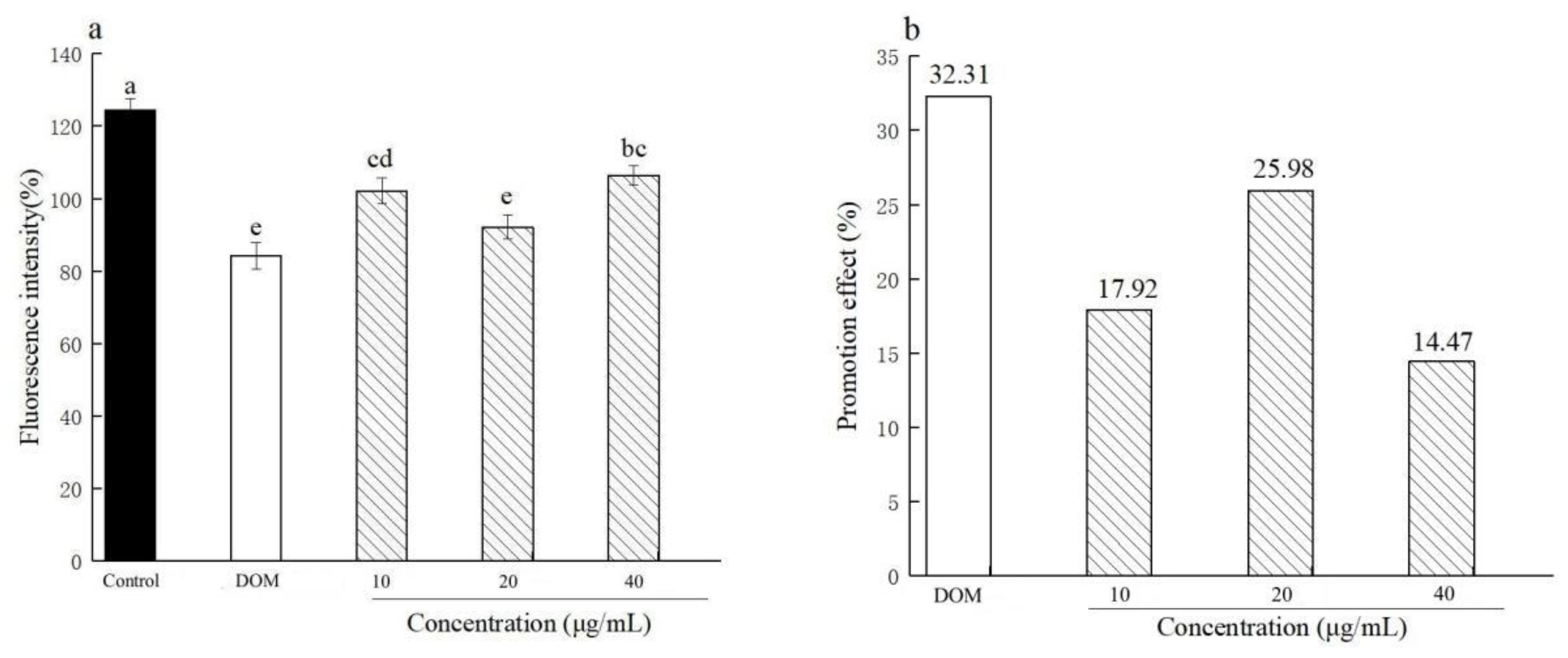
3.1. Experiment 1: Effect of FWBP on Promoting Intestinal Peristalsis and Alleviating Constipation in Zebrafish
3.2. Experiment 2 Effects of FWBP on Immune Activity, Intestinal Morphology and Gut Microbiota of Zebrafish
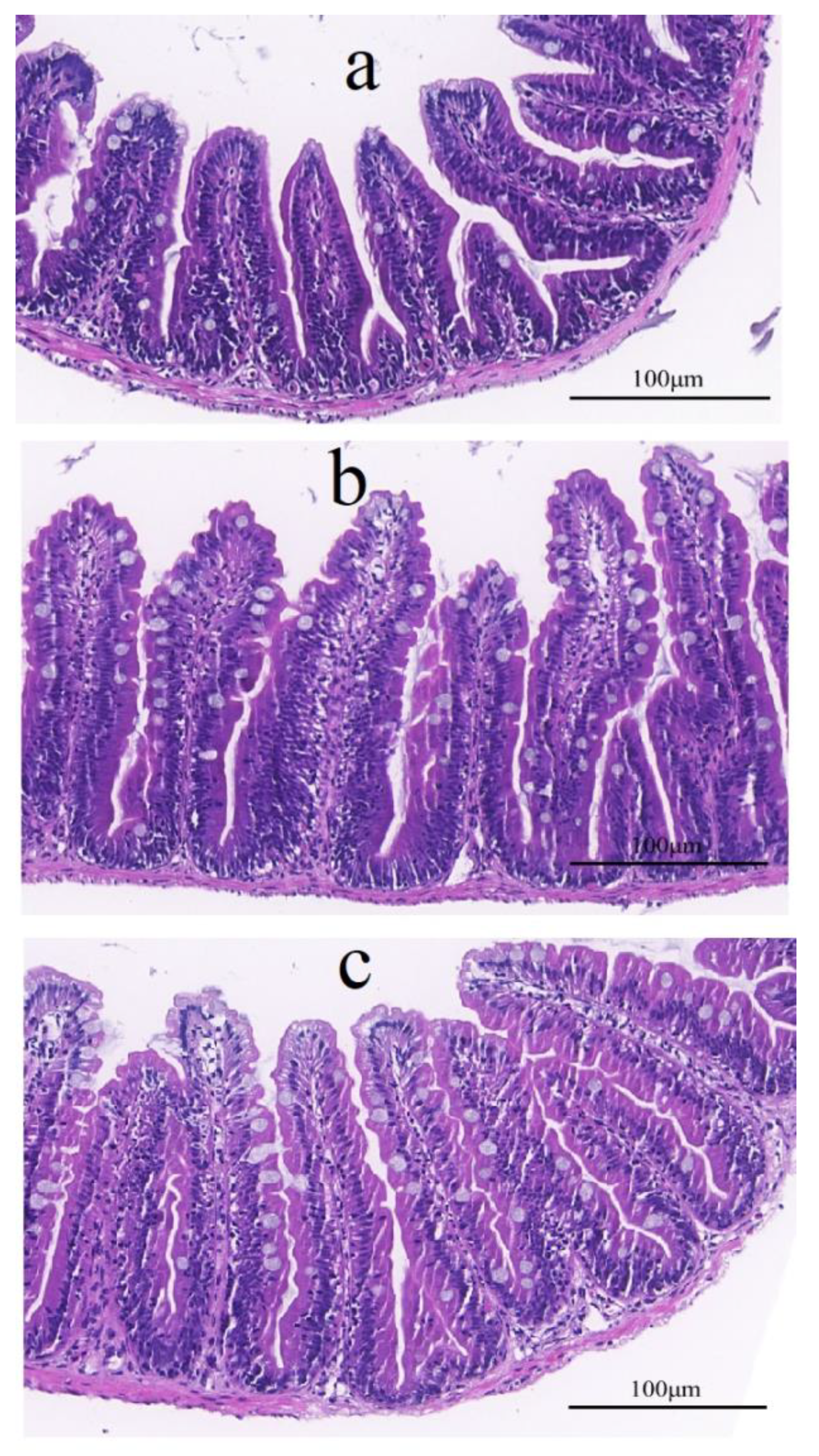
3.2.1. Effect of FWBP on Intestinal Morphology
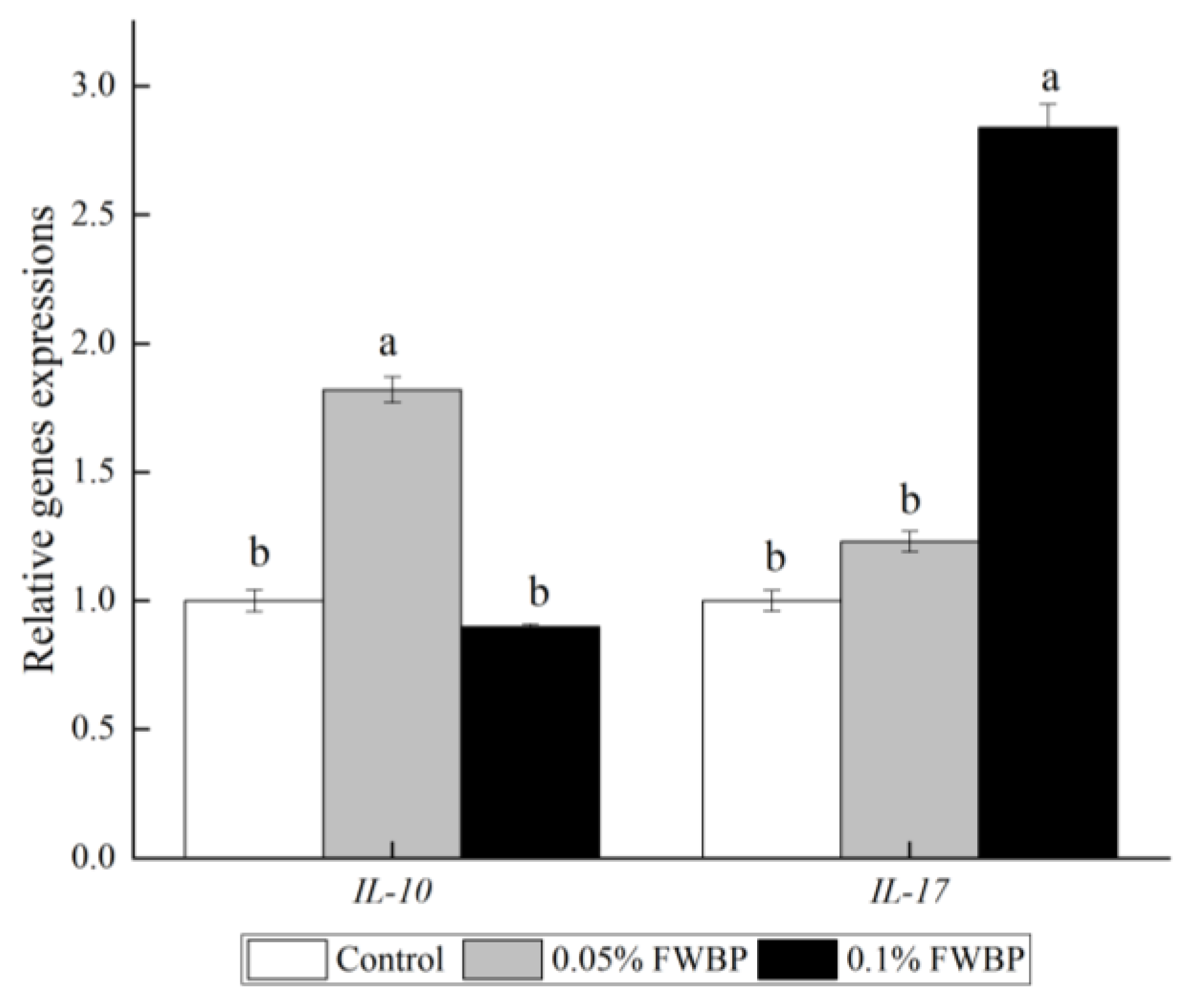
3.2.2. Expression of Intestinal Inflammation-Related Genes and Intestinal Mucosal Barrier-Related Genes
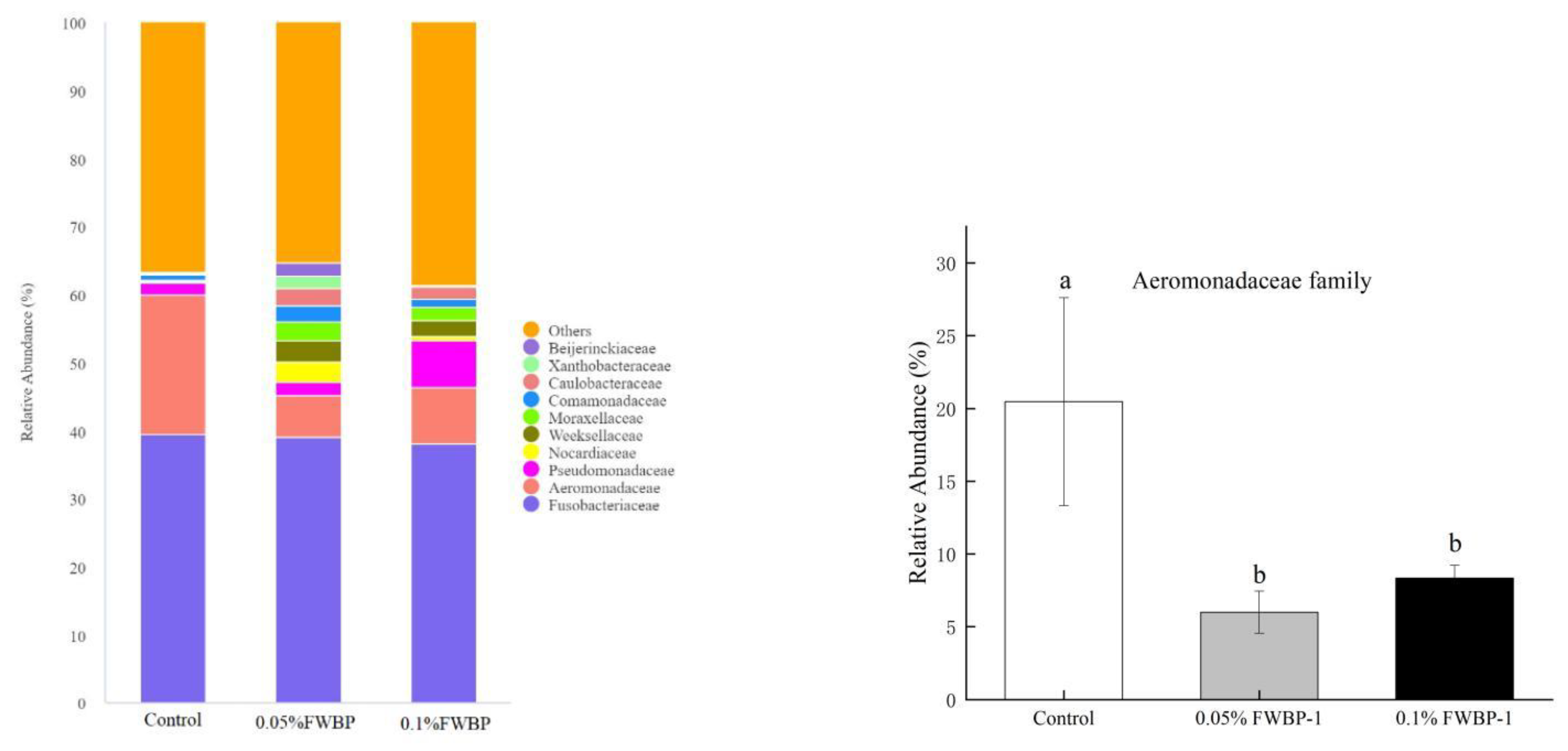
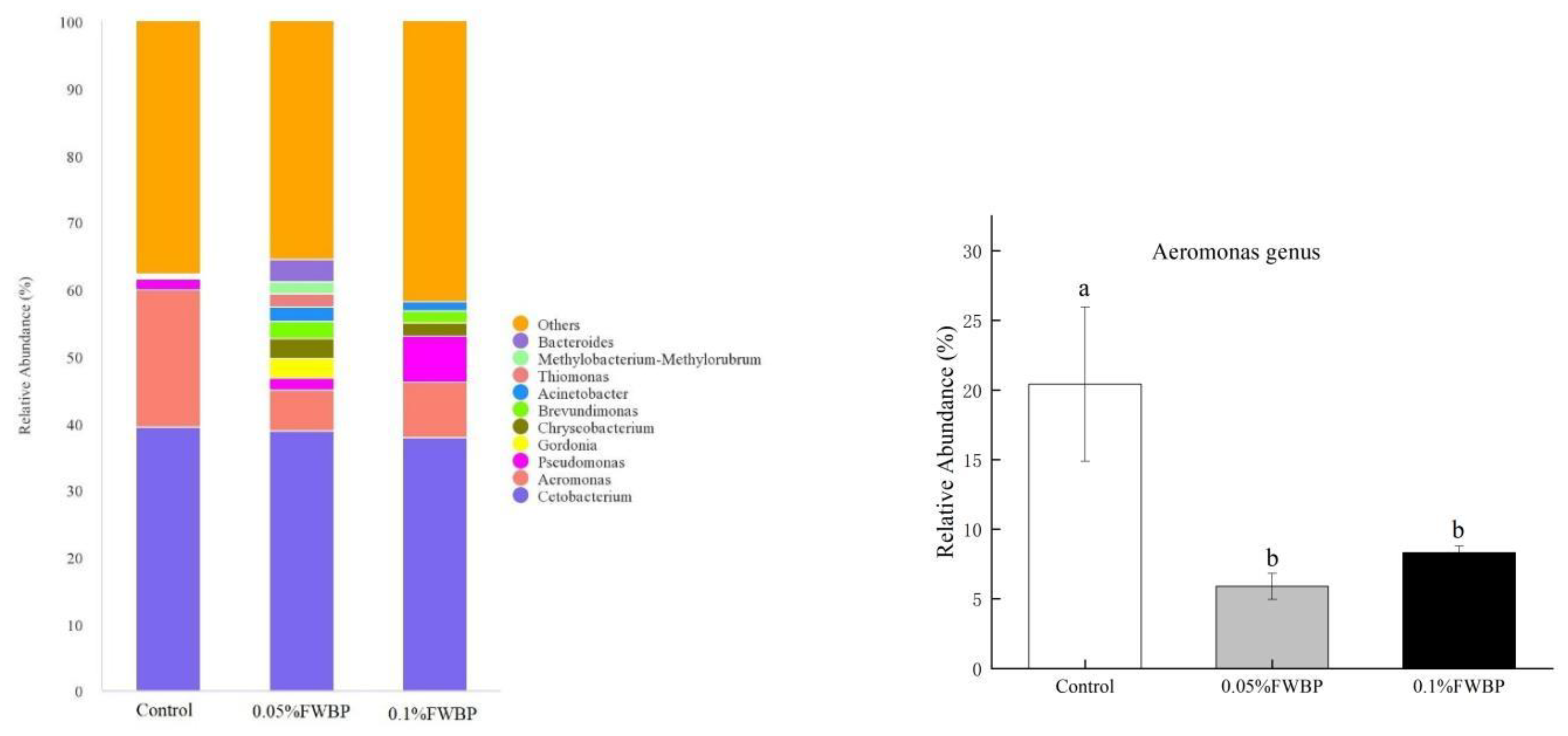
3.2.3. Effect of FWBP on Gut Microbiota
Diversity of Gut Microbiota
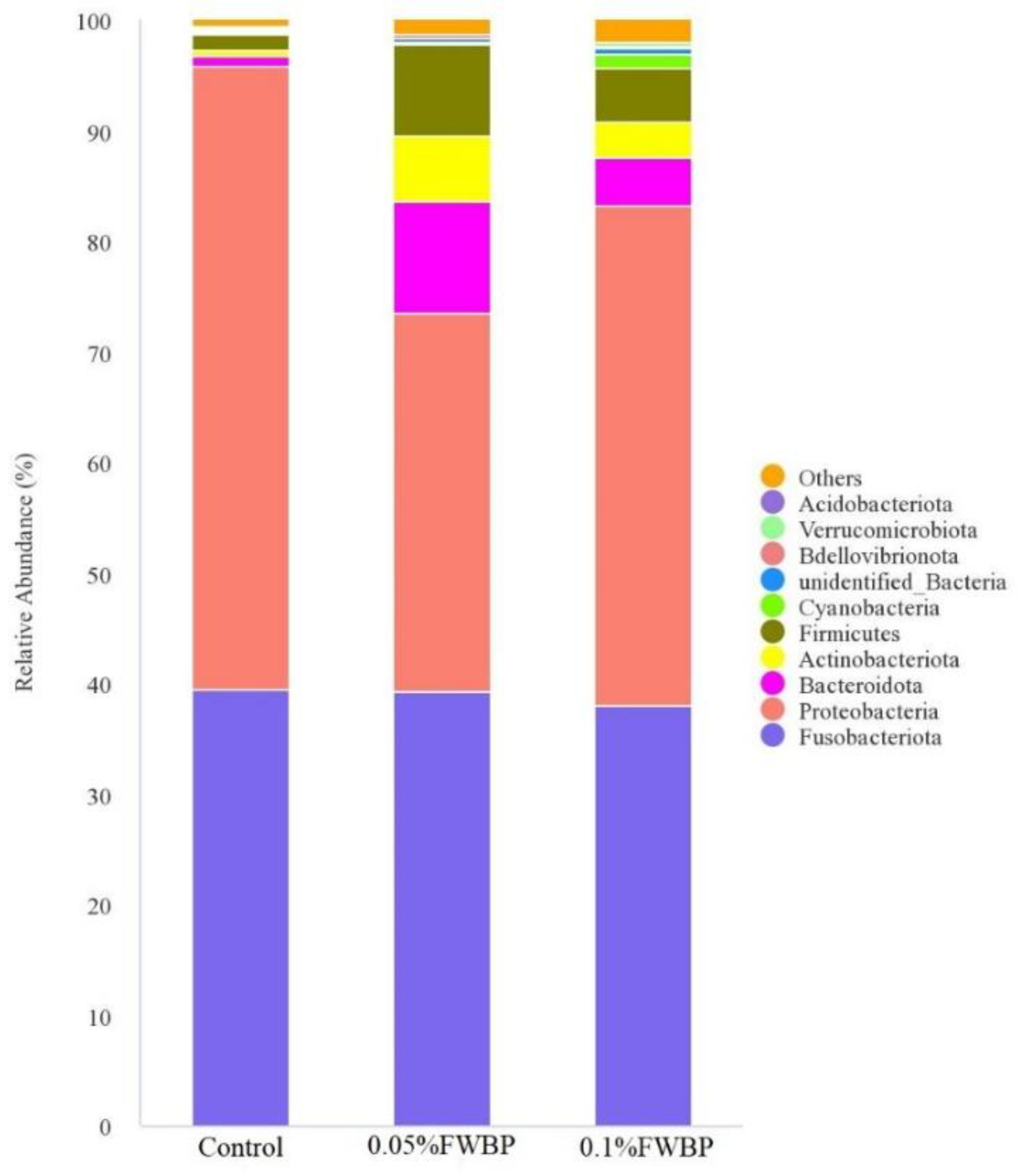
Composition of Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hao, Y.; Wang, X.; Yuan, S.; Wang, Y.; Liao, X.; Zhong, M.; He, Q.; Shen, H.; Liao, W.; Shen, J. Flammulina velutipes polysaccharide improves C57BL/6 mice gut health through regulation of intestine microbial metabolic activity. Int. J. Biol. Macromol. 2020, 167, 1308–1318. [Google Scholar] [CrossRef]
- Shi, H.T.; Zhao, S.Z.; Wang, K.L.; Fan, M.X.; Han, Y.Q.; Wang, H.L. Effects of dietary Astragalus Membranaceus supplementation on growth performance, and intestinal morphology, microbiota and metabolism in common carp (Cyprinus carpio). Aquac. Rep. 2022, 22, 100955. [Google Scholar] [CrossRef]
- Zhou, J.; Guo, S.-Y.; Zhang, Y.; Li, C.-Q. Human prokinetic drugs promote gastrointestinal motility in zebrafish. Neurogastroenterol. Motil. 2014, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.J.; Li, M.Z.; Gao, H.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Zhang, Y.L.; Xie, M.Y.; Nie, S.P. Polysaccharides from fermented Momordica charantia L. with Lactobacillus plantarum NCU116 ameliorate metabolic disorders and gut microbiota change in obese rats. Food Funct. 2021, 12, 2617–2630. [Google Scholar] [CrossRef]
- Do, M.H.; Seo, Y.S.; Park, H.-Y. Polysaccharides: Bowel health and gut microbiota. Crit. Rev. Food Sci. Nutr. 2020, 61, 1212–1224. [Google Scholar] [CrossRef]
- Jiang, H.; Dong, J.; Jiang, S.; Liang, Q.; Zhang, Y.; Liu, Z.; Ma, C.; Wang, J.; Kang, W. Effect of Durio zibethinus rind polysaccharide on functional constipation and intestinal microbiota in rats. Food Res. Int. 2020, 136, 109316. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Wang, G.; You, L.; Zhang, L.; Ren, H.; Hu, W.; Qiang, Q.; Wang, X.; Ji, L.; Gu, Z.; et al. Polysaccharide from wheat bran induces cytokine expression via the toll-like receptor 4-mediated p38 MAPK signaling pathway and prevents cyclophosphamide-induced immunosuppression in mice. Food Nutr. Res. 2017, 61, 1344523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendis, M.; Simsek, S. Arabinoxylans and human health. Food Hydrocoll. 2014, 42, 239–243. [Google Scholar] [CrossRef]
- Hromádková, Z.; Paulsen, B.S.; Polovka, M.; Košťálová, Z.; Ebringerová, A. Structural features of two heteroxylan polysaccharide fractions from wheat bran with anti-complementary and antioxidant activities. Carbohydr. Polym. 2013, 93, 22–30. [Google Scholar] [CrossRef]
- Lv, Q.Q.; Cao, J.J.; Liu, R.; Chen, H.Q. Structural characterization, α-amylase and α-glucosidase inhibitory activities of polysaccharides from wheat bran. Food Chem. 2021, 341 Pt 1, 128218. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, R.; Wang, Y.; An, X.; Liu, N.; Song, M.; Yang, Y.; Yin, N.; Qi, J. Characterization and antioxidant activity of wheat bran polysaccharides modified by Saccharomyces cerevisiae and Bacillus subtilis fermentation. J. Cereal Sci. 2020, 97, 103157. [Google Scholar] [CrossRef]
- Shi, J.X. Research on the Microbial Fermentation of Weat Bran Polysaccharide and the Antioxidant Activity of Crude Polysaccharide. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2017. [Google Scholar]
- Chen, Q.; Wang, Y.; Yin, N.; Wang, R.; Zheng, Y.; Yang, Y.; An, X.; Qi, J. Polysaccharides from fermented wheat bran enhanced the growth performance of zebrafish (Danio rerio) through improving gut microflora and antioxidant status. Aquac. Rep. 2022, 25, 101188. [Google Scholar] [CrossRef]
- Caro, M.; Iturria, I.; Martinez-Santos, M.; Pardo, M.A.; Rainieri, S.; Tueros, I.; Navarro, V. Zebrafish dives into food research: Effectiveness assessment of bioactive compounds. Food Funct. 2016, 7, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.N.; Zhang, J.L.; Ren, H.T.; Zhou, B.H.; Wu, Q.J.; Sun, P. Effect of tributyltin on antioxidant ability and immune responses of zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalili, M.; Tabarraei, A.; Van Doan, H.; Spanò, N.; Faggio, C. Mucosal immune parameters, immune and antioxidant defence related genes expression and growth performance of zebrafish (Danio rerio) fed on Gracilaria gracilis powder. Fish Shellfish. Immunol. 2018, 83, 232–237. [Google Scholar] [CrossRef]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ikeda-Ohtsubo, W.; Nadal, A.L.; Zaccaria, E.; Iha, M.; Kitazawa, H.; Kleerebezem, M.; Brugman, S. Intestinal Microbiota and Immune Modulation in Zebrafish by Fucoidan From Okinawa Mozuku (Cladosiphon okamuranus). Front. Nutr. 2020, 7, 67. [Google Scholar] [CrossRef]
- Song, X.J.; Feng, Z.F.; Tan, J.B.; Wang, Z.Y.; Zhu, W. Dietary administration of Pleurotus ostreatus polysaccharides (POPS) modulates the non-specific immune response and gut microbiota diversity of Apostichopus japonicus. Aquac. Rep. 2021, 19, 100578. [Google Scholar] [CrossRef]
- Maes, C.; Delcour, J.A. Structural characterisation of water-extractable and water-unextractable arabinoxylans in wheat bran. J. Cereal. Sci. 2002, 35, 315–326. [Google Scholar] [CrossRef]
- Hussain, A.; Bose, S.; Wang, J.-H.; Yadav, M.K.; Mahajan, G.B.; Kim, H. Fermentation, a feasible strategy for enhancing bioactivity of herbal medicines. Food Res. Int. 2016, 81, 1–16. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Wang, R.; Chen, Q.; Guo, J.; Feng, X.; Zheng, X.; An, X.; Qi, J. Enzyme-assisted extraction of fermented wheat bran polysaccharides and it biological activity analysis. Nat. Prod. Res. Dev. 2019, 31, 155–162. [Google Scholar]
- Xia, C.; Ai, Q.; Xiang, N.; Zhan, H.; Liu, X.; Fang, B. The Function of Fermented Wheat Bran and Its Application in Animal Feed. Chin. J. Anim. Sci. 2017, 53, 17–21. [Google Scholar]
- Wang Yuan, S.J.; Yuanxiao, D.; Qianru, Z.; Ziqi, M.; Xuan, L.; Xiaoping, A.; Jingwei, Q. Optimization of Fermentation Conditions for Improved Production of Polysaccharides from Wheat Bran and Anti-Inflammatory Effects of the Extracted Polysaccharides. Food Sci. 2018, 39, 192–198. [Google Scholar]
- de Alvarenga, K.A.F.; Sacramento, E.K.; Rosa, D.V.; Souza, B.R.; de Rezende, V.B.; Romano-Silva, M.A. Effects of antipsychotics on intestinal motility in zebrafish larvae. Neurogastroenterol. Motil. 2016, 29, e13006. [Google Scholar] [CrossRef]
- Kim, D.; Koun, S.; Kim, S.Y.; Ha, Y.R.; Choe, J.W.; Jung, S.W.; Hyun, J.J.; Jung, Y.K.; Koo, J.S.; Yim, H.J.; et al. Prokinetic effects of diatrizoate meglumine (Gastrografin) in a zebrafish for opioid-induced constipation model. Anim. Cells Syst. 2021, 25, 264–271. [Google Scholar] [CrossRef]
- Li, W.; Liu, B.; Liu, Z.; Yin, Y.; Xu, G.; Han, M.; Xie, L. Effect of dietary histamine on intestinal morphology, inflammatory status, and gut microbiota in yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish. Immunol. 2021, 117, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Cheng, N.; Nakata, P.; Zhao, L.; Hu, Q. Consumption of polysaccharides from Auricularia auricular modulates the intestinal microbiota in mice. Food Res. Int. 2019, 123, 383–392. [Google Scholar] [CrossRef]
- Liu, W.C.; Zhou, S.H.; Balasubramanian, B.; Zeng, F.Y.; Sun, C.B.; Pang, H.Y. Dietary seaweed (Enteromorpha) polysaccharides improves growth performance involved in regulation of immune responses, intestinal morphology and microbial community in banana shrimp Fenneropenaeus merguiensis. Fish Shellfish. Immunol. 2020, 104, 202–212. [Google Scholar] [CrossRef]
- Tan, X.; Sun, Z.; Zhou, C.; Huang, Z.; Tan, L.; Xun, P.; Huang, Q.; Lin, H.; Ye, C.; Wang, A. Effects of dietary dandelion extract on intestinal morphology, antioxidant status, immune function and physical barrier function of juvenile golden pompano Trachinotus ovatus. Fish Shellfish Immunol. 2018, 73, 197–206. [Google Scholar] [CrossRef]
- Zahran, E.; Risha, E.; AbdelHamid, F.; Mahgoub, H.A.; Ibrahim, T. Effects of dietary Astragalus polysaccharides (APS) on growth performance, immunological parameters, digestive enzymes, and intestinal morphology of Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2014, 38, 149–157. [Google Scholar] [CrossRef]
- Wang, K.; Wu, L.-Y.; Dou, C.-Z.; Guan, X.; Wu, H.-G.; Liu, H.-R. Research Advance in Intestinal Mucosal Barrier and Pathogenesis of Crohn’s Disease. Gastroenterol. Res. Pr. 2016, 2016, 9686238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Zhu, X.M.; Niu, X.T.; Chen, X.M.; Tian, J.X.; Kong, Y.D.; Zhang, D.M.; Zhao, L.; Wang, G.Q. Effects of dietary Allium mongolicum Regel polysaccharide on growth, lipopolysaccharide-induced antioxidant responses and immune responses in Channa argus. Mol. Biol. Rep. 2019, 46, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ran, C.; Wei, K.; Xie, Y.; Xie, M.; Zhou, W.; Yang, Y.; Zhang, Z.; Lv, H.; Ma, X.; et al. The effect of Astragalus polysaccharide on growth, gut and liver health, and anti-viral immunity of zebrafish. Aquaculture 2021, 540, 736677. [Google Scholar] [CrossRef]
- Chi, X.; Liu, Z.; Wang, H.; Wang, Y.; Wei, W.; Xu, B. Royal jelly enhanced the antioxidant activities and modulated the gut microbiota in healthy mice. J. Food Biochem. 2021, 45, e13701. [Google Scholar] [CrossRef] [PubMed]
- Li, X. Extraction, Separation Purification and Biological Activity Analysis of Fermented Wheat Bran Polysaccharide. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2019. [Google Scholar]
- Li, Z.; Tran, N.T.; Ji, P.; Sun, Z.; Wen, X.; Li, S. Effects of prebiotic mixtures on growth performance, intestinal microbiota and immune response in juvenile chu’s croaker, Nibea coibor. Fish Shellfish. Immunol. 2019, 89, 564–573. [Google Scholar] [CrossRef]
- Su, C.; Liu, X.; Lu, Y.; Pan, L.; Zhang, M. Effect of dietary Xiao-Chaihu-Decoction on growth performance, immune response, detoxification and intestinal microbiota of pacific white shrimp (Litopenaeus vannamei). Fish Shellfish. Immunol. 2021, 114, 320–329. [Google Scholar] [CrossRef]
- Su, C.; Fan, D.; Pan, L.; Lu, Y.; Wang, Y.; Zhang, M. Effects of Yu-Ping-Feng polysaccharides (YPS) on the immune response, intestinal microbiota, disease resistance and growth performance of Litopenaeus vannamei. Fish Shellfish. Immunol. 2020, 105, 104–116. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′→3′) | Product Size (bp) | Accession Number |
|---|---|---|---|
| IL-10 | F:ACGCTTCTTCTTTGCGACTG | 295 | NM_001020785.2 |
| R:TTGGGGTTGTGGAGTGCTT | |||
| IL-17 | F:CCCATCCATCCAATCAACAA | 134 | NM_001020787.1 |
| R:ACCTCAACGCCGTCTATCAG | |||
| TJPα | F:CCATTGAGACAGGAGTAAGCATT | 153 | XM_009303250.3 |
| R:ACATCACCAGAGGACTCAACAGA | |||
| Occludin1 | F:GATGTGGAGGACTGGGTCAATA | 183 | NM_212832.2 |
| R:GCCGCTGCTAATAGGGACTG | |||
| Mucin2.1 | F:GCCGCTGCTAATAGGGACTG | 210 | XM_021470771.1 |
| R:CGACAGTTTTCGATTTACGTG | |||
| Mucin5.1 | F:AATAATCTTGCCTGCCCAGAGT | 190 | XM_021470622.1 |
| R:CGACATTGATTTCAGTGATGTTCA | |||
| MMP9 | F:GCCTGCCAAATCAAGGAGTT | 101 | NM_213123.1 |
| R:CGTTCACCATTGCCTGAGAT | |||
| defensin1 | F:GCATCCTTTCCCTGGAGTT | 91 | NM_001081553.1 |
| R:AGCCTAATGGTCCGAAGTAAA |
| Groups | Observed-Species | Goods-Coverage | PD-Whole-Tree | Shannon | Simpson | Chao1 |
|---|---|---|---|---|---|---|
| control | 341.67 ± 98.96 | 0.999 ± 0.0001 | 24.99 ± 4.02 | 1.76 ± 0.09 b | 0.61 ± 0.03 | 274.12 ± 74.44 b |
| 0.05% FWBP | 422.67 ± 78.65 | 0.999 ± 0.0001 | 41.93 ± 5.65 | 3.53 ± 0.51 a | 0.71 ± 0.09 | 565.43 ± 32.47 a |
| 0.1% FWBP | 342.66 ± 88.91 | 0.999 ± 0.0002 | 31.51 ± 6.29 | 1.63 ± 0.31 b | 0.57 ± 0.13 | 270.33 ± 46.10 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Mao, J.; Wang, Y.; Yin, N.; Liu, N.; Zheng, Y.; An, X.; Qi, J.; Wang, R.; Yang, Y. Fermented Wheat Bran Polysaccharides Improved Intestinal Health of Zebrafish in Terms of Intestinal Motility and Barrier Function. Fermentation 2023, 9, 293. https://doi.org/10.3390/fermentation9030293
Chen Q, Mao J, Wang Y, Yin N, Liu N, Zheng Y, An X, Qi J, Wang R, Yang Y. Fermented Wheat Bran Polysaccharides Improved Intestinal Health of Zebrafish in Terms of Intestinal Motility and Barrier Function. Fermentation. 2023; 9(3):293. https://doi.org/10.3390/fermentation9030293
Chicago/Turabian StyleChen, Qiuyan, Jinju Mao, Yuan Wang, Na Yin, Na Liu, Yue Zheng, Xiaoping An, Jingwei Qi, Ruifang Wang, and Yanping Yang. 2023. "Fermented Wheat Bran Polysaccharides Improved Intestinal Health of Zebrafish in Terms of Intestinal Motility and Barrier Function" Fermentation 9, no. 3: 293. https://doi.org/10.3390/fermentation9030293
APA StyleChen, Q., Mao, J., Wang, Y., Yin, N., Liu, N., Zheng, Y., An, X., Qi, J., Wang, R., & Yang, Y. (2023). Fermented Wheat Bran Polysaccharides Improved Intestinal Health of Zebrafish in Terms of Intestinal Motility and Barrier Function. Fermentation, 9(3), 293. https://doi.org/10.3390/fermentation9030293