

Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential

 and
and 
Abstract
:1. Introduction
2. Methodology
2.1. Data Collection
2.2. Data Selection
3. Metabolites of Aspergillus nidulans and their Bioactivities
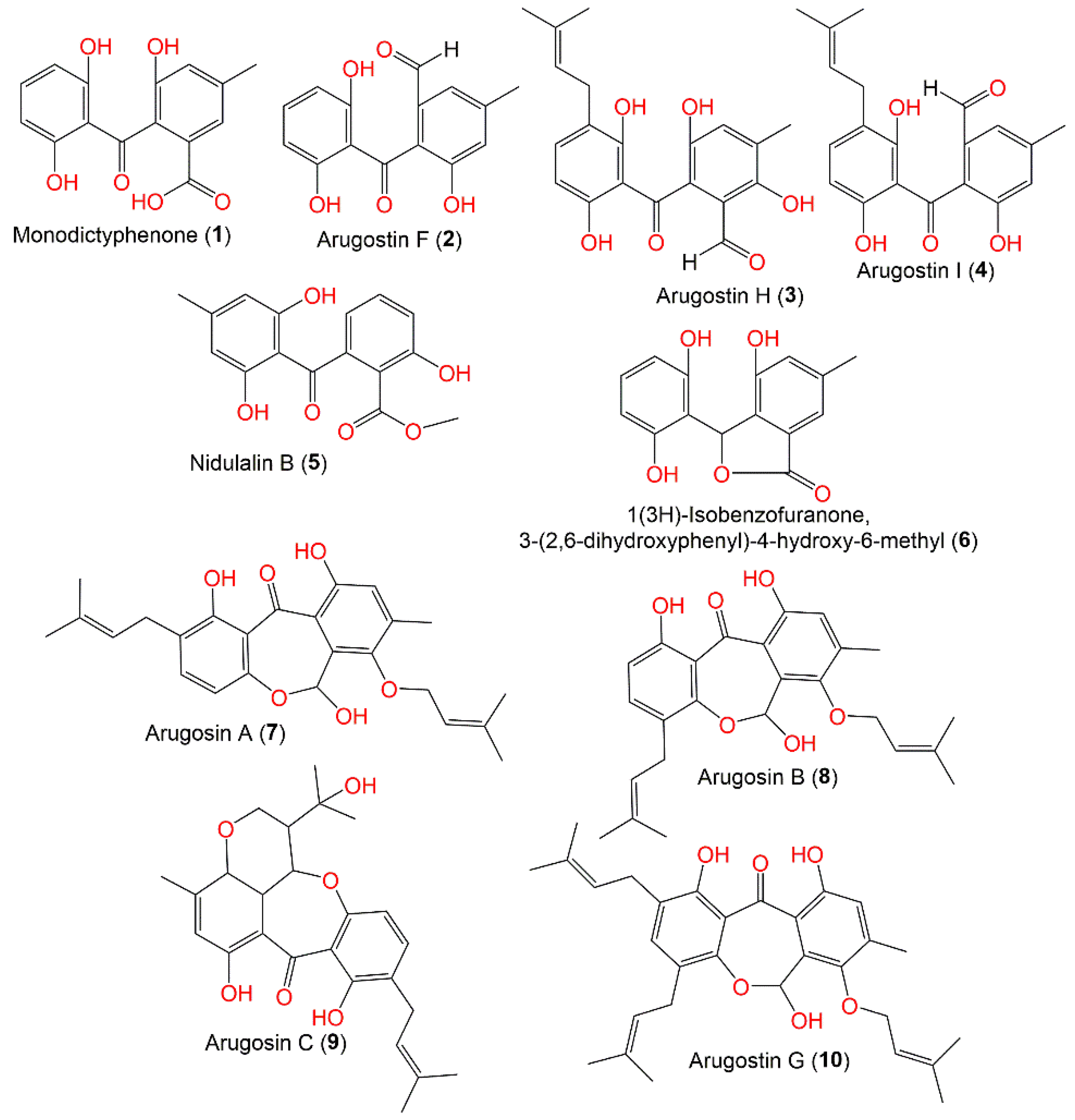
3.1. Benzophenones
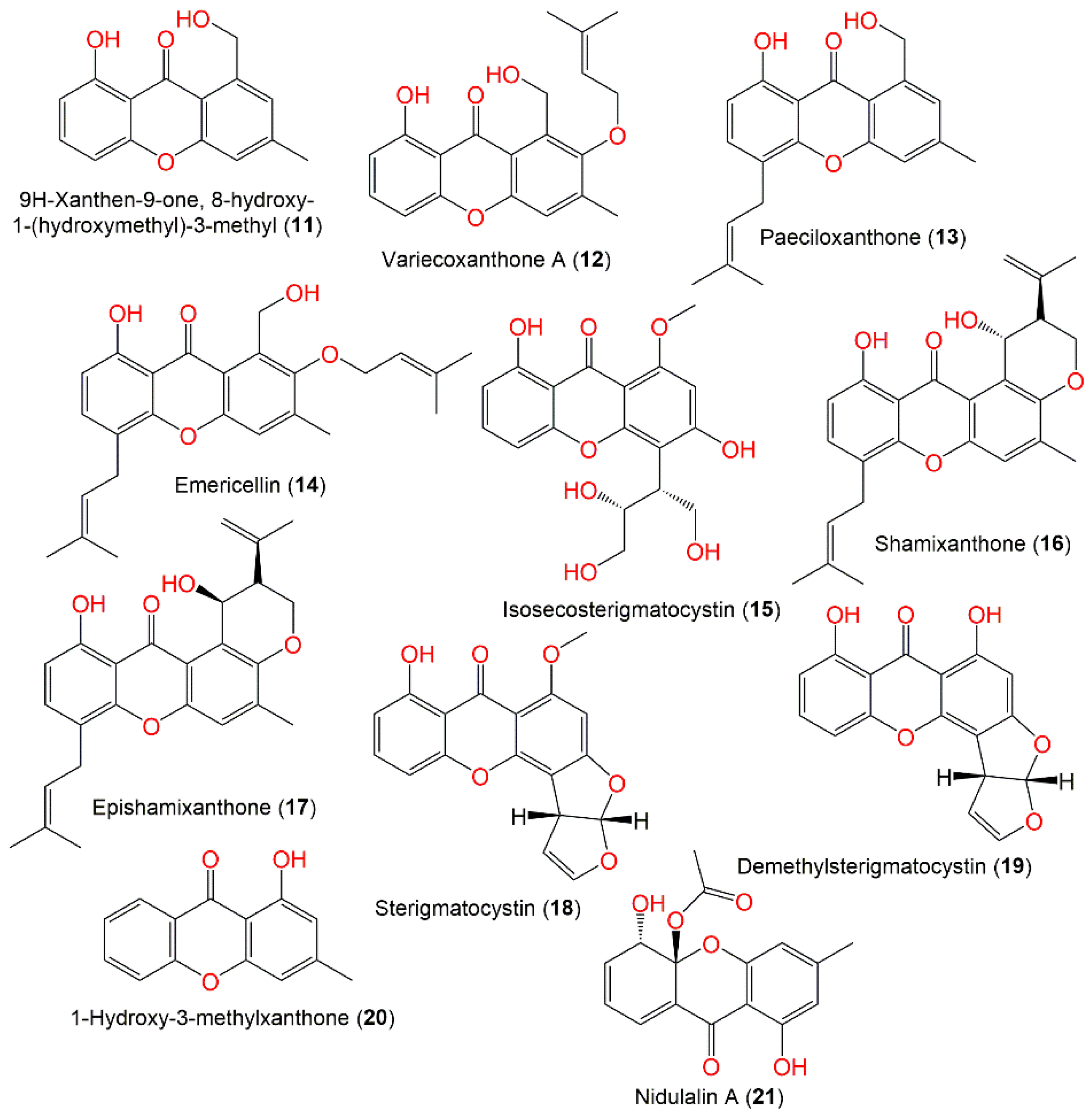
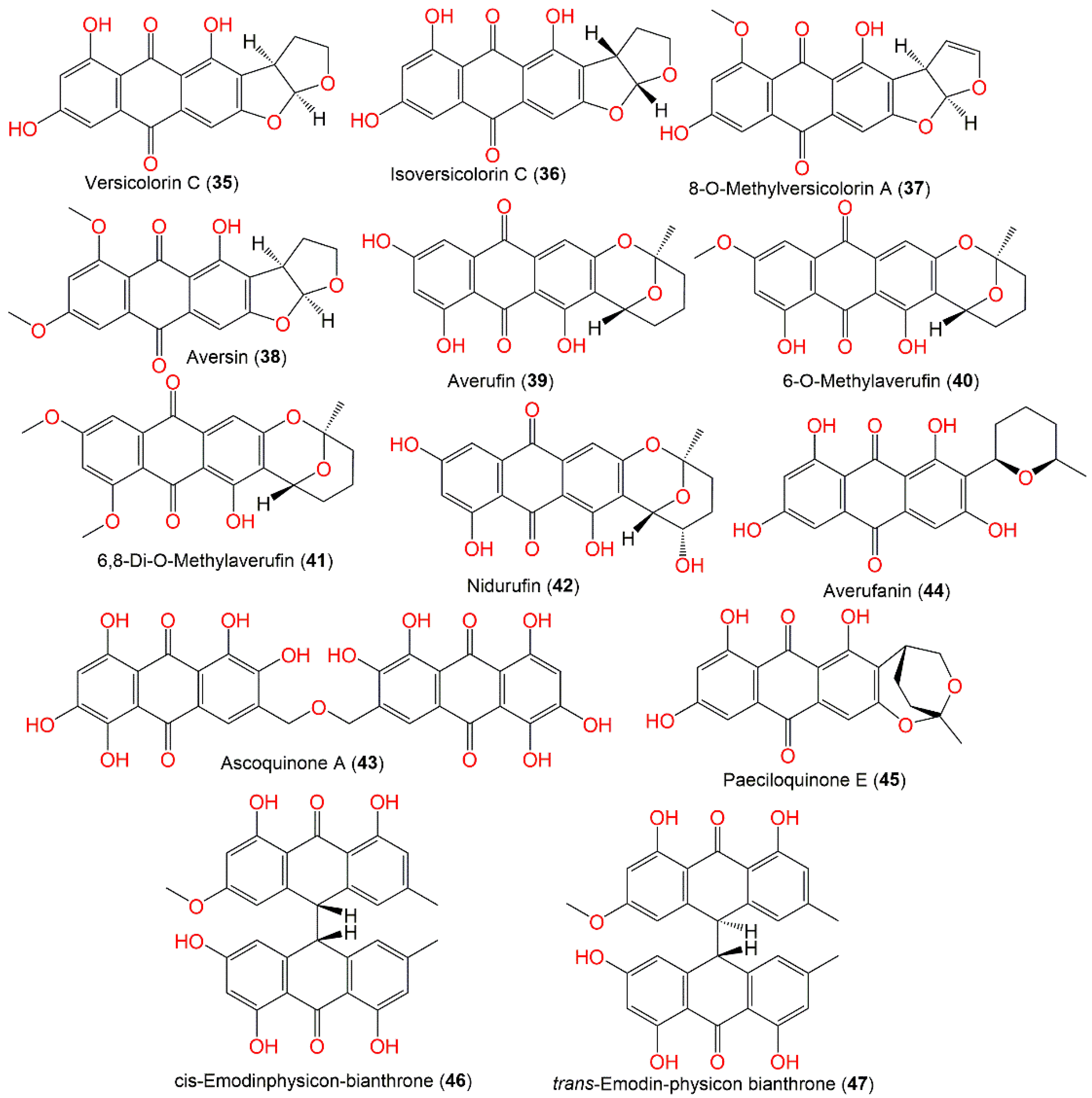
3.2. Xanthones and Quinones
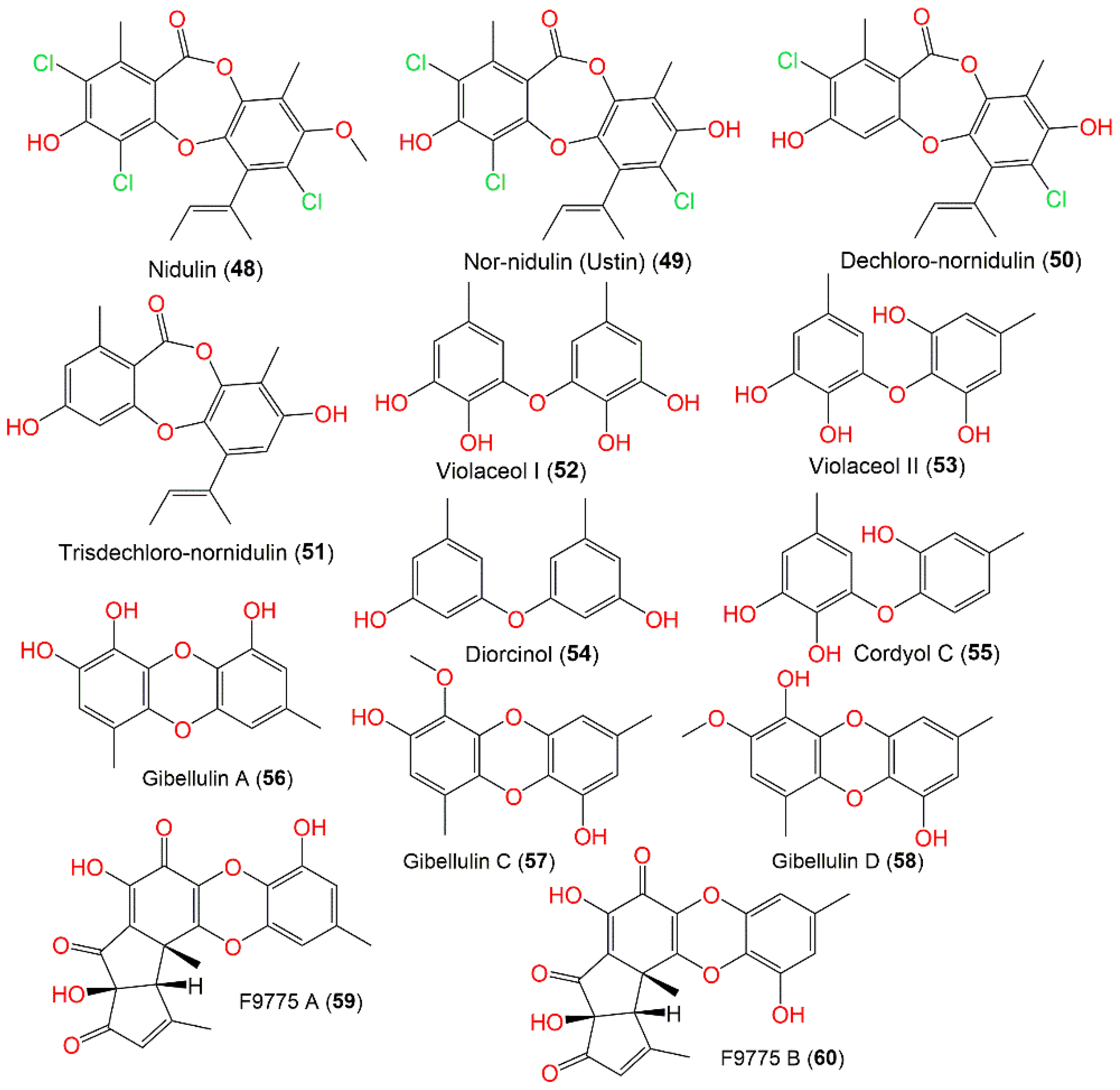
3.3. Depsidones and Biphenyl Ethers
3.4. Alkaloids
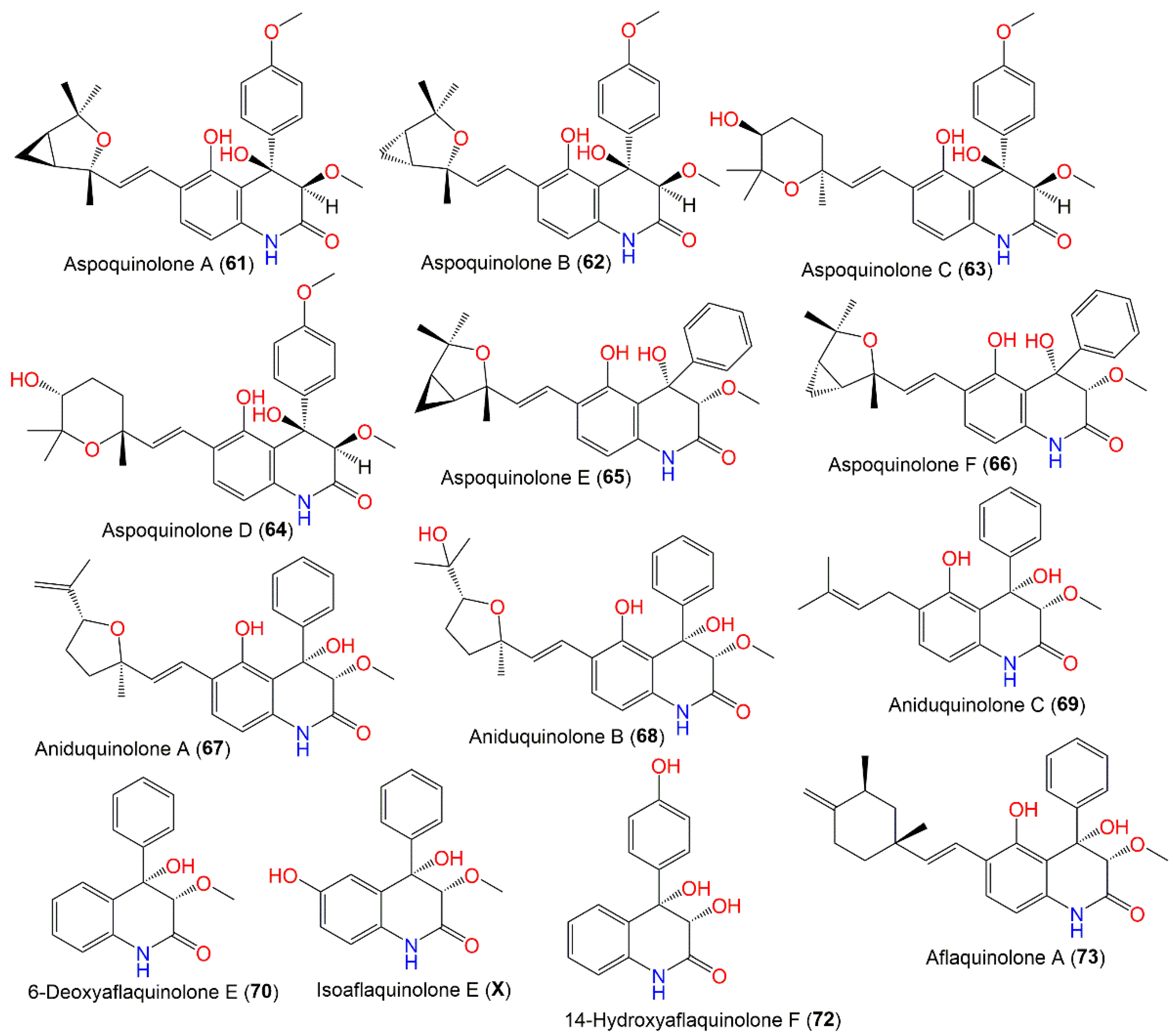
3.4.1. Quinolone Alkaloids
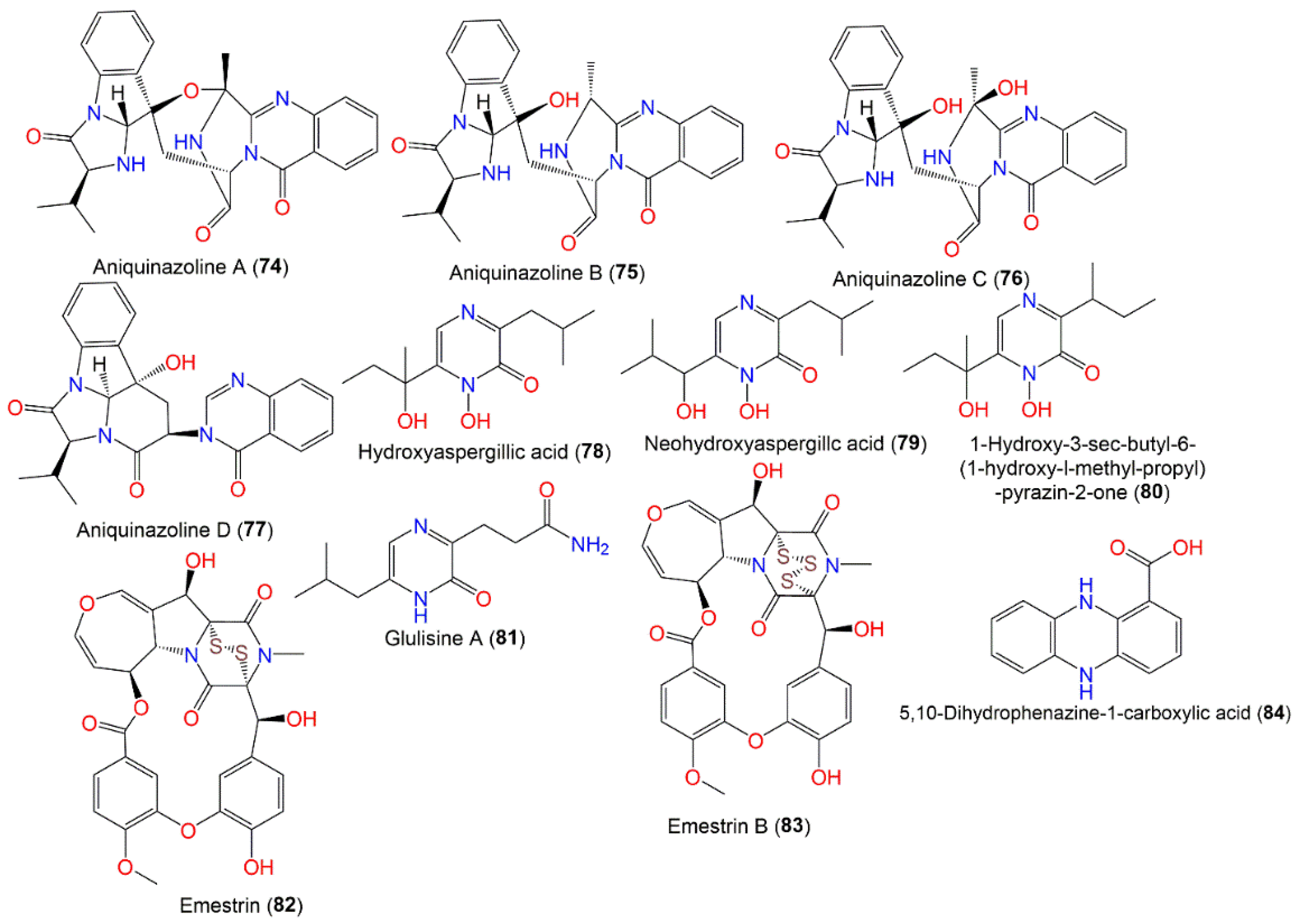
3.4.2. Quinazolinone, Pyrazine, and Dioxopiperazine Alkaloids
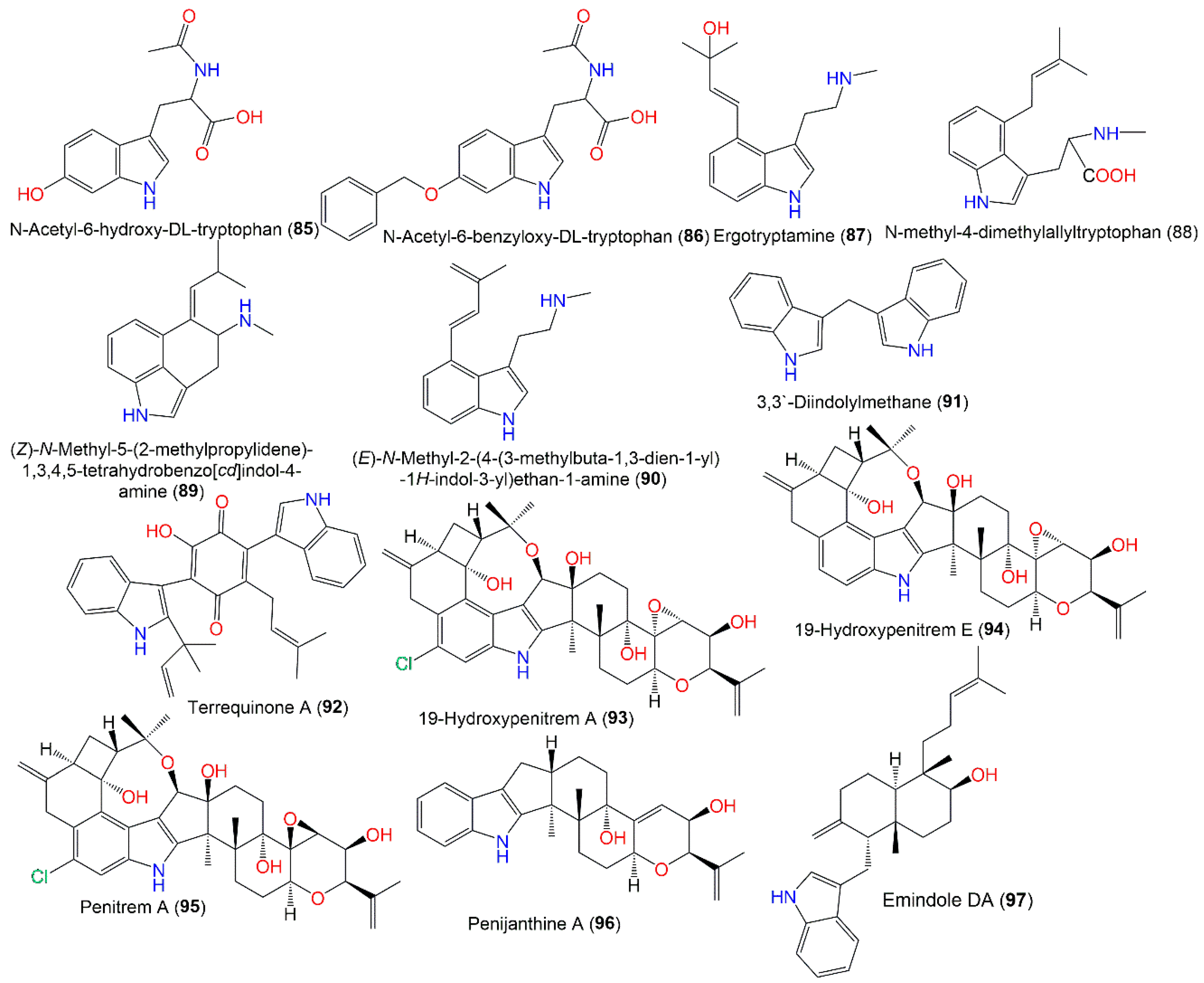
3.4.3. Indole Derivatives
3.4.4. Isoindole Derivatives
3.4.5. Other Nitrogenous Compounds
3.5. Peptides
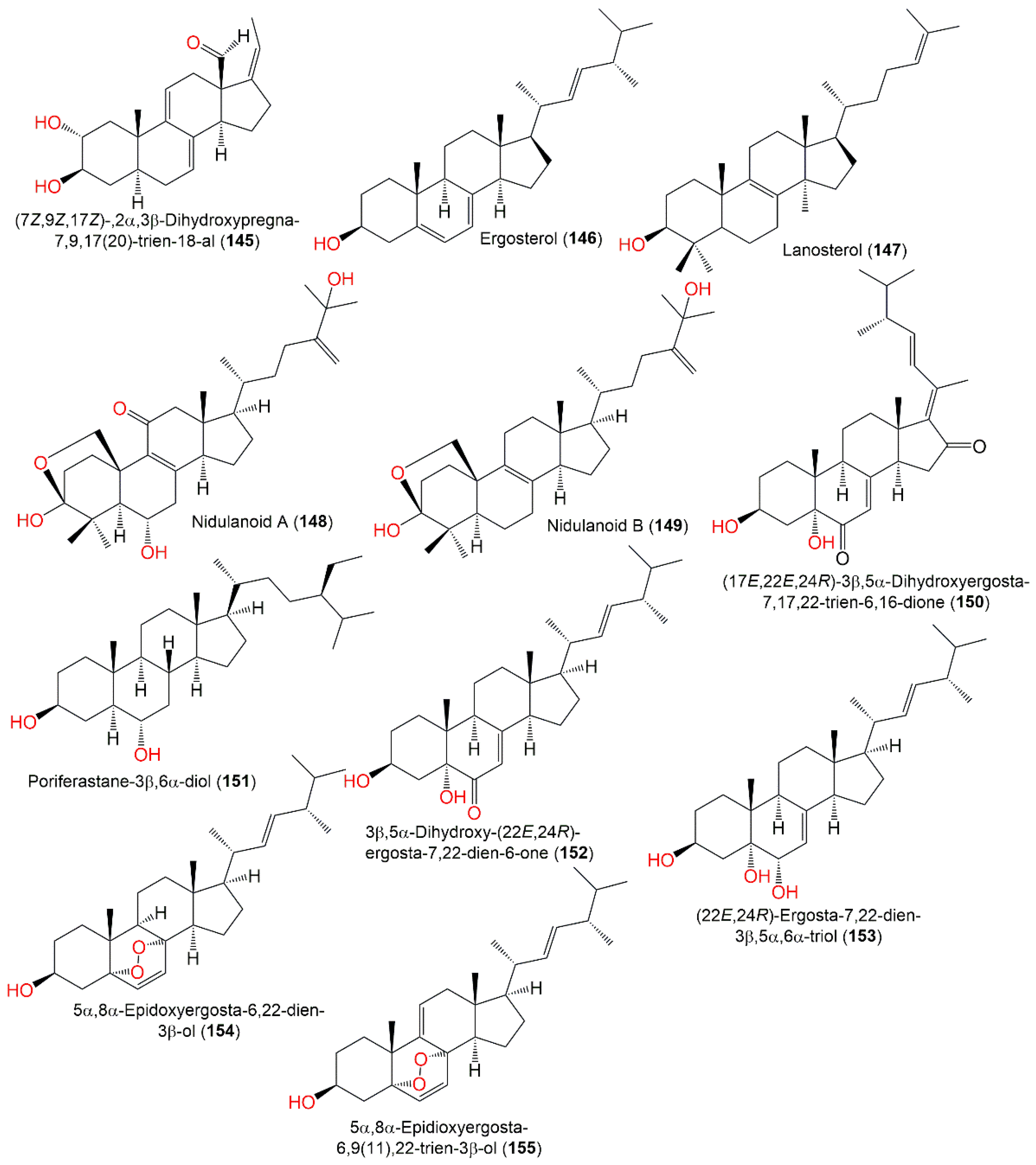
3.6. Terpenoids and Sterols
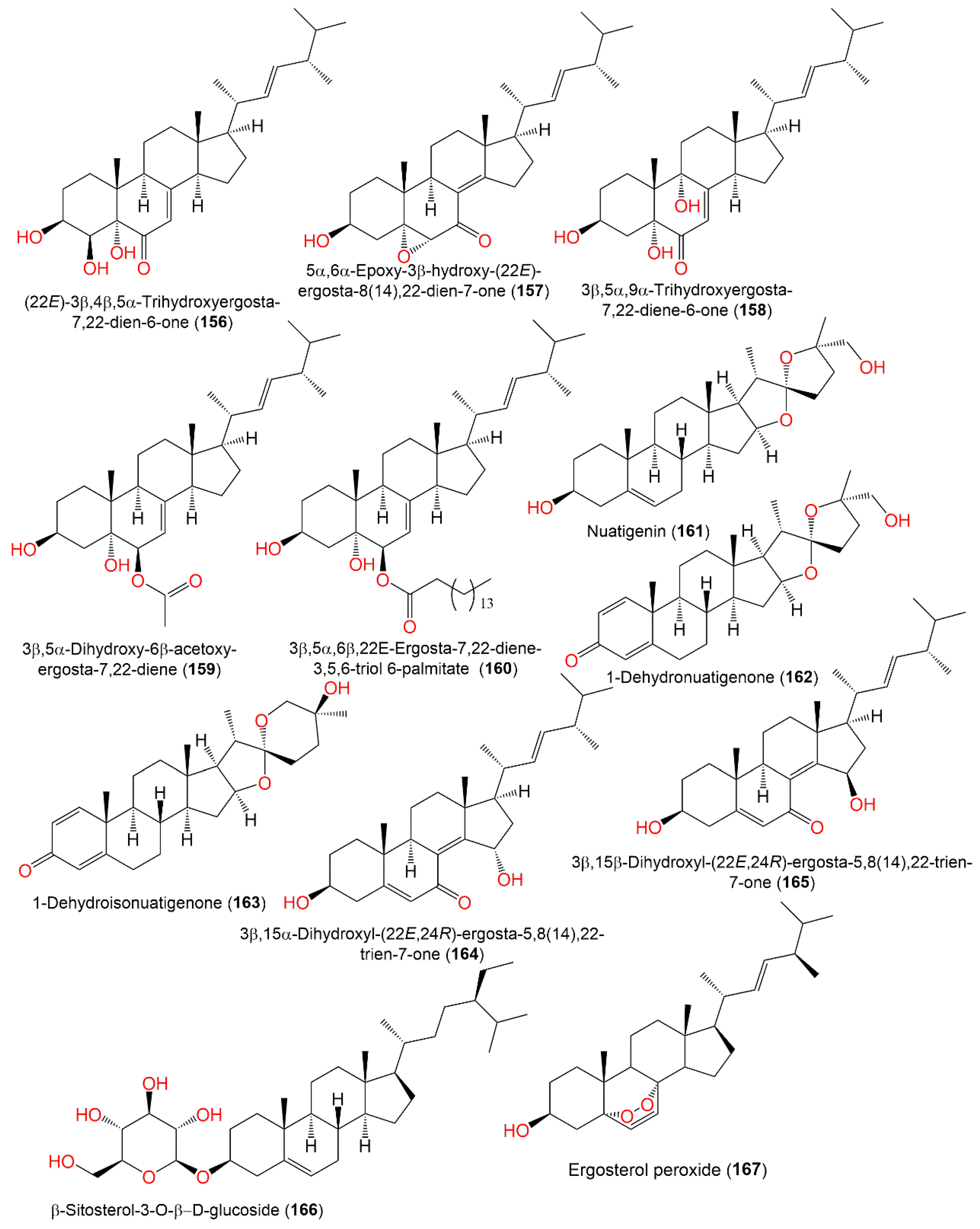
3.7. Lactones and Furanones
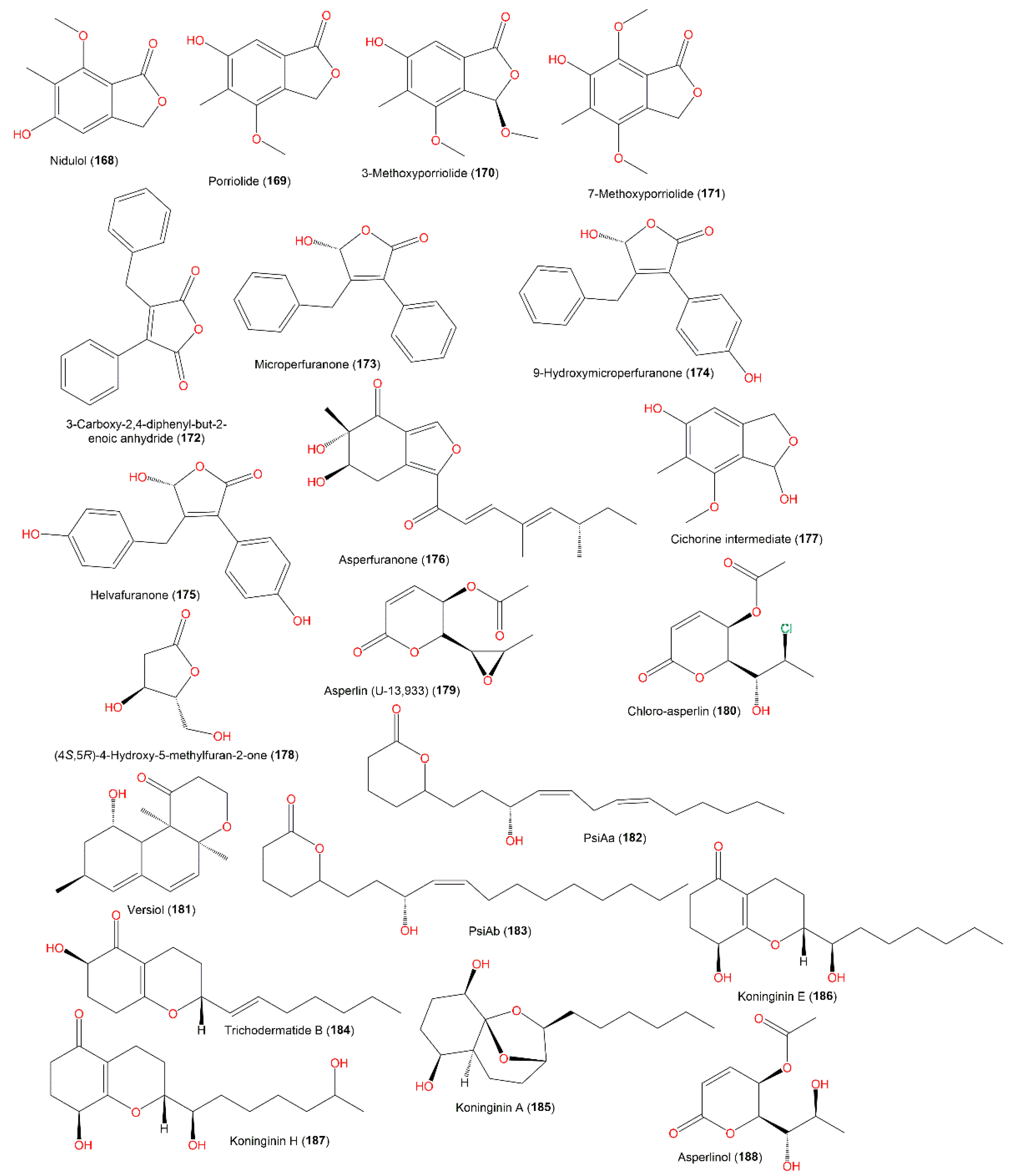
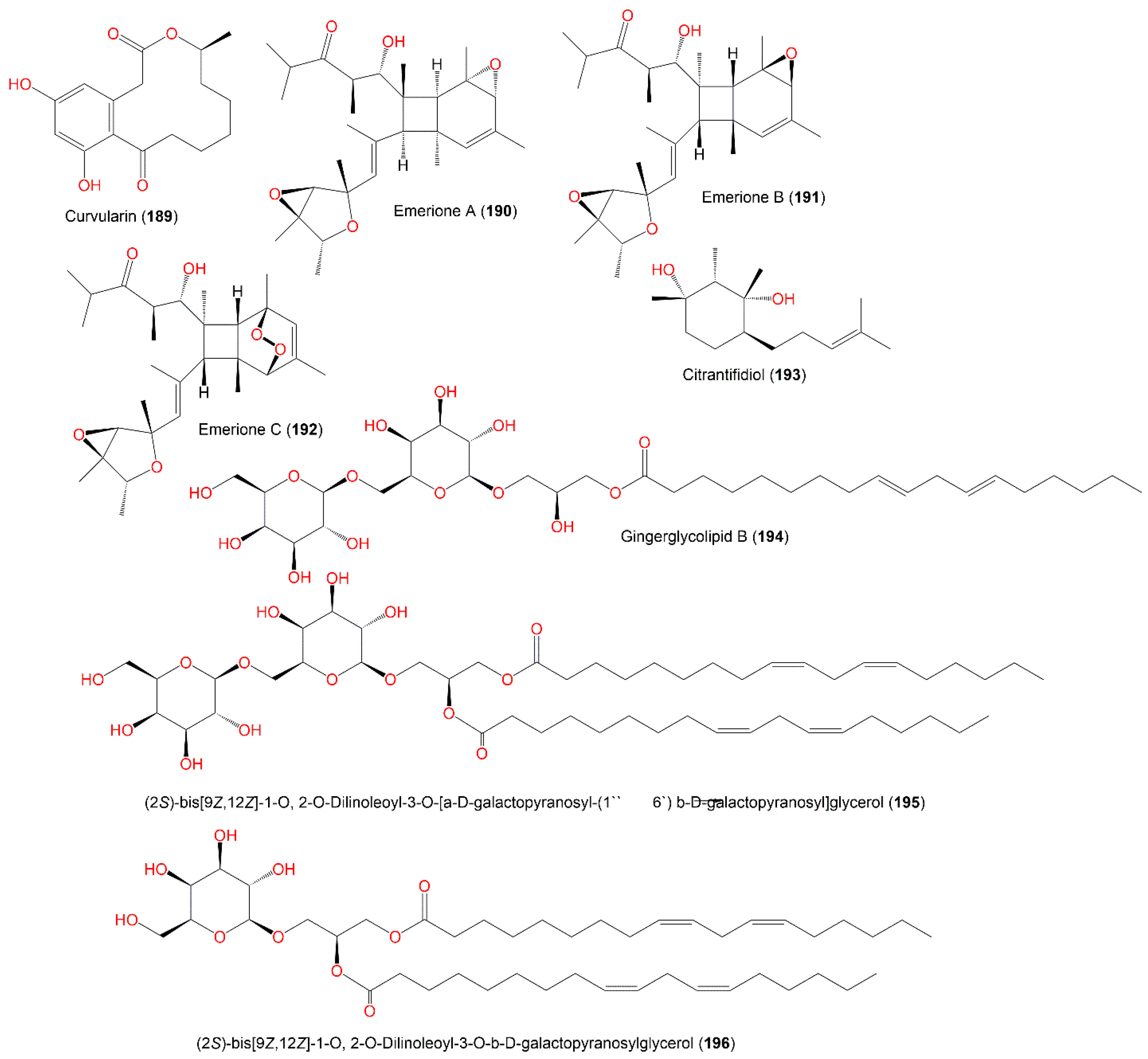
3.8. Polyketides and Glycerides
3.9. Other Metabolites
4. Bioactivities of A. nidulans Extracts
5. Importance of A. nidulans and Its Enzymes
5.1. Aryl Alcohol Oxidase
5.2. Laccases
5.3. Pectate Lyases and Nitroreductases
5.4. Xylanases
5.5. Meta-Cresol Production
5.6. Nanoparticles Synthesis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BEL-7402 | Human hepatocellular carcinoma cell line |
| C28 | Human colon Cell Line |
| CCRF-CEM | Human leukemia cell line |
| CCK-8 | Cell counting kit-8 |
| CD | Circular dichroism |
| ECD | Electronic circular dichroism |
| EtOH | Ethanol |
| EtOAc | Ethyl acetate |
| H-125 | Human lung cancer cell line |
| HCT-116 | Human colon cancer cell line |
| HepG2 | Human hepatocellular liver carcinoma cell line |
| HeLa | Human cervical epitheloid carcinoma cell line |
| HL-60 | Human promyelocytic leukemia cell line |
| HPLC | High-performance liquid chromatography |
| IC50 | Half-maximal inhibitory concentration |
| K562 | Human myelocytic leukemia cell line |
| KB | Human oral epidermoid carcinoma cell line |
| L1210 | Mouse lymphocytic leukemia cell line |
| LD50 | Half maximal lethal concentration |
| LPS | Lipopolysaccharide |
| MCF-7 | Human breast adenocarcinoma cell line |
| MDA-MB-231 | Human breast cancer cell line |
| MIC | Minimum inhibitory concentrations |
| MS | Mass spectrometry |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NCI-H460 | Human lung cancer cell line |
| NMR | Nuclear magnetic resonance |
| NO | Nitric oxide |
| PC12 | Rat brain cancer cell line |
| RP-18 | Reversed phase-18 |
| SRB | Sulforhodamine B |
| SiO2 CC | Silica gel column chromatography |
| T47D | Human breast cancer cell line |
| TLC | Thin layer chromatography |
| WiDr | Human colon adenocarcinoma cell line |
References
- Meyer, V.; Andersen, M.R.; Brakhage, A.A.; Braus, G.H.; Caddick, M.X.; Cairns, T.C.; de Vries, R.P.; Haarmann, T.; Hansen, K.; Hertz-Fowler, C. Current Challenges of Research on Filamentous Fungi in Relation to Human Welfare and a Sustainable Bio-Economy: A White Paper. Fungal Biol. Biotechnol. 2016, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Meyer, V.; Basenko, E.Y.; Benz, J.P.; Braus, G.H.; Caddick, M.X.; Csukai, M.; De Vries, R.P.; Endy, D.; Frisvad, J.C.; Gunde-Cimerman, N. Growing a Circular Economy with Fungal Biotechnology: A White Paper. Fungal Biol. Biotechnol. 2020, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.R.; Choudhry, H.; Asseri, A.H.; Elfaky, M.A.; Mohamed, S.G.; Mohamed, G.A. Stachybotrys Chartarum—A Hidden Treasure: Secondary Metabolites, Bioactivities, and Biotechnological Relevance. J. Fungi 2022, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, D.S.; Ibrahim, S.R.; Ahmed, N.; Khoshhal, S.; Abo-Haded, H.M.; Elkablawy, M.A.; Aljuhani, N.; Mohamed, G.A. Aspernolide F, as a New Cardioprotective Butyrolactone Against Doxorubicin-Induced Cardiotoxicity. Int. Immunopharmacol. 2019, 72, 429–436. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Elkhayat, E.S.; Mohamed, G.A.; Khedr, A.I.; Fouad, M.A.; Kotb, M.H.; Ross, S.A. Aspernolides F and G, New Butyrolactones from the Endophytic Fungus Aspergillus terreus. Phytochem. Lett. 2015, 14, 84–90. [Google Scholar] [CrossRef]
- Chen, S.; Cai, R.; Liu, Z.; Cui, H.; She, Z. Secondary metabolites from mangrove-associated fungi: Source, chemistry and bioactivities. Nat. Prod. Rep. 2022, 39, 560–595. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.; Mohamed, S.G.; Sindi, I.A.; Mohamed, G.A. Biologically Active Secondary Metabolites and Biotechnological Applications of Species of the Family Chaetomiaceae (Sordariales): An Updated Review from 2016 to 2021. Mycol. Progress 2021, 20, 595–639. [Google Scholar] [CrossRef]
- Rustamova, N.; Bozorov, K.; Efferth, T.; Yili, A. Novel secondary metabolites from endophytic fungi: Synthesis and biological properties. Phytochem. Rev. 2020, 19, 425–448. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Bagalagel, A.A.; Diri, R.M.; Noor, A.O.; Bakhsh, H.T.; Muhammad, Y.A.; Mohamed, G.A.; Omar, A.M. Exploring the Activity of Fungal Phenalenone Derivatives as Potential CK2 Inhibitors using Computational Methods. J. Fungi 2022, 8, 443. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.; Mohamed, G.A. Fungal depsides—Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities. Metabolites 2021, 11, 683. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Fadil, S.A.; Fadil, H.A.; Eshmawi, B.A.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Naphthalenones; Promising Metabolites for Drug Discovery: Structures, Biosynthesis, Sources, and Pharmacological Potential. Toxins 2022, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Elkhayat, E.S.; Ross, S.A. Fusarithioamide B, a New Benzamide Derivative from the Endophytic Fungus Fusarium Chlamydosporium with Potent Cytotoxic and Antimicrobial Activities. Bioorg. Med. Chem. 2018, 26, 786–790. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Abdallah, H.M.; Elkhayat, E.S.; Al Musayeib, N.M.; Asfour, H.Z.; Zayed, M.F.; Mohamed, G.A. Fusaripeptide A: New Antifungal and Anti-Malarial Cyclodepsipeptide from the Endophytic Fungus Fusarium sp. J. Asian Nat. Prod. Res. 2018, 20, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.E.; Torres-Mendoza, D.; Caballero E., Z.; Cubilla-Rios, L. Structurally uncommon secondary metabolites derived from endophytic fungi. J. Fungi 2021, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Bright Side of Fusarium Oxysporum: Secondary Metabolites Bioactivities and Industrial Relevance in Biotechnology and Nanotechnology. J. Fungi 2021, 7, 943. [Google Scholar] [CrossRef]
- Hareeri, R.H.; Aldurdunji, M.M.; Abdallah, H.M.; Alqarni, A.A.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Aspergillus Ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential. Molecules 2022, 27, 6759. [Google Scholar] [CrossRef]
- Klich, M.A. Health Effects of Aspergillus in Food and Air. Toxicol. Ind. Health 2009, 25, 657–667. [Google Scholar] [CrossRef]
- Bennett Scott, E.; Bakerand Joan, W. An Overview of the Genus Aspergillus. In The Aspergilli; CRC Press: Boca Raton, FL, USA, 2007; pp. 23–34. [Google Scholar]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the Fungal Workhorses of Industrial Biotechnology: Update on the Mycotoxin and Secondary Metabolite Potential of Aspergillus Niger, Aspergillus Oryzae, and Trichoderma Reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.R.; Mohamed, G.A.; Khedr, A.I. Γ-Butyrolactones from Aspergillus Species: Structures, Biosynthesis, and Biological Activities. Nat. Prod. Commun. 2017, 12, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, Z.; Gao, J. Chemistry and Biology of Secondary Metabolites from Aspergillus Genus. Nat. Prod. J. 2018, 8, 275–304. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.M.; Abdel-Azeem, M.A.; Abdul-Hadi, S.Y.; Darwish, A.G. Aspergillus: Biodiversity, ecological significances, and industrial applications. In Recent Advancement in White Biotechnology through Fungi; Springer: Berlin/Heidelberg, Germany, 2019; pp. 121–179. [Google Scholar]
- Romero, S.M.; Giudicessi, S.L.; Vitale, R.G. Is the Fungus Aspergillus a Threat to Cultural Heritage? J. Cult. Herit. 2021, 51, 107–124. [Google Scholar] [CrossRef]
- Ahluwalia, S.K.; Matsui, E.C. Indoor Environmental Interventions for Furry Pet Allergens, Pest Allergens, and Mold: Looking to the Future. J. Allergy Clin. Immunol. Pract. 2018, 6, 9–19. [Google Scholar] [CrossRef]
- Segal, B.H.; DeCarlo, E.S.; Kwon-Chung, K.J.; Malech, H.L.; Gallin, J.I.; Holland, S.M. Aspergillus Nidulans Infection in Chronic Granulomatous Disease. Medicine 1998, 77, 345–354. [Google Scholar] [CrossRef]
- Henriet, S.S.; Verweij, P.E.; Warris, A. Aspergillus Nidulans and Chronic Granulomatous Disease: A Unique Host–pathogen Interaction. J. Infect. Dis. 2012, 206, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, M.; Hedayati, M.T.; Mirhendi, H.; Nouripour-Sisakht, S.; Hedayati, N.; Saghafi, F.; Mamishi, S. The First Rare and Fatal Case of Invasive Aspergillosis of Spinal Cord due to Aspergillus Nidulans in an Iranian Child with Chronic Granulomatosis Disease: Review of Literature. Curr. Med. Mycol. 2020, 6, 55. [Google Scholar] [CrossRef]
- Kumar, A. Aspergillus nidulans: A Potential Resource of the Production of the Native and Heterologous Enzymes for Industrial Applications. Int. J. Microbiol. 2020, 2020, 8894215. [Google Scholar] [CrossRef]
- Brandl, J.; Andersen, M.R. Aspergilli: Models for Systems Biology in Filamentous Fungi. Curr. Opin. Syst. Biol. 2017, 6, 67–73. [Google Scholar] [CrossRef]
- Yang, S.; Li, X.; Xu, G.; Li, X.; An, C.; Wang, B. Antibacterial Anthraquinone Derivatives Isolated from a Mangrove-Derived Endophytic Fungus Aspergillus nidulans by Ethanol Stress Strategy. J. Antibiot. 2018, 71, 778–784. [Google Scholar] [CrossRef]
- Lü, F.; Li, X.; Chi, L.; Meng, L.; Wang, B. A New Acyclic Peroxide from Aspergillus nidulans SD-531, a Fungus obtained from Deep-Sea Sediment of Cold Spring in the South China Sea. J. Oceanol. Limnol. 2020, 38, 1225–1232. [Google Scholar] [CrossRef]
- Ninomiya, A.; Urayama, S.; Hagiwara, D. Antibacterial Diphenyl Ether Production Induced by Co-Culture of Aspergillus nidulans and Aspergillus fumigatus. Appl. Microbiol. Biotechnol. 2022, 106, 4169–4185. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, C.; He, Y.; Wei, M.; Cheng, L.; Kang, X.; Wang, J.; Hao, X.; Zhu, H.; Zhang, Y. Prenylated Quinolinone Alkaloids and Prenylated Isoindolinone Alkaloids from the Fungus Aspergillus nidulans. Phytochemistry 2020, 169, 112177. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Li, X.; Luo, H.; Li, C.; Wang, M.; Xu, G.; Wang, B. 4-Phenyl-3, 4-Dihydroquinolone Derivatives from Aspergillus nidulans MA-143, an Endophytic Fungus Isolated from the Mangrove Plant Rhizophora stylosa. J. Nat. Prod. 2013, 76, 1896–1901. [Google Scholar] [CrossRef]
- An, C.; Li, X.; Li, C.; Wang, M.; Xu, G.; Wang, B. Aniquinazolines A–D, Four New Quinazolinone Alkaloids from Marine-Derived Endophytic Fungus Aspergillus nidulans. Mar. Drugs 2013, 11, 2682–2694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nursid, M.; Ekowati, C.; Murwantoko; Wahyuono, S. Isolation and Identification of Emestrin from Emericella nidulans and Investigation of its Anticancer Properties. Microbiol. Indones. 2011, 5, 3. [Google Scholar] [CrossRef]
- Nursid, M.; Namirah, I.; Cahyana, A.H.; Fajarningsih, N.D.; Chasanah, E. Emestrin B: Epipolythiodioxypiperazine from Marine Derived Fungus Emericella nidulans. J. Med. Bioeng. 2015, 4, 441–445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, X.; Li, X.; Wang, B. New Indole-Diterpenoids from the Algal-Associated Fungus Aspergillus nidulans. Phytochem. Lett. 2015, 12, 182–185. [Google Scholar] [CrossRef]
- Zhou, H.; Sun, X.; Li, N.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Isoindolone-Containing Meroperpenoids from the Endophytic Fungus Emericella nidulans HDN12–249. Org. Lett. 2016, 18, 4670–4673. [Google Scholar] [CrossRef]
- Li, Q.; Chen, C.; Wei, M.; Dai, C.; Cheng, L.; Tao, J.; Li, X.; Wang, J.; Sun, W.; Zhu, H. Niduterpenoids A and B: Two Sesterterpenoids with a Highly Congested Hexacyclic 5/5/5/5/3/5 Ring System from the Fungus Aspergillus nidulans. Org. Lett. 2019, 21, 2290–2293. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Y.; Fu, A.; Wei, M.; Kang, X.; Chen, C.; Zhu, H.; Zhang, Y. 30-Norlanostane Triterpenoids and Steroid Derivatives from the Endophytic Fungus Aspergillus nidulans. Phytochemistry 2022, 201, 113257. [Google Scholar] [CrossRef]
- Somoza, A.D.; Lee, K.; Chiang, Y.; Oakley, B.R.; Wang, C.C. Reengineering an Azaphilone Biosynthesis Pathway in Aspergillus nidulans to Create Lipoxygenase Inhibitors. Org. Lett. 2012, 14, 972–975. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.F.; Entwistle, R.; Hung, J.; Yaegashi, J.; Jain, S.; Chiang, Y.; Wang, C.C.; Oakley, B.R. Genome-Based Deletion Analysis Reveals the Prenyl Xanthone Biosynthesis Pathway in Aspergillus nidulans. J. Am. Chem. Soc. 2011, 133, 4010–4017. [Google Scholar] [CrossRef] [Green Version]
- Pockrandt, D.; Ludwig, L.; Fan, A.; König, G.M.; Li, S. New Insights into the Biosynthesis of Prenylated Xanthones: Xptb from Aspergillus nidulans Catalyses an O-Prenylation of Xanthones. Chembiochem 2012, 13, 2764–2771. [Google Scholar] [CrossRef] [PubMed]
- Kralj, A.; Kehraus, S.; Krick, A.; Eguereva, E.; Kelter, G.; Maurer, M.; Wortmann, A.; Fiebig, H.; König, G.M. Arugosins G and H: Prenylated Polyketides from the Marine-Derived Fungus Emericella nidulans Var. a Cristata. J. Nat. Prod. 2006, 69, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, N.; Sekita, S.; Satake, M.; Udagawa, S.; Kawai, K. Structures of a New Dihydroxanthone Derivative, Nidulalin A, and a New Benzophenone Derivative, Nidulalin B, from Emericella nidulans. Chem. Pharm. Bull. 1994, 42, 1720–1723. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; El-Kassem, L.T.; Ahmed, E.F.; Emam, M. In-Vitro Bioassays on the Metabolites of the Fungus Emericella nidulans Isolated from the Egyptian Red Sea Algae. Egypt. Pharm. J. 2012, 11, 124. [Google Scholar]
- Ishida, M.; Hamasaki, T.; Hatsuda, Y. The Structure of Two New Metabolites, Emerin and Emericellin, from Aspergillus nidulans. Agric. Biol. Chem. 1975, 39, 2181–2184. [Google Scholar] [CrossRef]
- Yaegashi, J.; Praseuth, M.B.; Tyan, S.; Sanchez, J.F.; Entwistle, R.; Chiang, Y.; Oakley, B.R.; Wang, C.C. Molecular Genetic Characterization of the Biosynthesis Cluster of a Prenylated Isoindolinone Alkaloid Aspernidine A in Aspergillus nidulans. Org. Lett. 2013, 15, 2862–2865. [Google Scholar] [CrossRef] [Green Version]
- Nasr, H.M.; Hawas, U.W.; Mousa, S.A.; Alasmaey, M.; Ahmed, E.F. Isolation and Identification of Emericella nidulans Secondary Metabolites. Chem. Res. J. 2018, 3, 114–119. [Google Scholar]
- Aukamp, P.J.; Holzapfel, C.W. Polyhydroxyanthraquinones from Aspergillus Versicolor, Aspergillus nidulans and Bipolaris Sp. their Significance in Relation to Biogenetic Theories on Aflatoxin B1. S. Afr. J. Chem. 1970, 23, 40–56. [Google Scholar]
- Furukawa, T.; Fukuda, T.; Nagai, K.; Uchida, R.; Tomoda, H. Helvafuranone Produced by the Fungus Aspergillus nidulans BF0142 Isolated from Hot Spring-Derived Soil. Nat. Prod. Commun. 2016, 11, 1001–1003. [Google Scholar] [CrossRef] [Green Version]
- Sana, T.; Siddiqui, B.S.; Shahzad, S.; Farooq, A.D.; Siddiqui, F.; Sattar, S.; Begum, S. Antiproliferative Activity and Characterization of Metabolites of Aspergillus nidulans: An Endophytic Fungus from Nyctanthes Arbor-Tristis Linn. Against Three Human Cancer Cell Lines. Med. Chem. 2019, 15, 352–359. [Google Scholar] [CrossRef]
- Paranjape, S.R.; Chiang, Y.; Sanchez, J.F.; Entwistle, R.; Wang, C.C.; Oakley, B.R.; Gamblin, T.C. Inhibition of Tau Aggregation by Three Aspergillus nidulans Secondary Metabolites: 2, Ω-Dihydroxyemodin, Asperthecin, and Asperbenzaldehyde. Planta Med. 2014, 80, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Salvo, J.J. Isolation and Characterization of Sexual Spore Pigments from Aspergillus nidulans. Appl. Environ. Microbiol. 1994, 60, 979–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, M.F.; Entwistle, R.; Oakley, C.E.; Wang, C.C.; Oakley, B.R. Overexpression of an LaeA-Like Methyltransferase Upregulates Secondary Metabolite Production in Aspergillus nidulans. ACS Chem. Biol. 2019, 14, 1643–1651. [Google Scholar] [CrossRef]
- Lin, H.; Lyu, H.; Zhou, S.; Yu, J.; Keller, N.P.; Chen, L.; Yin, W. Deletion of a Global Regulator LaeB Leads to the Discovery of Novel Polyketides in Aspergillus nidulans. Org. Biomol. Chem. 2018, 16, 4973–4976. [Google Scholar] [CrossRef]
- Xing, C.; Wu, J.; Xia, J.; Fan, S.; Yang, X. Steroids and Anthraquinones from the Deep-Sea-Derived Fungus Aspergillus nidulans MCCC 3A00050. Biochem. Syst. Ecol. 2019, 83, 103–105. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically Active Fungal Depsidones: Chemistry, Biosynthesis, Structural Characterization, and Bioactivities. Fitoterapia 2018, 129, 317–365. [Google Scholar] [CrossRef] [PubMed]
- Dean, F.M.; Robertson, A.; ROBERTS, J.C.; Raper, K.B. Nidulin and ‘Ustin’: Two Chlorine-Containing Metabolic Products of Aspergillus nidulans. Nature 1953, 172, 344. [Google Scholar] [CrossRef] [PubMed]
- Sierankiewicz, J.; Gatenbeck, S. A New Depsidone from Aspergillus nidulans. Acta Chem. Scand. 1972, 26, 455–458. [Google Scholar] [CrossRef] [Green Version]
- Scherlach, K.; Hertweck, C. Discovery of Aspoquinolones A–D, Prenylated Quinoline-2-One Alkaloids from Aspergillus nidulans, Motivated by Genome Mining. Org. Biomol. Chem. 2006, 4, 3517–3520. [Google Scholar] [CrossRef]
- Perry, M.J.; Makins, J.F.; Adlard, M.W.; Holt, G. Aspergillic Acids Produced by Mixed Cultures of Aspergillus Flavus and Aspergillus nidulans. J. Gen. Microbiol. 1984, 130, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.L.; Akhmedov, N.G.; Panaccione, D.G. Identification and Structural Elucidation of Ergotryptamine, a New Ergot Alkaloid Produced by Genetically Modified Aspergillus nidulans and Natural Isolates of Epichloë Species. J. Agric. Food Chem. 2015, 63, 61–67. [Google Scholar] [CrossRef]
- Scherlach, K.; Schuemann, J.; Dahse, H.; Hertweck, C. Aspernidine A and B, Prenylated Isoindolinone Alkaloids from the Model Fungus Aspergillus nidulans. J. Antibiot. 2010, 63, 375–377. [Google Scholar] [CrossRef]
- McCorkindale, N.J.; Hayes, D.; Johnston, G.A.; Clutterbuck, A.J. N-Acetyl-6-Hydroxytryptophan a Natural Substrate of a Monophenol Oxidase from Aspergillus nidulans. Phytochemistry 1983, 22, 1026–1028. [Google Scholar] [CrossRef]
- Kaczka, E.A.; Dulaney, E.L.; Gitterman, C.O.; Woodruff, H.B.; Folkers, K. Isolation and Inhibitory Effects on KB Cell Cultures of 3′-Deoxyadenosine from Aspergillus nidulans (Eidam) Wint. Biochem. Biophys. Res. Commun. 1964, 14, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Tarawneh, A.H.; León, F.; Radwan, M.M.; Rosa, L.H.; Cutler, S.J. Secondary Metabolites from the Fungus Emericella nidulans. Nat. Prod. Commun. 2013, 8, 1285–1288. [Google Scholar] [CrossRef] [Green Version]
- Henke, M.T.; Soukup, A.A.; Goering, A.W.; McClure, R.A.; Thomson, R.J.; Keller, N.P.; Kelleher, N.L. New Aspercryptins, Lipopeptide Natural Products, Revealed by HDAC Inhibition in Aspergillus nidulans. ACS Chem. Biol. 2016, 11, 2117–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benz, F.; Knüsel, F.; Nüesch, J.; Treichler, H.; Voser, W.; Nyfeler, R.; Keller-Schierlein, W. Stoffwechselprodukte Von Mikroorganismen 143. Mitteilung. Echinocandin B, Ein Neuartiges Polypeptid-antibioticum Aus Aspergillus nidulans Var. Echinulatus: Isolierung Und Bausteine. Helv. Chim. Acta 1974, 57, 2459–2477. [Google Scholar] [CrossRef] [PubMed]
- Bromann, K.; Viljanen, K.; Moreira, V.M.; Yli-Kauhaluoma, J.; Ruohonen, L.; Nakari-Setälä, T. Isolation and Purification of Ent-Pimara-8 (14), 15-Diene from Engineered Aspergillus nidulans by Accelerated Solvent Extraction Combined with HPLC. Anal. Methods 2014, 6, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Urra, A.B.; Jiménez, C.; Nieto, M.I.; Rodríguez, J.; Hayashi, H.; Ugalde, U. Signaling the Induction of Sporulation Involves the Interaction of Two Secondary Metabolites in Aspergillus nidulans. ACS Chem. Biol. 2012, 7, 599–606. [Google Scholar] [CrossRef]
- Gealt, M.A. Isolation of Β-Amyrin from the Fungus Aspergillus nidulans. J. Gen. Microbiol. 1983, 129, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, B.E.; Gealt, M.A. Ergosterol and Lanosterol from Aspergillus nidulans. J. Gen. Microbiol. 1982, 128, 1053–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamasaki, T.; Nakajima, H.; Yokota, T.; Kimura, Y. A New Metabolite, 3-Carboxy-2, 4-Diphenyl-but-2-Enoic Anhydride, Produced by Aspergillus nidulans. Agric. Biol. Chem. 1983, 47, 891–892. [Google Scholar] [CrossRef]
- Chiang, Y.; Oakley, C.E.; Ahuja, M.; Entwistle, R.; Schultz, A.; Chang, S.; Sung, C.T.; Wang, C.C.; Oakley, B.R. An Efficient System for Heterologous Expression of Secondary Metabolite Genes in Aspergillus nidulans. J. Am. Chem. Soc. 2013, 135, 7720–7731. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, K.; Katsube, Y.; Noda, A.; Hamasaki, T.; Hatsuda, Y. Absolute Configurations of Asperlin, a Metabolite of Aspergillus nidulans, and its Related Compounds. Bull. Chem. Soc. Jpn. 1978, 51, 3175–3181. [Google Scholar] [CrossRef] [Green Version]
- Argoudelis, A.D.; Zieserl, J.F. The Structure of U-13,933, a New Antibiotic. Tetrahedron Lett. 1966, 7, 1969–1973. [Google Scholar] [CrossRef]
- Mazur, P.; Meyers, H.V.; Nakanishi, K.; Champe, S.P. Structural Elucidation of Sporogenic Fatty Acid Metabolites from Aspergillus nidulans. Tetrahedron Lett. 1990, 31, 3837–3840. [Google Scholar] [CrossRef]
- Li, Q.; Chen, C.; He, Y.; Guan, D.; Cheng, L.; Hao, X.; Wei, M.; Zheng, Y.; Liu, C.; Li, X. Emeriones A–C: Three Highly Methylated Polyketides with Bicyclo [4.2. 0] Octene and 3, 6-Dioxabicyclo [3.1. 0] Hexane Functionalities from Emericella nidulans. Org. Lett. 2019, 21, 5091–5095. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial Enzymes: Industrial Progress in 21st Century. 3 Biotech. 2016, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Han, S.; Kim, H. Industrial Applications of Enzyme Biocatalysis: Current Status and Future Aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Steyer, J.T.; Downes, D.J.; Hunter, C.C.; Migeon, P.A.; Todd, R.B. Duplication and Functional Divergence of Branched-Chain Amino Acid Biosynthesis Genes in Aspergillus nidulans. Mbio 2021, 12, 768. [Google Scholar] [CrossRef]
- Hernández-Ortega, A.; Ferreira, P.; Martínez, A.T. Fungal Aryl-Alcohol Oxidase: A Peroxide-Producing Flavoenzyme Involved in Lignin Degradation. Appl. Microbiol. Biotechnol. 2012, 93, 1395–1410. [Google Scholar] [CrossRef]
- Mathieu, Y.; Piumi, F.; Valli, R.; Aramburu, J.C.; Ferreira, P.; Faulds, C.B.; Record, E. Activities of Secreted Aryl Alcohol Quinone Oxidoreductases from Pycnoporus Cinnabarinus Provide Insights into Fungal Degradation of Plant Biomass. Appl. Environ. Microbiol. 2016, 82, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Li, M.; Abdella, A.; Wilkins, M.R. Development of a Cost-Effective Medium for Submerged Production of Fungal Aryl Alcohol Oxidase using a Genetically Modified Aspergillus nidulans Strain. Bioresour. Technol. 2020, 305, 123038. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Altyar, A.E.; Mohamed, S.G.; Mohamed, G.A. Genus Thielavia: Phytochemicals, Industrial Importance and Biological Relevance. Nat. Prod. Res. 2021, 36, 5108–5123. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R. Untapped Potential of Marine-Associated Cladosporium Species: An Overview on Secondary Metabolites, Biotechnological Relevance, and Biological Activities. Mar. Drugs 2021, 19, 645. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Altyar, A.E.; Mohamed, G.A. Natural Products of the Fungal Genus Humicola: Diversity, Biological Activity, and Industrial Importance. Curr. Microbiol. 2021, 78, 2488–2509. [Google Scholar] [CrossRef]
- Sahay, S.; Chaurse, V.; Chauhan, D. Laccase from Aspergillus nidulans TTF6 Showing Pb Activation for Smaller Substrates and Dyes Remediation in all Climates. In Proceedings of the National Academy of Sciences, India Section B: Biological Sciences; Springer: Berlin/Heidelberg, Germany, 2020; Volume 90, pp. 143–150. [Google Scholar]
- Dutta, N.; Mukhopadhyay, A.; Dasgupta, A.K.; Chakrabarti, K. Nanotechnology Enabled Enhancement of Enzyme Activity and Thermostability: Study on Impaired Pectate Lyase from Attenuated Macrophomina Phaseolina in Presence of Hydroxyapatite Nanoparticle. PLoS ONE 2013, 8, e63567. [Google Scholar] [CrossRef] [Green Version]
- Oumer, O.J. Pectinase: Substrate, Production and their Biotechnological Applications. Int. J. Environ. Agric. Biotechnol. 2017, 2, 238761. [Google Scholar] [CrossRef]
- Suzuki, H.; Morishima, T.; Handa, A.; Tsukagoshi, H.; Kato, M.; Shimizu, M. Biochemical Characterization of a Pectate Lyase AnPL9 from Aspergillus nidulans. Appl. Biochem. Biotechnol. 2022, 194, 5627–5643. [Google Scholar] [CrossRef]
- Roldán, M.D.; Pérez-Reinado, E.; Castillo, F.; Moreno-Vivián, C. Reduction of Polynitroaromatic Compounds: The Bacterial Nitroreductases. FEMS Microbiol. Rev. 2008, 32, 474–500. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lv, H.; Li, H.; Li, J.; Yan, Y.; Liu, F.; Hao, W.; Zhou, Z.; Wang, P.; Zhou, S. Nitroreductase Increases Menadione-Mediated Oxidative Stress in Aspergillus nidulans. Appl. Environ. Microbiol. 2021, 87, 1758. [Google Scholar] [CrossRef]
- Abdella, A.; Segato, F.; Wilkins, M.R. Optimization of Process Parameters and Fermentation Strategy for Xylanase Production in a Stirred Tank Reactor using a Mutant Aspergillus nidulans Strain. Biotechnol. Rep. 2020, 26, e00457. [Google Scholar] [CrossRef]
- Wang, W.; An, C.; Yao, Y.; Meng, X.; Gao, S. De Novo Biosynthesis and Gram-Level Production of M-Cresol in Aspergillus nidulans. Appl. Microbiol. Biotechnol. 2021, 105, 6333–6343. [Google Scholar] [CrossRef]
- Bing, X.; Wang, Z.; Wei, F.; Gao, J.; Xu, D.; Zhang, L.; Wang, Y. Separation of M-Cresol from Coal Tar Model Oil using Propylamine-Based Ionic Liquids: Extraction and Interaction Mechanism Exploration. ACS Omega 2020, 5, 23090–23098. [Google Scholar] [CrossRef]
- Vrsaljko, D.; Haramija, V.; Hadži-Skerlev, A. Determination of Phenol, M-Cresol and O-Cresol in Transformer Oil by HPLC Method. Electr. Power Syst. Res. 2012, 93, 24–31. [Google Scholar] [CrossRef]
- Vijayanandan, A.S.; Balakrishnan, R.M. Biosynthesis of Cobalt Oxide Nanoparticles using Endophytic Fungus Aspergillus nidulans. J. Environ. Manag. 2018, 218, 442–450. [Google Scholar] [CrossRef]
- Vijayanandan, A.S.; Valappil, R.S.K.; Balakrishnan, R.M. Evaluation of Photothermal Properties for Absorption of Solar Energy by Co3O4 Nanofluids Synthesized using Endophytic Fungus Aspergillus nidulans. Sustain. Energy Technol. Assess. 2020, 37, 100598. [Google Scholar] [CrossRef]
- Caesar, L.K.; Kelleher, N.L.; Keller, N.P. In the Fungus Where it Happens: History and Future Propelling Aspergillus nidulans as the Archetype of Natural Products Research. Fungal Genet. Biol. 2020, 144, 103477. [Google Scholar] [CrossRef]




























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Biological Activity | Assay, Organism or Cell Line | Biological Results | Ref. | |
|---|---|---|---|---|---|
| Compound | Positive Control | ||||
| Isosecosterigmatocystin (15) | Antimicrobial | Microplate/Edwardsiella ictaluri | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] |
| Serial dilution/Edwardsiella ictarda | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [31] | ||
| Sterigmatocystin (18) | Antimicrobial | Serial dilution/Vibrio parahaemolyticus | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Versicolorin C (35) | Antimicrobial | Microplate/Escherichia coli | 1.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [30] |
| Microplate/Micrococcus luteus | 32.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio anguillarum | 4.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio alginolyticus | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] | ||
| Microplate/Edwardsiella ictaluri | 8.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio parahaemolyticus | 1.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [30] | ||
| Isoversicolorin C (36) | Antimicrobial | Microplate/Escherichia coli | 32.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [30] |
| Microplate/Micrococcus luteus | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio vulnificus | 64.0 µg/mL (MIC) | Chloramphenicol 8.0 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio alginolyticus | 1.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] | ||
| Microplate/Edwardsiella ictaluri | 4.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] | ||
| Microplate/Vibrio parahaemolyticus | 32.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [30] | ||
| Violaceol I (52) | Antimicrobial | Microdilution/Bacillus subtilis | 13.0 µg/mL (MIC) | Chloramphenicol 6.3 µg/mL (MIC) | [32] |
| Violaceol II (53) | Antimicrobial | Microdilution/Bacillus subtilis | 50 µg/mL (MIC) | Chloramphenicol 6.3 µg/mL (MIC) | [32] |
| Diorcinol (54) | Antimicrobial | Microdilution/Bacillus subtilis | 100.0 µg/mL (MIC) | Chloramphenicol 6.3 µg/mL (MIC) | [32] |
| Cordyol C (55) | Antimicrobial | Microdilution/Bacillus subtilis | 100.0 µg/mL (MIC) | Chloramphenicol 6.3 µg/mL (MIC) | [32] |
| Gibellulin A (56) | Antimicrobial | Serial dilution/Aeromonas hydrophilia | 4.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Colletotrichum gloeosporioides | 4.0 µg/mL (MIC) | Amphotericin B 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Edwardsiella ictarda | 2.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Edwardsiella tarda | 4.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Escherichia coli | 8.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio alginolyticus | 8.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harvey | 4.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio parahaemolyticus | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Aspoquinolone E (65) | Cytotoxicity | MTT/HL-60 | 29.15 µM (IC50) | cis-platin 3.22 µM (IC50) | [33] |
| MTT/A-549 | 3.50 µM (IC50) | cis-platin 13.17 µM (IC50) | [33] | ||
| MTT/MCF-7 | 24.50 µM (IC50) | cis-platin 22.96 µM (IC50) | [33] | ||
| Aniduquinolone B (68) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 7.1 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [34] |
| 6-Deoxyaflaquinolone E (70) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 4.5 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [34] |
| Aflaquinolone A (73) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 5.5 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [34] |
| Aniquinazoline A (74) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 1.27 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [35] |
| Aniquinazoline B (75) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 2.11 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [35] |
| Aniquinazoline C (76) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 4.95 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [35] |
| Aniquinazoline D (77) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 3.42 μM (LD50) | Colchicine 88.4 μΜ (LD50) | [35] |
| Glulisine A (81) | Antimicrobial | Microplate/Edwardsiella ictaluri | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [30] |
| Emestrin (82) | MTT/T47D | 1.8 μg/mL (IC50) | - | [36] | |
| MTT/HepG2 | 4.2 μg/mL (IC50) | - | |||
| MTT/C28 | 2.6 μg/mL (IC50) | - | |||
| MTT/HeLa | 13.8 μg/mL (IC50) | - | |||
| Emestrin B (83) | MTT/T47D | 0.16 μg/mL (IC50) | - | [37] | |
| MTT/HeLa | 1.56 μg/mL (IC50) | - | |||
| MTT/WiDr | and 1.02 μg/mL (IC50) | - | |||
| 3,3′-Diindolylmethane (91) | Antimicrobial | Serial dilution/Aeromonas hydrophilia | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Colletotrichum gloeosporioides | 64.0 µg/mL (MIC) | Amphotericin B 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Edwardsiella tarda | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Terrequinone A (92) | Antimicrobial | Serial dilution/Aeromonas hydrophilia | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Colletotrichum gloeosporioides | 2.0 µg/mL (MIC) | Amphotericin B 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Edwardsiella tarda | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Escherichia coli | 2.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio alginolyticus | 2.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harvey | 64.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio parahaemolyticus | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| 19-Hydroxypenitrem A (93) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 3.2 μM (LD50) | Colchicine 10.7 μΜ (LD50) | [38] |
| Antibacterial | Well diffusion/Escherichia coli | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [38] | |
| Well diffusion/Edwardsiella tarda | 16.0 µg/mL (MIC) | Chloramphenicol 16.0 µg/mL (MIC) | [38] | ||
| Well diffusion/S. aureus | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [38] | ||
| Well diffusion/Vibrio anguillarum | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [38] | ||
| 19-Hydroxypenitrem E (94) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 4.6 μM (LD50) | Colchicine 10.7 μΜ (LD50) | [38] |
| Antibacterial | Well diffusion/Escherichia coli | 64.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [38] | |
| Well diffusion/Edwardsiella tarda | 64.0 µg/mL (MIC) | Chloramphenicol 16.0 µg/mL (MIC) | [38] | ||
| Penitrem A (95) | Cytotoxicity | Brine shrimp lethality/ ArtemiArtemia salina | 1.7 μM (LD50) | Colchicine 10.7 μΜ (LD50) | [38] |
| Antibacterial | Well diffusion/Escherichia coli | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [38] | |
| Well diffusion/Edwardsiella tarda | 16.0 µg/mL (MIC) | Chloramphenicol 16.0 µg/mL (MIC) | [38] | ||
| Well diffusion/S. aureus | 16.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [38] | ||
| Well diffusion/Vibrio anguillarum | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [38] | ||
| Penijanthine A (96) | Cytotoxicity | Brine shrimp lethality/Artemia salina | 8.7 μM (LD50) | Colchicine 10.7 μΜ (LD50) | [38] |
| Aspernidine G (103) | Cytotoxicity | MTT/HL-60 | 28.75 µM (IC50) | cis-platin 3.22 µM (IC50) | [33] |
| MTT/MCF-7 | 28.31 µM (IC50) | cis-platin 22.96 µM (IC50) | [33] | ||
| MTT/SW-480 | 21.67 µM (IC50) | cis platin 18.01 µM (IC50) | [33] | ||
| Aspernidine H (104) | Cytotoxicity | MTT/HL-60 | 15.81 µM (IC50) | cis-platin 3.22 µM (IC50) | [33] |
| MTT/A-549 | 33.03 µM (IC50) | cis-platin 13.17 µM (IC50) | [33] | ||
| MTT/SMMC-7721 | 17.17 µM (IC50) | cis-platin 12.65 µM (IC50) | [33] | ||
| MTT/MCF-7 | 17.80 µM (IC50) | cis-platin 22.96 µM (IC50) | [33] | ||
| MTT/SW-480 | 4.77 µM (IC50) | cis-platin 18.01 µM (IC50) | [33] | ||
| Emeriphenolicin E (108) | Cytotoxicity | SRB/HeLa | 4.77 µM (IC50) | Adriamycin | [39] |
| SRB/A549 | 12.04 µM (IC50) | Adriamycin | [39] | ||
| SRB/HCT-116 | 33.05 µM (IC50) | Adriamycin | [39] | ||
| Niduterpenoid A (136) | ERα inhibitors | MCF-7/MTT | 11.42 µM (IC50) | - | [40] |
| (7Z,9Z,17Z)-,2α,3β-dihydroxypregna-7,9,17 (20)-trien-18-al (145) | Cytotoxicity | MTT/PC12 | 7.34 µM (IC50) | Doxorubicin 5.71 µM (IC50) | [41] |
| Microperfuranone (173) | Antimicrobial | Serial dilution/Colletotrichum gloeosporioides | 32.0 µg/mL (MIC) | Amphotericin B 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Edwardsiella ictarda | 4.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Edwardsiella tarda | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Escherichia coli | 32.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harvey | 16.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio parahaemolyticus | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| 9-Hydroxymicroperfuranone (174) | Antimicrobial | Serial dilution/Colletotrichum gloeosporioides | 64.0 µg/mL (MIC) | Amphotericin B 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Edwardsiella tarda | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Escherichia coli | 64.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harvey | 16.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Curvularin (189) | Antimicrobial | Serial dilution/Edwardsiella ictarda | 32.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [31] |
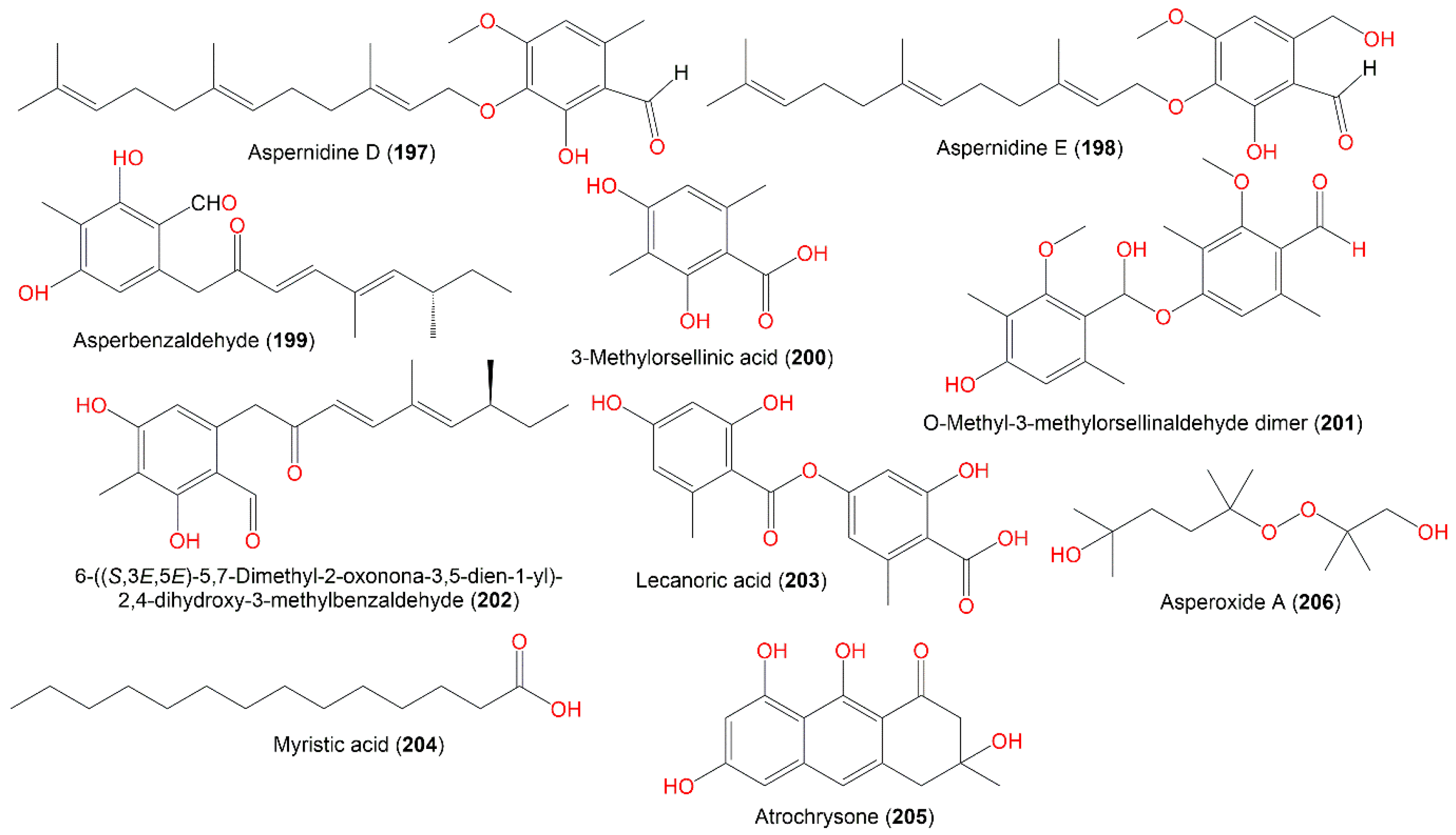
| 6-((S,3E,5E)-5,7-Dimethyl-2-oxonona-3,5-dien-1-yl)-2,4-dihydroxy-3-methylbenzaldehyde (202) | Lipoxygenase-1 inhibition | Spectrophotometric/Soybean lipoxygenase 1 | 97.2 µM (IC50) | - | [42] |
| Lecanoric acid (203) | Antimicrobial | Serial dilution/Aeromonas hydrophilia | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Edwardsiella ictarda | 32.0 µg/mL (MIC) | Chloramphenicol 2.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Escherichia coli | 16.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harvey | 8.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio parahaemolyticus | 4.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Asperoxide A (206) | Antimicrobial | Serial dilution/Aeromonas hydrophilia | 32.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] |
| Serial dilution/Edwardsiella tarda | 16.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio harveyi | 32.0 µg/mL (MIC) | Chloramphenicol 1.0 µg/mL (MIC) | [31] | ||
| Serial dilution/Vibrio parahaemolyticus | 64.0 µg/mL (MIC) | Chloramphenicol 0.5 µg/mL (MIC) | [31] | ||
| Compound Name | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Monodictyphenone (1) | 288 | C15H12O6 | Cultured | [44] |
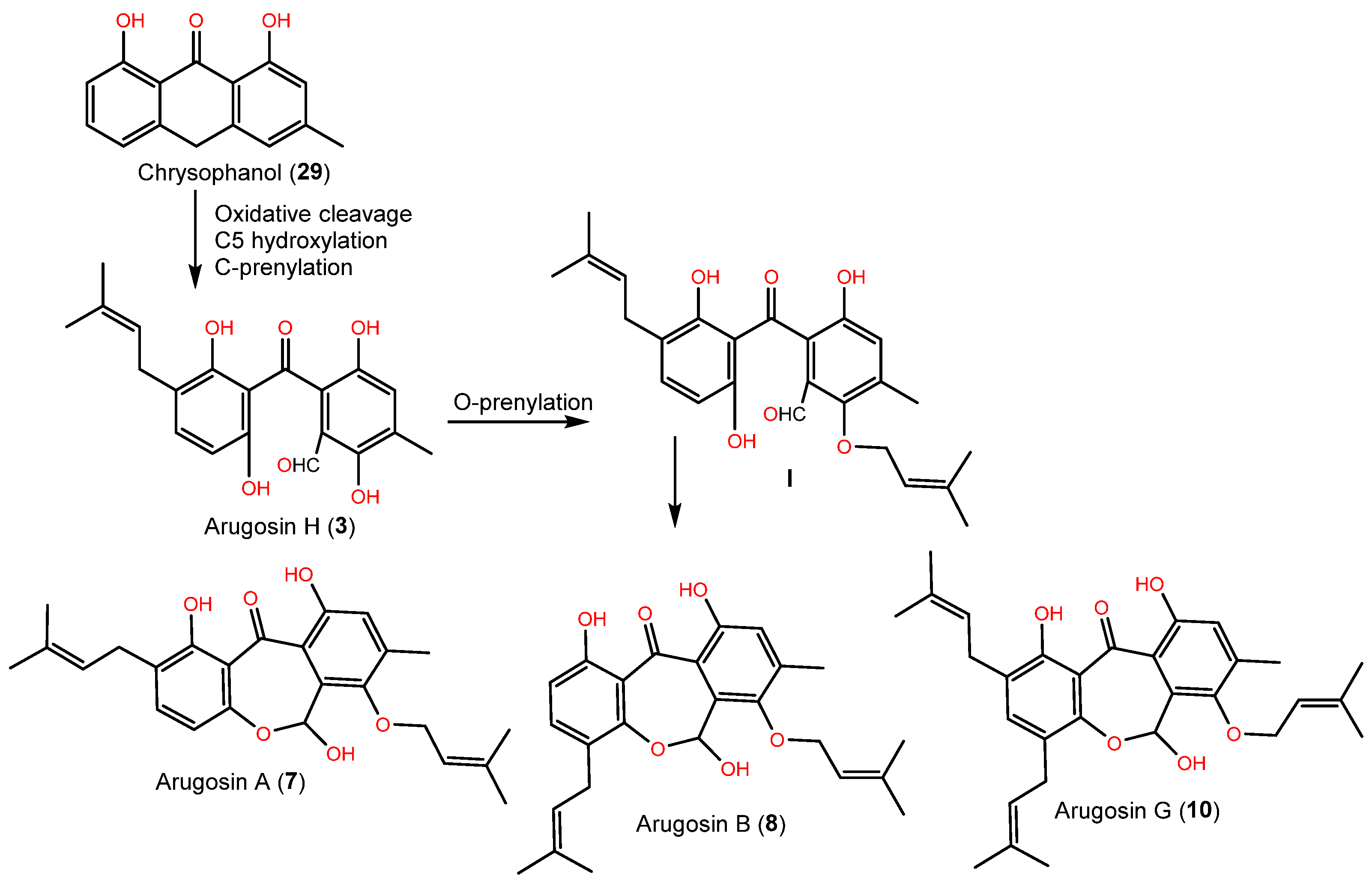
| Arugosin F (2) | 272 | C15H12O5 | Cultured | [45] |
| Arugosin H (3) | 356 | C20H20O6 | Cultured | [45] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| Arugosin I (4) | 340 | C20H20O5 | Cultured | [45] |
| Nidulalin B (5) | 302 | C16H14O6 | Cultured | [46] |
| 1(3H)-Isobenzofuranone, 3-(2,6-dihydroxyphenyl)-4-hydroxy-6-methyl (6) | 272 | C15H12O5 | Cultured | [44] |
| Arugosin A (7) | 424 | C25H28O6 | Cultured | [45] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| Arugosin B (8) | 424 | C25H28O6 | Cultured | [45] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| Arugosin C (9) | 426 | C25H30O6 | Deep-sea sediment, South China Sea | [31] |
| Arugosin G (10) | 492 | C30H36O6 | Green alga, Sardinia, Italy, Mediterranean Sea | [43] |
| Compound Name/Chemical Class | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Xanthones | ||||
| 9H-Xanthen-9-one, 8-hydroxy-1-(hydroxymethyl)-3-methyl (11) | 256 | C15H12O4 | Cultured | [44] |
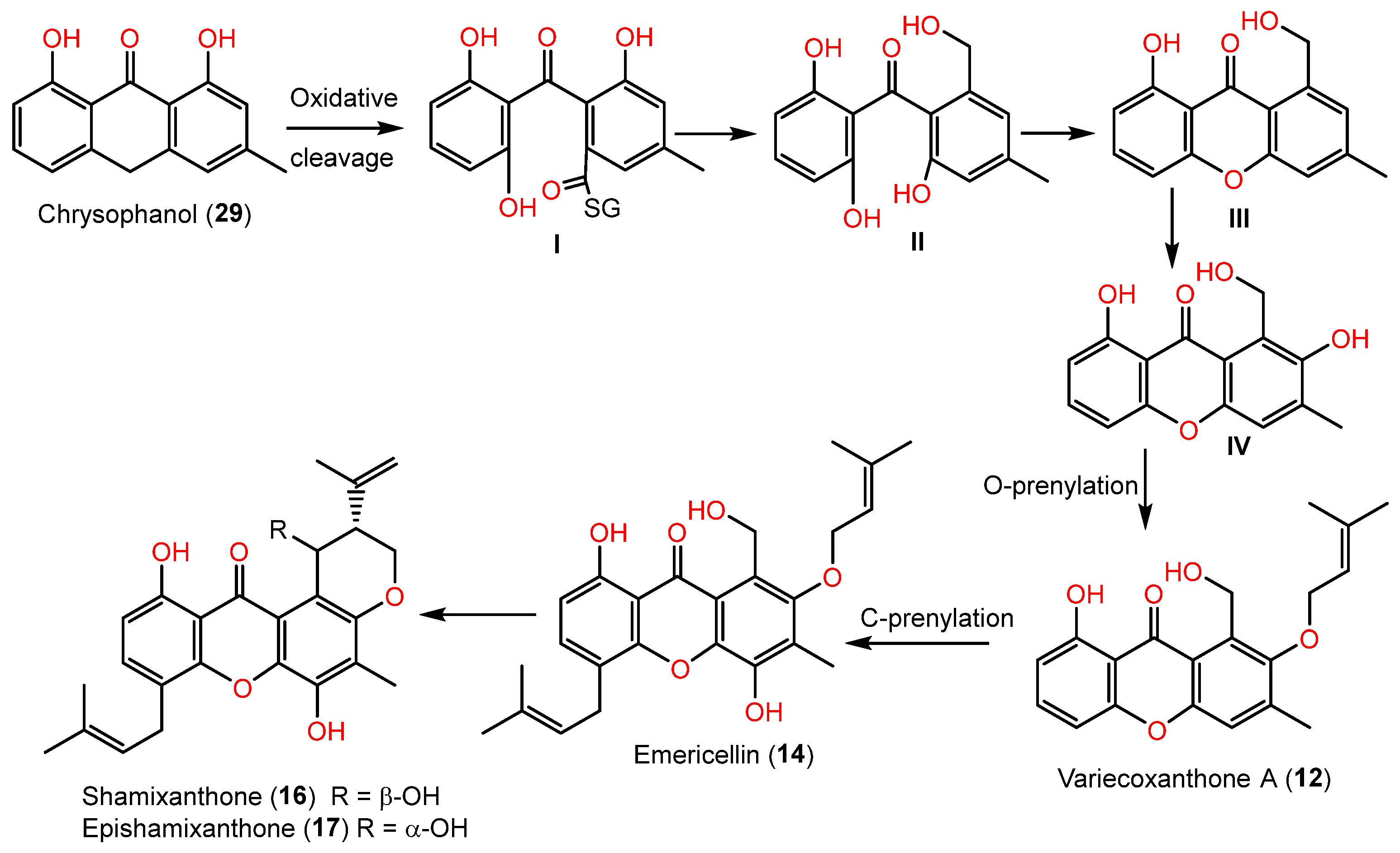
| Variecoxanthone A (12) | 340 | C20H20O5 | Cultured | [44,45] |
| Paeciloxanthone (13) | 324 | C20H20O4 | Cultured | [44] |
| Emericellin (14) | 408 | C25H28O5 | Cultured | [44,45,48,49] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| - | - | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47,50] | |
| Isosecosterigmatocystin (15) | 362 | C18H18O8 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| Shamixanthone (16) | 406 | C25H26O5 | Cultured | [44,45,48,49] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| - | - | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [50] | |
| Epishamixanthone (17) | 406 | C25H26O5 | Cultured | [44,45,49] |
| Sterigmatocystin (18) | 324 | C18H12O6 | Cultured | [48,49,51] |
| - | - | Green alga, Sardinia, Italy, Mediterranean Sea | [43] | |
| - | - | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [52] | |
| - | - | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] | |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| - | - | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47] | |
| - | - | Nyctanthes arbor-tristis (Oleaceae), Sabira | [53] | |
| Demethylsterigmatocystin (19) | 310 | C17H10O6 | Cultured | [54] |
| 1-Hydroxy-3-methylxanthone (20) | 226 | C14H10O3 | Nyctanthes arbor-tristis (Oleaceae), Sabira | [53] |
| Nidulalin A (21) | 302 | C16H14O6 | Cultured | [46] |
| Quinones | ||||
| 1,6,8-Trihydroxyxanthone (22) | 256 | C14H8O5 | Cultured | [55] |
| Emodin (23) | 270 | C15H10O5 | Cultured | [44,55] |
| 2-Hydroxyemodin (24) | 286 | C15H10O6 | Cultured | [56] |
| 2,ω-Dihydroxyemodin (25) | 302 | C15H10O7 | Cultured | [44] |
| ω-Hydroxyemodin (26) | 286 | C15H10O6 | Cultured | [44] |
| Aloe-emodin (27) | 270 | C15H10O5 | Cultured | [44] |
| Endocrocin (28) | 314 | C16H10O7 | Cultured | [44] |
| Chrysophanol (29) | 254 | C15H10O4 | Cultured | [44,45] |
| Asperthecin (30) | 318 | C15H10O8 | Cultured | [49,57] |
| 3′-Hydroxyversiconol (31) | 376 | C18H16O9 | Cultured | [54] |
| Versiconol (32) | 360 | C18H16O8 | Cultured | [56] |
| Averantin (33) | 372 | C20H20O7 | Cultured | [56] |
| Norsolorinic acid (34) | 370 | C20H18O7 | Cultured | [55] |
| - | - | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] | |
| Versicolorin C (35) | 340 | C18H12O7 | Cultured | [48,51] |
| - | - | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] | |
| Isoversicolorin C (36) | 340 | C18H12O7 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] |
| 8-O-Methylversicolorin A (37) | 352 | C19H12O7 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| Aversin (38) | 368 | C20H16O7 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| Averufin (39) | 368 | C20H16O7 | Cultured | [48,51] |
| - | - | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] | |
| 6-O-Methylaverufin (40) | 382 | C21H18O7 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 6,8-Di-O-Methylaverufin (41) | 396 | C22H20O7 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| Nidurufin (42) | 384 | C20H16O8 | Cultured | [51] |
| Ascoquinone A (43) | 618 | C30H18O15 | Cultured | [55] |
| Averufanin (44) | 370 | C20H18O7 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] |
| Paeciloquinone E (45) | 368 | C20H16O7 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] |
| cis-Emodinphysicon-bianthrone (46) | 524 | C31H24O8 | Cultured | [56] |
| trans-Emodin-physicon bianthrone (47) | 524 | C31H24O8 | Cultured | [56] |
| Compound Name/Chemical Class | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Depsidone | ||||
| Nidulin (48) | 443 | C20H17Cl3O5 | Cultured | [60] |
| Nor-nidulin (Ustin) (49) | 427 | C19H15Cl3O5 | Cultured | [60] |
| Dechloro-nornidulin (50) | 394 | C19H16Cl2O5 | Cultured | [61] |
| Trisdechloro-nornidulin (51) | 326 | C19H18O5 | Cultured | [61] |
| Diphenyl ethers | ||||
| Violaceol I (52) | 262 | C14H14O5 | Cultured | [32] |
| Violaceol II (53) | 262 | C14H14O5 | Cultured | [32] |
| Diorcinol (54) | 230 | C14H14O3 | Cultured | [32] |
| - | - | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [52] | |
| - | - | Deep-sea sediment, western Pacific Ocean, China | [58] | |
| Cordyol C (55) | 246 | C14H14O4 | Cultured | [32] |
| Gibellulin A (56) | 360 | C14H12O5 | Deep-sea sediment, South China Sea | [31] |
| Gibellulin C (57) | 274 | C15H14O5 | Cultured | [57] |
| Gibellulin D (58) | 274 | C15H14O5 | Cultured | [57] |
| F9775 A (59) | 396 | C21H16O8 | Cultured | [54] |
| F9775 B (60) | 396 | C21H16O8 | Cultured | [54] |
| Compound Name | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
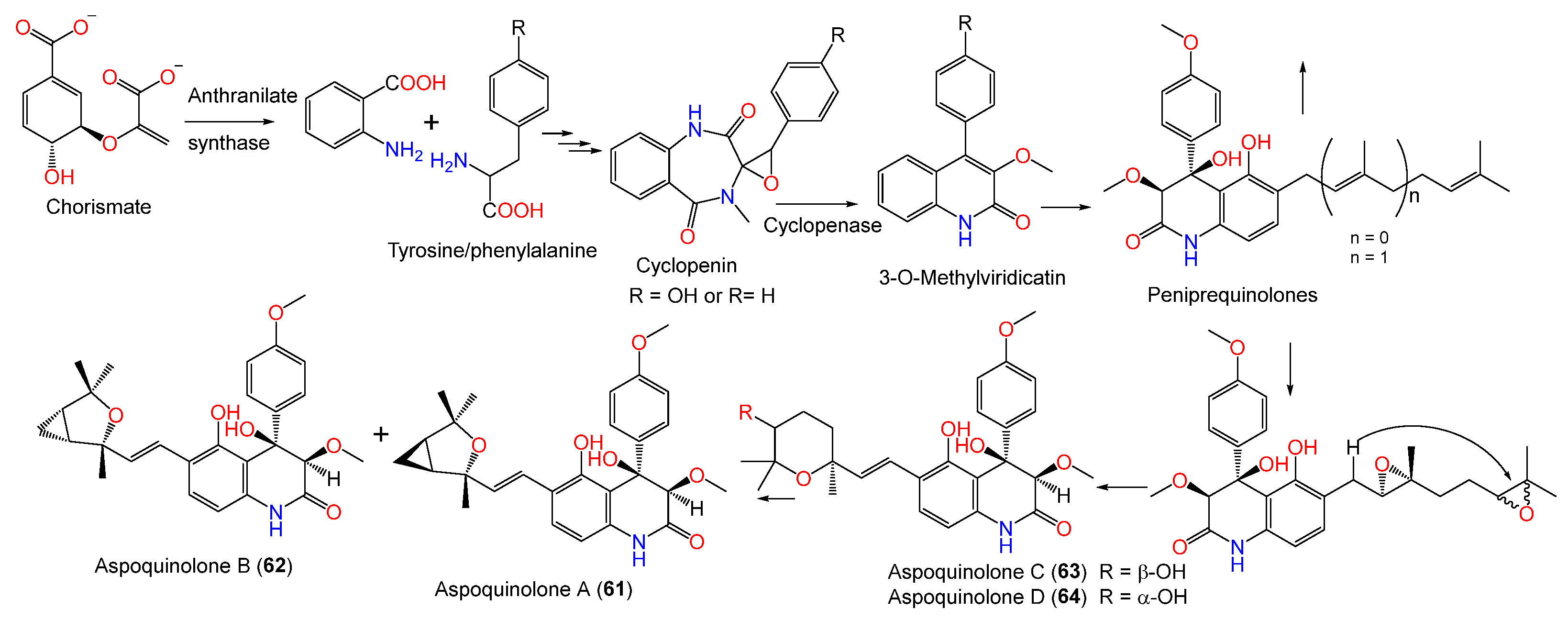
| Aspoquinolone A (61) | 465 | C27H31NO6 | Cultured | [62] |
| Aspoquinolone B (62) | 465 | C27H31NO6 | Cultured | [62] |
| Aspoquinolone C (63) | 483 | C27H33NO7 | Cultured | [62] |
| Aspoquinolone D (64) | 483 | C27H33NO7 | Cultured | [62] |
| Aspoquinolone E (65) | 435 | C26H29NO5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [33] |
| Aspoquinolone F (66) | 435 | C26H29NO5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [33] |
| Aniduquinolone A (67) | 435 | C26H29NO5 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| Aniduquinolone B (68) | 453 | C26H31NO6 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| Aniduquinolone C (69) | 353 | C21H23NO4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| 6-Deoxyaflaquinolone E (70) | 269 | C16H15NO3 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| Isoaflaquinolone E (71) | 285 | C16H15NO4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| 14-Hydroxyaflaquinolone F (72) | 271 | C15H13NO4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| Aflaquinolone A (73) | 433 | C27H31NO4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [34] |
| Compound Name/Chemical Class | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Quinazolinone alkaloids | ||||
| Aniquinazoline A (74) | 471 | C26H25N5O4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [35] |
| Aniquinazoline B (75) | 473 | C26H27N5O4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [35] |
| Aniquinazoline C (76) | 489 | C26H27N5O5 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [35] |
| Aniquinazoline D (77) | 430 | C22H24N4O4 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [35] |
| Pyrazine alkaloids | ||||
| Hydroxyaspergillic acid (78) | 240 | C12H20N2O3 | Cultured | [63] |
| Neohydroxyaspergillc acid (79) | 240 | C12H20N2O3 | Cultured | [63] |
| 1-Hydroxy-3-sec-butyl-6-(1-hydroxy-l-methyl-propyl) -pyrazin-2-one (80) | 240 | C12H20N2O3 | Cultured | [63] |
| Glulisine A (81) | 223 | C11H17N3O2 | Rhizophora stylosa (Mangrove plant fresh leaves, Rhizophoraceae) | [30] |
| Dioxopiperazine alkaloids | ||||
| Emestrin (82) | 598 | C27H22N2O10S2 | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [36,52] |
| Aplidium longithorax (Ascidin, Polyclinidae), Wakatobi Marine National Park, Southeast Sulawesi, Indonesia | [36] | |||
| Emestrin B (83) | 630 | C27H22N2O10S3 | Aplidium longithorax (Marine Ascidin, Polyclinidae), Wakatobi Marine National Park, Southeast Sulawesi, Indonesia | [37] |
| Phenazine alkaloids | ||||
| 5,10-Dihydrophenazine-1-carboxylic acid (84) | 226 | C13H10N2O2 | Nyctanthes arbor-tristis Linn (Oleaceae), Sabira | [53] |
| Compound Name/Chemical Class | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Indole alkaloids | ||||
| N-Acetyl-6-hydroxy-DL-tryptophan (85) | 262 | C13H14N2O4 | Cultured | [66] |
| N-Acetyl-6-benzyloxy-DL-tryptophan (86) | 352 | C20H20N2O4 | Cultured | [66] |
| Ergotryptamine (87) | 258 | C16H22N2O | Mutant strain cultured | [64] |
| N-methyl-4-dimethylallyltryptophan (88) | 286 | C17H22N2O2 | Mutant strain cultured | [64] |
| (Z)-N-Methyl-5-(2-methylpropylidene)-1,3,4,5-tetrahydrobenzo[cd]indol-4-amine (89) | 240 | C16H20N2 | Mutant strain cultured | [64] |
| (E)-N-Methyl-2-(4-(3-methylbuta-1,3-dien-1-yl)-1H-indol-3-yl)ethan-1-amine (90) | 240 | C16H20N2 | Mutant strain cultured | [64] |
| 3,3′-Diindolylmethane (91) | 246 | C17H30N2O3 | Deep-sea sediment, South China Sea | [31] |
| Terrequinone A (92) | 490 | C32H30N2O3 | Deep-sea sediment, South China Sea | [31] |
| 19-Hydroxypenitrem A (93) | 649 | C37H44ClNO7 | Polysiphonia scopulorum var. villum (Red alga, Rhodomelaceae), Yantai, North China | [38] |
| 19-Hydroxypenitrem E (94) | 615 | C37H45NO7 | Polysiphonia scopulorum var. villum (Red alga, Rhodomelaceae), Yantai, North China | [38] |
| Penitrem A (95) | 649 | C37H44ClNO7 | Polysiphonia scopulorum var. villum (Red alga, Rhodomelaceae), Yantai, North China | [38] |
| Penijanthine A (96) | 419 | C27H33NO3 | Polysiphonia scopulorum var. villum (Red alga, Rhodomelaceae), Yantai, North China | [38] |
| Emindole DA (97) | 405 | C28H39NO | Green alga, Sardinia, Italy, Mediterranean Sea | [43] |
| Isoindole alkaloids | ||||
| Cichorine (98) | 193 | C10H11NO3 | Cultured | [44,57] |
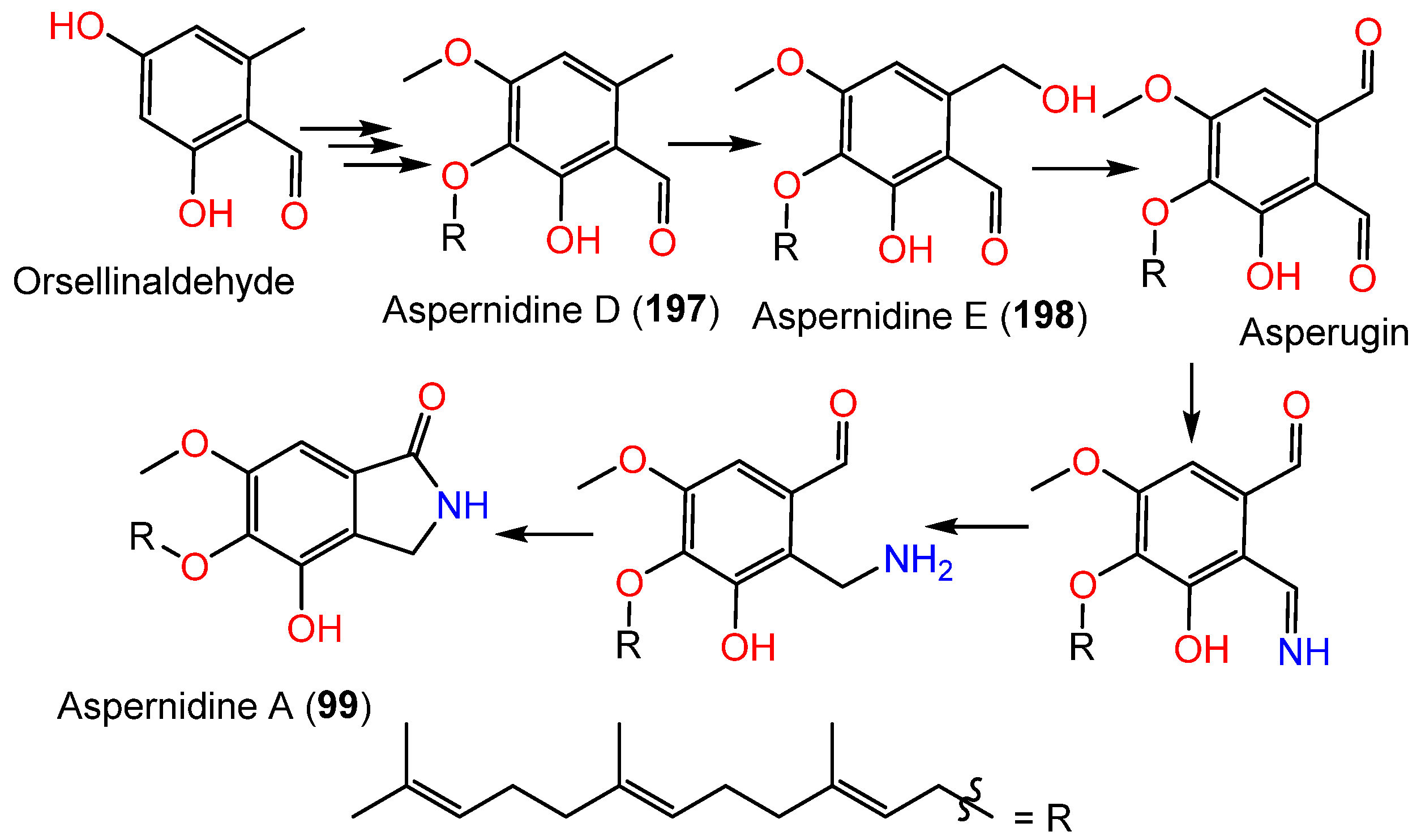
| Aspernidine A (99) | 399 | C24H33NO4 | Cultured | [65] |
| Aspernidine B (100) | 385 | C23H31NO4 | Cultured | [65] |
| Aspernidine C (101) | 413 | C25H35NO4 | Cultured | [49] |
| Aspernidine F (102) | 443 | C26H37NO5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [33] |
| Aspernidine G (103) | 413 | C24H31NO5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [33] |
| Aspernidine H (104) | 457 | C26H35NO6 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [33] |
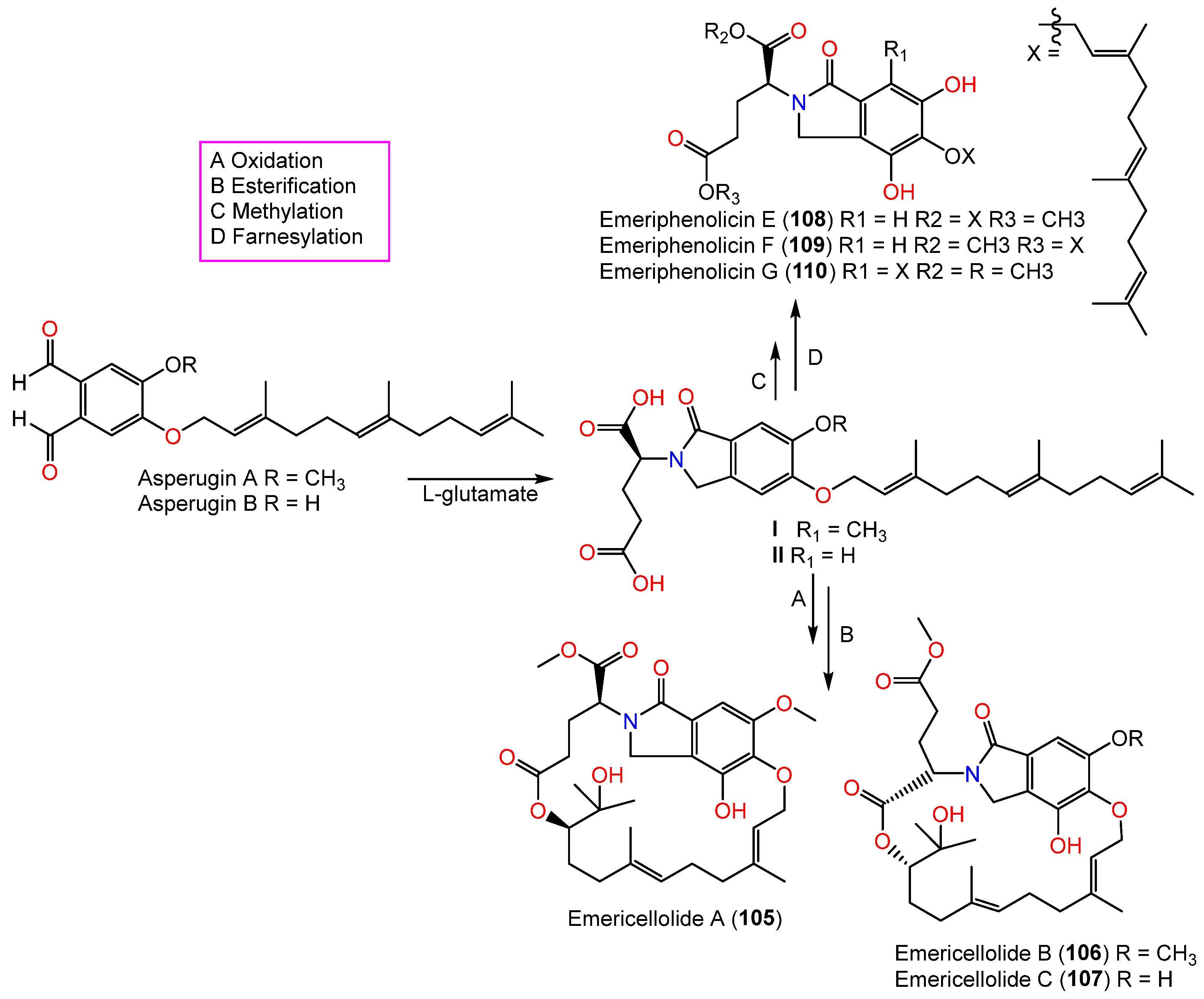
| Emericellolide A (105) | 559 | C30H41NO9 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
| Emericellolide B (106) | 559 | C30H41NO9 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
| Emericellolide C (107) | 545 | C29H39NO9 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
| Emeriphenolicin E (108) | 733 | C44H63NO8 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
| Emeriphenolicin F (109) | 733 | C44H63NO8 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
| Emeriphenolicin G (110) | 747 | C45H65NO8 | Tamarix chinensis (Leaves, Tamaricaceae), Laizhou Bay, China | [39] |
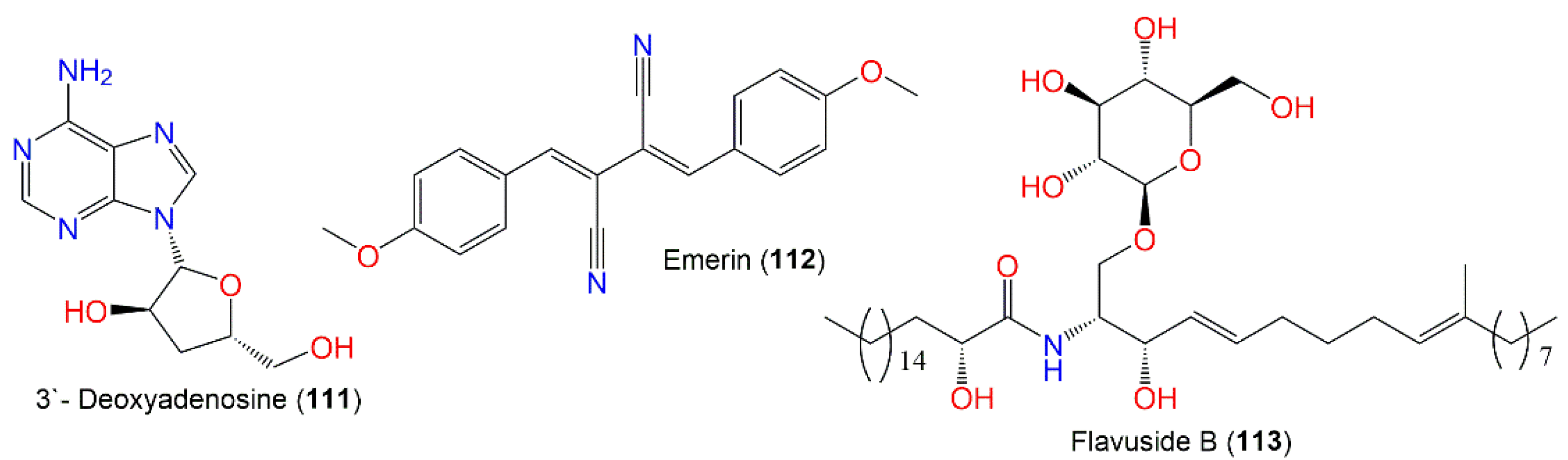
| Compound Name | Mol. Wt. * | Mol. Formula * | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| 3′- Deoxyadenosine (111) | 251 | C10H13N5O3 | Cultured | [67] |
| - | - | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47] | |
| Emerin (112) | 316 | C20H16N2O2 | Cultured | [48] |
| Flavuside B (113) | 755 | C43H81N7O9 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Compound Name | Mol. Wt. | Mol. Formula | Host (Part, Family) | Ref. |
|---|---|---|---|---|
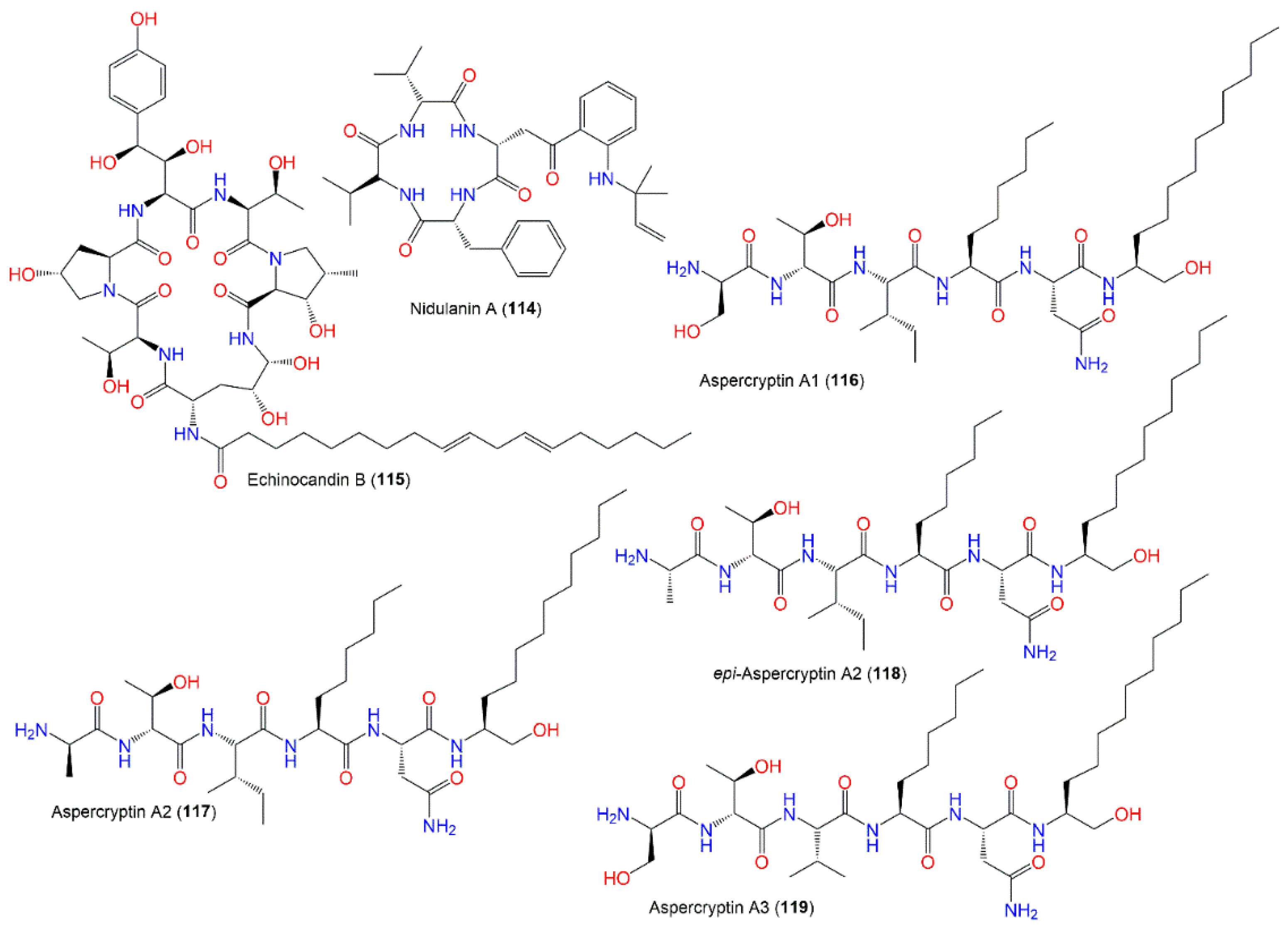
| Nidulanin A (114) | 603 | C34H45N5O5 | Cultured | [56] |
| Echinocandin B (115) | 1059 | C52H81N7O16 | Cultured | [70] |
| Aspercryptin A1 (116) | 757 | C37H71N7O9 | Mutant strain cultured | [69] |
| Aspercryptin A2 (117) | 741 | C37H71N7O8 | Mutant strain cultured | [69] |
| epi-Aspercryptin A2 (118) | 741 | C37H71N7O8 | Mutant strain cultured | [69] |
| Aspercryptin A3 (119) | 743 | C36H69N7O9 | Mutant strain cultured | [69] |
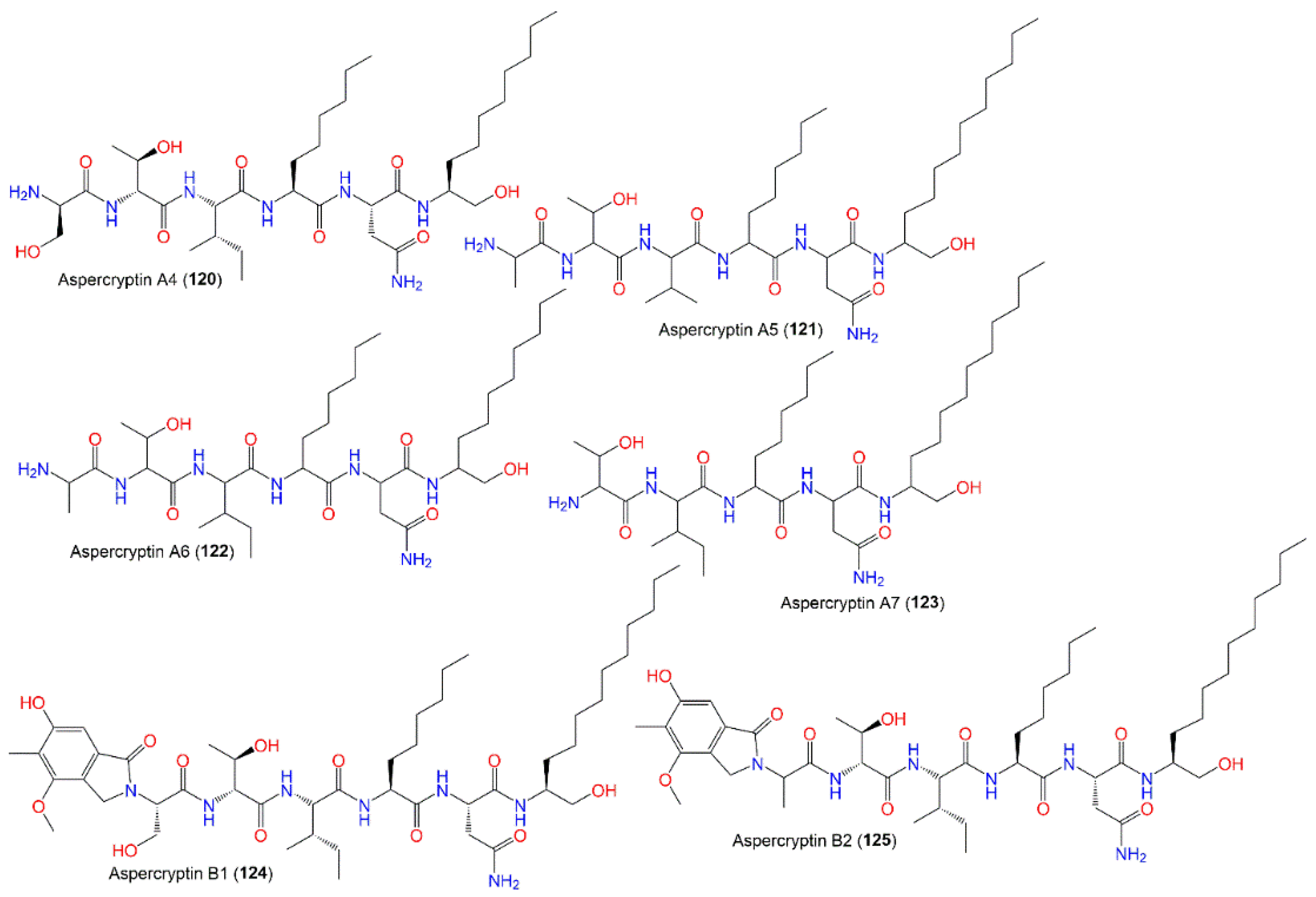
| Aspercryptin A4 (120) | 729 | C35H67N7O9 | Mutant strain cultured | [69] |
| Aspercryptin A5 (121) | 727 | C36H69N7O8 | Mutant strain cultured | [69] |
| Aspercryptin A6 (122) | 713 | C35H67N7O8 | Mutant strain cultured | [69] |
| Aspercryptin A7 (123) | 670 | C34H66N6O7 | Mutant strain cultured | [69] |
| Aspercryptin B1 (124) | 933 | C47H79N7O12 | Mutant strain cultured | [69] |
| Aspercryptin B2 (125) | 917 | C47H79N7O11 | Mutant strain cultured | [69] |
| Aspercryptin B3 (126) | 919 | C46H77N7O12 | Mutant strain cultured | [69] |
| Aspercryptin B4 (127) | 905 | C45H75N7O12 | Mutant strain cultured | [69] |
| Aspercryptin C1 (128) | 799 | C39H73N7O10 | Mutant strain cultured | [69] |
| Aspercryptin C2 (129) | 783 | C39H73N7O9 | Mutant strain cultured | [69] |
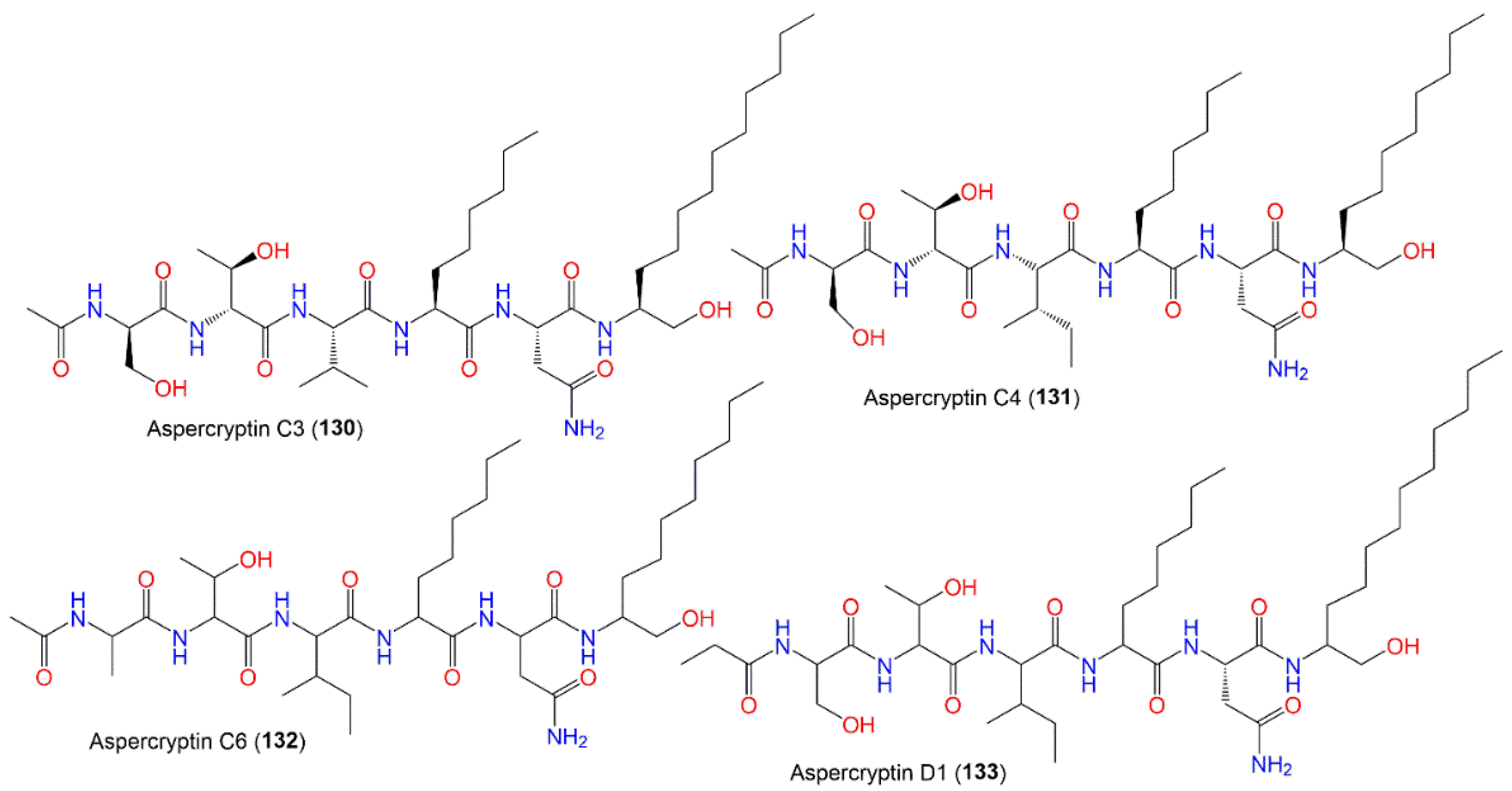
| Aspercryptin C3 (130) | 785 | C38H71N7O10 | Mutant strain cultured | [69] |
| Aspercryptin C4 (131) | 771 | C37H69N7O10 | Mutant strain cultured | [69] |
| Aspercryptin C6 (132) | 755 | C37H69N7O9 | Mutant strain cultured | [69] |
| Aspercryptin D1 (133) | 813 | C40H75N7O10 | Mutant strain cultured | [69] |
| Compound Name | Mol. Wt. | Mol. Formula | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
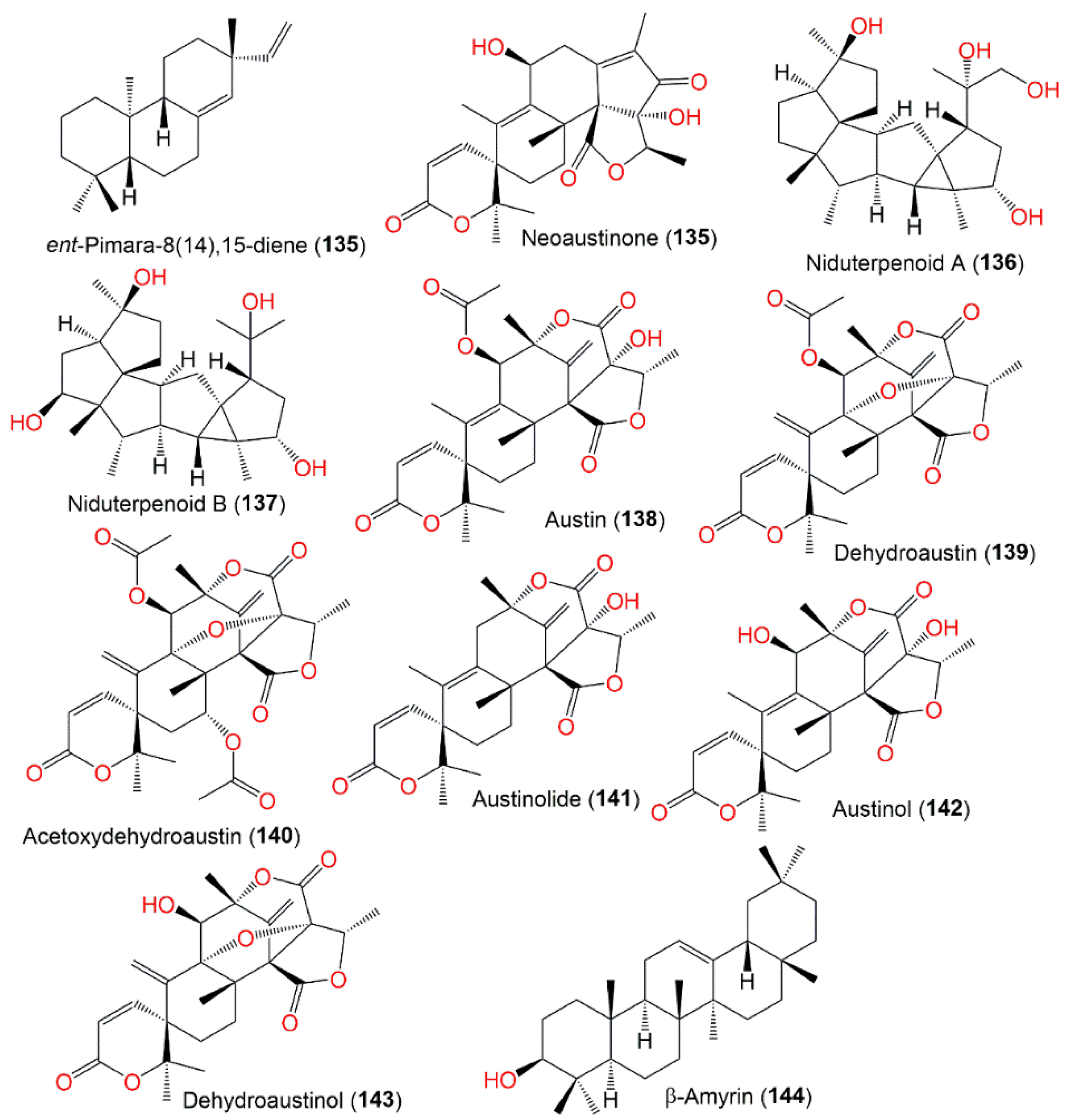
| ent-Pimara-8(14),15-diene (134) | 272 | C20H32 | Engineered cultured | [71] |
| Neoaustinone (135) | 442 | C25H30O7 | Cultured | [56] |
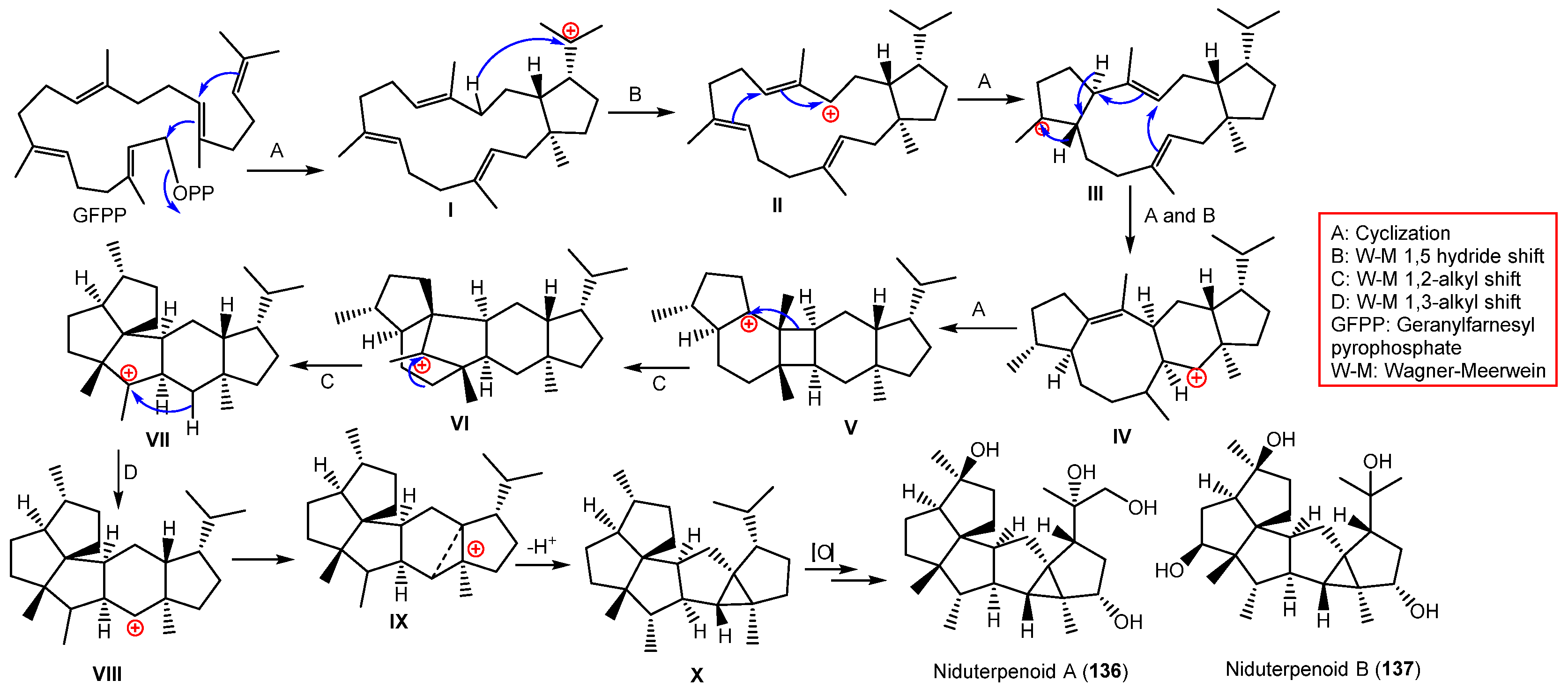
| Niduterpenoid A (136) | 404 | C25H40O4 | Whitmania pigra Whitman (Annelida, segmented worm, Hirudinidae), Hubei, China | [40] |
| Niduterpenoid B (137) | 404 | C25H40O4 | Whitmania pigra Whitman (Annelida, segmented worm, Hirudinidae), Hubei, China | [40] |
| Austin (138) | 500 | C27H32O9 | Cultured | [72] |
| Dehydroaustin (139) | 498 | C27H30O9 | Cultured | [72] |
| Acetoxydehydroaustin (140) | 556 | C29H32O11 | Cultured | [72] |
| Austinolide (141) | 442 | C25H30O7 | Cultured | [56] |
| Austinol (142) | 458 | C25H30O8 | Cultured | [44,49,72] |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| Dehydroaustinol (143) | 456 | C25H28O8 | Cultured | [44,49,72] |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| β-Amyrin (144) | 426 | C30H50O | Cultured | [73] |
| Compound Name | Mol. Wt. | Mol. Formula | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| (7Z,9Z,17Z)-,2α,3β-Dihydroxypregna-7,9,17(20)-trien-18-al (145) | 328 | C21H28O3 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| Ergosterol (146) | 396 | C28H44O | Cultured | [41,74] |
| - | - | Deep-sea sediment of the western Pacific Ocean, China | [58] | |
| - | - | Nyctanthes arbor-tristis (Oleaceae), Sabira | [53] | |
| - | - | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] | |
| Lanosterol (147) | 426 | C30H50O | Cultured | [74] |
| Nidulanoid A (148) | 486 | C30H46O5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| Nidulanoid B (149) | 456 | C30H48O3 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| (17E,22E,24R)-3β,5α-Dihydroxyergosta-7,17,22-trien-6,16-dione (150) | 440 | C28H40O4 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| Poriferastane-3β,6α-diol (151) | 432 | C29H52O2 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| 3β,5α-Dihydroxy-(22E, 24R)-ergosta-7,22-dien-6-one (152) | 428 | C28H44O3 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| (22E,24R)-Ergosta-7,22-dien-3β,5α,6α-triol (153) | 430 | C28H46O3 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| 5α,8α-Epidoxyergosta-6,22-dien-3β-ol (154) | 428 | C28H44O3 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| - | - | Deep-sea sediment, western Pacific Ocean, China | [58] | |
| 5α,8α-Epidioxyergosta-6,9(11),22-trien-3β-ol (155) | 426 | C28H42O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| - | - | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47] | |
| (22E)-3β,4β,5α-Trihydroxyergosta-7,22-dien-6-one (156) | 444 | C28H44O4 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Qichun city, Hubei, China | [41] |
| 5α,6α-Epoxy-3β-hydroxy-(22E)-ergosta-8(14),22-dien-7-one (157) | 426 | C28H42O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 3β,5α,9α-Trihydroxyergosta-7,22-diene-6-one (158) | 444 | C28H44O4 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 3β,5α-Dihydroxy-6β-acetoxy-ergosta-7,22-diene (159) | 458 | C30H50O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 3β,5α,6β,22E-Ergosta-7,22-diene-3,5,6-triol 6-palmitate (160) | 668 | C44H76O4 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| Nuatigenin (161) | 430 | C27H42O4 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 1-Dehydronuatigenone (162) | 426 | C27H38O4 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 1-Dehydroisonuatigenone (163) | 426 | C27H38O4 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 3β,15α-Dihydroxyl-(22E,24R)-ergosta-5,8(14),22-trien-7-one (164) | 426 | C28H42O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| 3β,15β-Dihydroxyl-(22E,24R)-ergosta-5,8(14),22-trien-7-one (165) | 426 | C28H42O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| β-Sitosterol-3-O-β-D-glucoside (166) | 576 | C35H60O6 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Ergosterol peroxide (167) | 428 | C28H44O3 | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47] |
| Compound Name | Mol. Wt. | Mol. Formula | Host (Part, Family) | Ref. |
|---|---|---|---|---|
| Nidulol (168) | 194 | C10H10O4 | Cultured | [51] |
| Porriolide (169) | 194 | C10H10O4 | Cultured | [57] |
| 3-Methoxyporriolide (170) | 224 | C11H12O5 | Cultured | [57] |
| 7-Methoxyporriolide (171) | 224 | C11H12O5 | Cultured | [57] |
| 3-Carboxy-2,4-diphenyl-but-2-enoic anhydride (172) | 264 | C17H12O3 | Cultured | [75] |
| Microperfuranone (173) | 266 | C17H14O3 | Green alga, Sardinia, Italy, Mediterranean Sea | [43] |
| - | - | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [52] | |
| - | - | Cultured | [57] | |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| 9-Hydroxymicroperfuranone (174) | 282 | C17H14O4 | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [52] |
| - | - | Deep-sea sediment, South China Sea | [31] | |
| Helvafuranone (175) | 298 | C17H14O5 | Soil, Hell Valley, Noboribetsu, Hokkaido, Japan | [52] |
| Asperfuranone (176) | 332 | C19H24O5 | Mutant strain cultured | [76] |
| Cichorine intermediate (177) | 196 | C10H12O4 | Cultured | [56] |
| (4S,5R)-4-Hydroxy-5-methylfuran-2-one (178) | 132 | C5H8O4 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Asperlin (U-13,933) (179) | 212 | C10H12O5 | Cultured | [75,77,78] |
| Chloro-asperlin (180) | 248 | C10H13ClO5 | Cultured | [77] |
| Versiol (181) | 262 | C16H22O3 | Deep-sea sediment, western Pacific Ocean, China | [58] |
| PsiAα (182) | 294 | C18H30O3 | Cultured | [79] |
| PsiAβ (183) | 296 | C18H32O3 | Cultured | [79] |
| Trichodermatide B (184) | 264 | C16H24O3 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Koninginin A (185) | 284 | C16H28O4 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Koninginin E (186) | 282 | C16H26O4 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Koninginin H (187) | 298 | C16H26O5 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Asperlinol (188) | 230 | C10H14O6 | Cultured | [77] |
| Compound Name/Chemical Class | Mol. Wt. | Mol. Formula | Host (Part, Family) | Ref. |
|---|---|---|---|---|
| Polyketides | ||||
| Curvularin (189) | 292 | C16H20O5 | Deep-sea sediment, South China Sea | [31] |
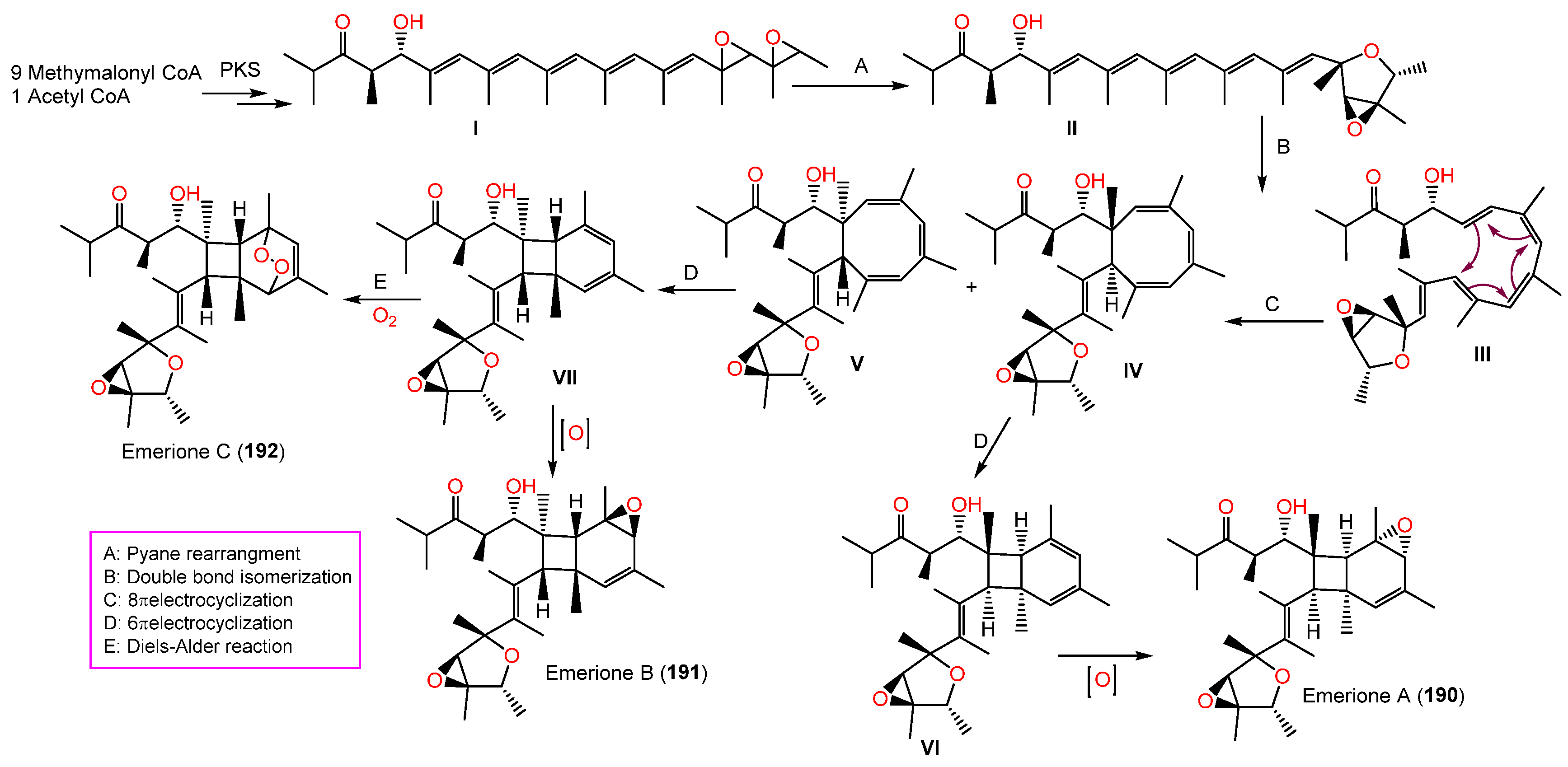
| Emerione A (190) | 472 | C29H44O5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [80] |
| Emerione B (191) | 472 | C29H44O5 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [80] |
| Emerione C (192) | 488 | C29H44O6 | Whitmania pigra (Annelida, segmented worm, Hirudinidae), Hubei, China | [80] |
| Citrantifidiol (193) | 240 | C15H28O2 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Glycerides | ||||
| Gingerglycolipid B (194) | 678 | C33H58O14 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| (2S)-bis[9Z,12Z]-1-O, 2- O-Dilinoleoyl-3-O-[α-D-galactopyranosyl-(1″→6′) β-D-galactopyranosyl]glycerol (195) | 940 | C51H88O15 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| (2S)-bis[9Z,12Z]-1-O, 2-O-dilinoleoyl-3-O-β-D-galactopyranosylglycerol (196) | 778 | C45H78O10 | Piece of orange peel (Rutaceae), Tifton, Georgia | [68] |
| Compound Name | Mol. Wt. | Mol. Formula | Host (Part, Family) | Ref. |
|---|---|---|---|---|
| Aspernidine D (197) | 386 | C24H34O4 | Cultured | [49] |
| Aspernidine E (198) | 402 | C24H34O5 | Cultured | [49] |
| Asperbenzaldehyde (199) | 316 | C19H24O4 | Cultured | [54] |
| 3-Methylorsellinic acid (200) | 182 | C9H10O4 | Cultured | [56] |
| O-Methyl-3-methylorsellinaldehyde dimer (201) | 360 | C20H24O6 | Cultured | [56] |
| 6-((S,3E,5E)-5,7-Dimethyl-2-oxonona-3,5-dien-1-yl)-2,4-dihydroxy- 3-methylbenzaldehyde (202) | 316 | C19H24O4 | Mutant strain cultured | [42] |
| Lecanoric acid (203) | 318 | C16H14O7 | Deep-sea sediment, South China Sea | [31] |
| Myristic acid (204) | 228 | C14H28O2 | Turbinaria elatensis (Brown alga, Phaeophyceae), Egyptian Red Sea, Ras Mohamed, South Sina, Egypt | [47] |
| Atrochrysone (205) | 274 | C15H14O5 | Cultured | [56] |
| Asperoxide A (206) | 234 | C12H26O4 | Deep-sea sediment, South China Sea | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghazawi, K.F.; Fatani, S.A.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential. Fermentation 2023, 9, 325. https://doi.org/10.3390/fermentation9040325
Ghazawi KF, Fatani SA, Mohamed SGA, Mohamed GA, Ibrahim SRM. Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential. Fermentation. 2023; 9(4):325. https://doi.org/10.3390/fermentation9040325
Chicago/Turabian StyleGhazawi, Kholoud F., Sarah A. Fatani, Shaimaa G. A. Mohamed, Gamal A. Mohamed, and Sabrin R. M. Ibrahim. 2023. "Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential" Fermentation 9, no. 4: 325. https://doi.org/10.3390/fermentation9040325
APA StyleGhazawi, K. F., Fatani, S. A., Mohamed, S. G. A., Mohamed, G. A., & Ibrahim, S. R. M. (2023). Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential. Fermentation, 9(4), 325. https://doi.org/10.3390/fermentation9040325