
Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review


Abstract
:1. Introduction
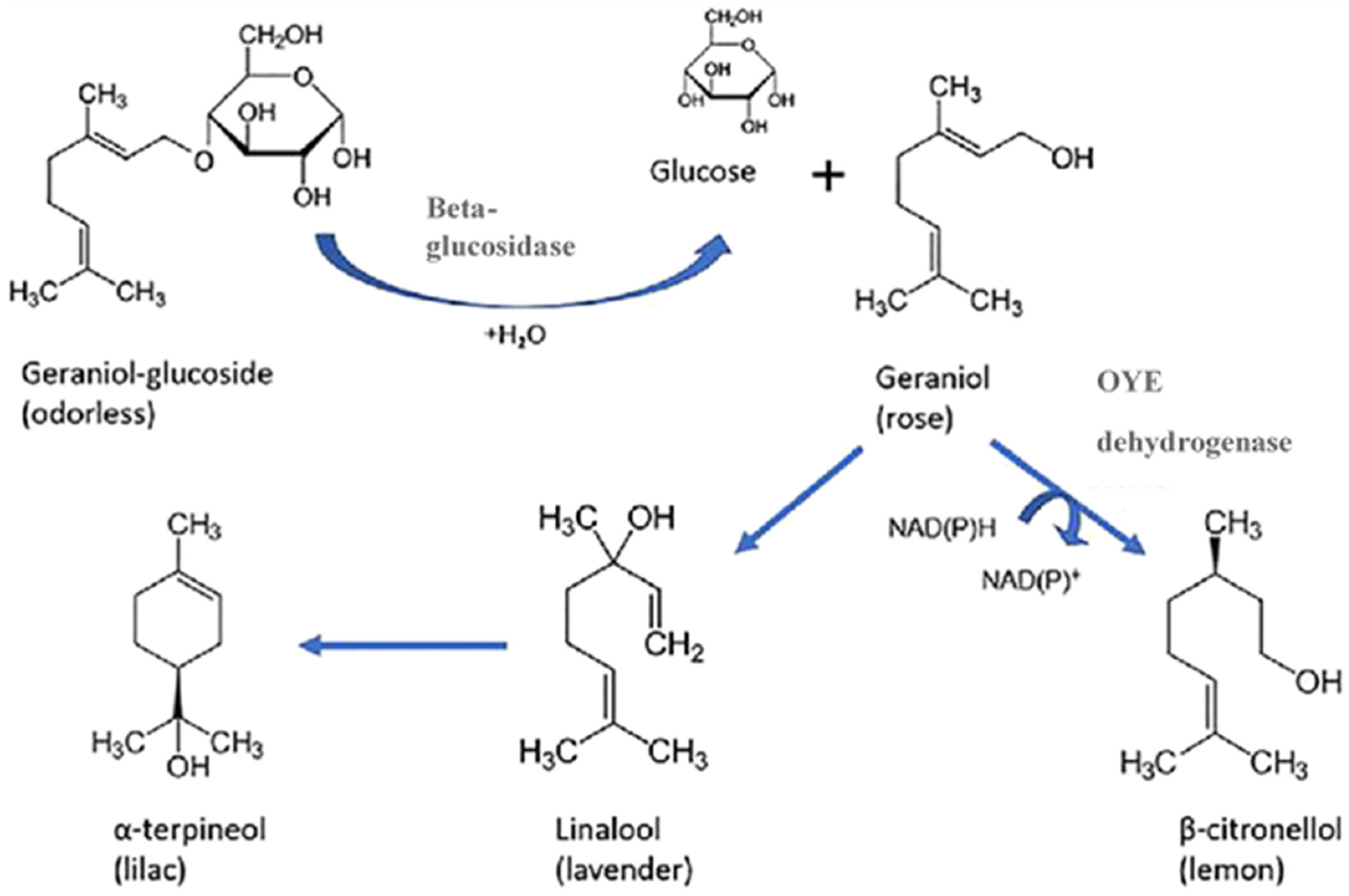
2. Yeasts Enzymatic Activity for the Release of Aromatic Compound from Precursors
3. The Yeasts
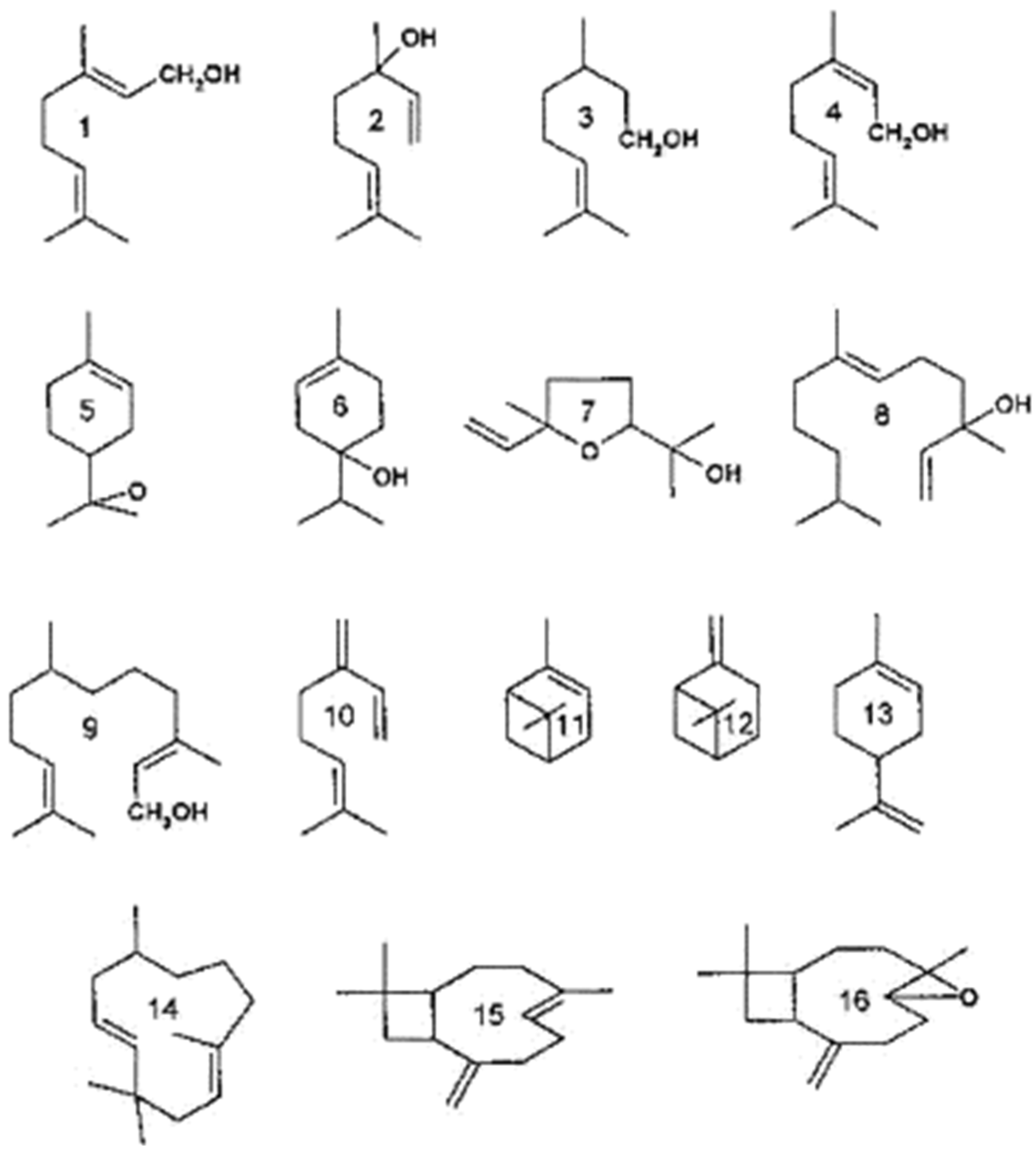
4. The Hops
5. Biotransformations during Beer Production
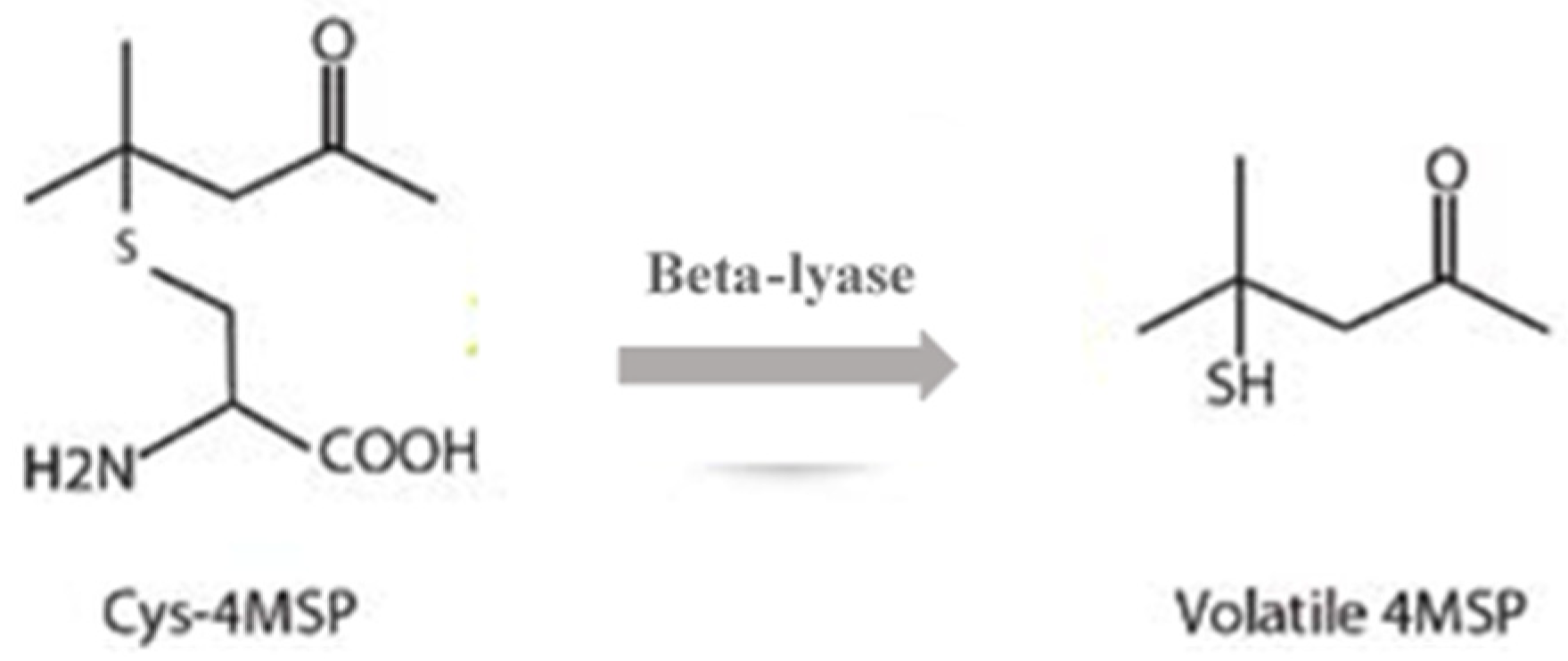
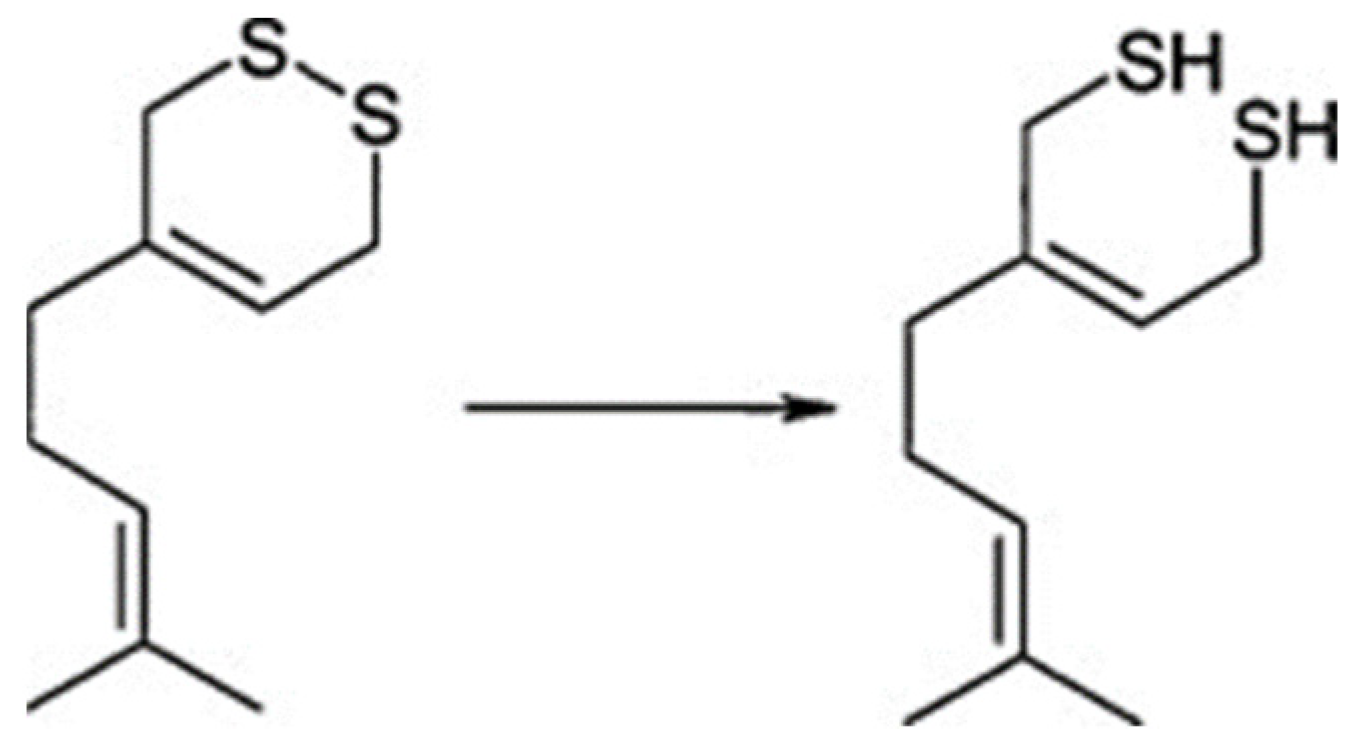
6. Other Reactions on Molecules Provided by Hop
7. Work in Progress and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Svedlund, N.; Evering, S.; Gibson, B.; Krogerus, K. Fruits of their labour: Biotransformation reactions of yeasts during brewery fermentation. Appl. Microbiol. Biotechnol. 2022, 106, 4929–4944. [Google Scholar] [CrossRef] [PubMed]
- Kind, C.; Kaiser, T. Heat, hops, Hallertau: Exploring implications of climate change for the German beer sector. In The Geography of Beer: Culture and Economics; Hoalst-Pullen, N., Patterson, M.W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 103–111. [Google Scholar]
- Verzele, M.; De Keukeleire, D. Chemistry and Analysis of Hop and Beer Bitter Acids; Elsevier Science: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Jaskula, B.; Kafarski, P.; Aerts, G.; De Cooman, L. A Kinetic Study on the Isomerization of Hop α-Acids. J. Agric. Food Chem. 2008, 56, 6408–6415. [Google Scholar] [CrossRef] [PubMed]
- Biendl, M.; Schmidt, C.; Maye, J.P.; Smith, R. New England IPA—The hop aroma champion of beers. MBAA Tech. Q. 2021, 58, 38–42. [Google Scholar]
- De Cooman, L.; Aerts, G.; Van Opstaele, F.; Goiris, K.; Syryn, E.; De Rouck, G.; De Ridder, M.; De Keukeleire, D. New trends in advanced hopping—Part 1: Application of pre-isomerised hop extracts. Cerevisia 2004, 29, 36–46. [Google Scholar]
- Lam, K.C.; Foster, R.T.; Deinzer, M.L. Aging of hops and their contribution to beer flavor. J. Agric. Food Chem. 1986, 34, 763–770. [Google Scholar] [CrossRef]
- Kishimoto, T.; Wanikawa, A.; Kagami, N.; Kawatsura, K. Analysis of Hop-Derived Terpenoids in Beer and Evaluation of Their Behavior Using the Stir Bar−Sorptive Extraction Method with GC-MS. J. Agric. Food Chem. 2005, 53, 4701–4707. [Google Scholar] [CrossRef]
- Steinhaus, M.; Schieberle, P. Transfer of the potent hop odorants linalool, geraniol and 4-methyl-4-sulfanyl-2-pentanone from hops into beer. In Proceedings of the 31st European Brewery Convention Congress, Venice, 2007 Fachverlag Hans Carl, Nürnberg, Germany, 6–10 May 2007; pp. 1004–1011. [Google Scholar]
- Steinhaus, M.; Wilhelm, W.; Schieberle, P. Comparison of the most odor-active volatiles in different hop varieties by application of a comparative aroma extract dilution analysis. Eur. Food Res. Technol. 2007, 226, 45–55. [Google Scholar] [CrossRef]
- Van Opstaele, F.; De Rouck, G.; De Clippeleer, J.; Aerts, G.; De Cooman, L. Analytical and Sensory Assessment of Hoppy Aroma and Bitterness of Conventionally Hopped and Advanced Hopped Pilsner Beers. J. Inst. Brew. 2010, 116, 445–458. [Google Scholar] [CrossRef]
- Sarry, J.-E.; Günata, Z. Plant and microbial glycoside hydrolases: Volatile release from glycosidic aroma precursors. Food Chem. 2004, 87, 509–521. [Google Scholar] [CrossRef]
- Daenen, L.; Saison, D.; Sterckx, F.; Delvaux, F.; Verachtert, H.; Derdelinckx, G. Screening and evaluation of the glucoside hydrolase activity in Saccharomyces and Brettanomyces brewing yeasts. J. Appl. Microbiol. 2007, 104, 478–488. [Google Scholar] [CrossRef]
- Goldstein, H.; Ting, P.; Navarro, A.; Ryder, D. Water-soluble hop flavor precursors and their role in beer flavour. In Proceedings of the 27th Congress of the European Brewery Convention, Cannes, France, 1999; IRL Press: Oxford, UK, 1999; pp. 53–62. [Google Scholar]
- Takoi, K.; Itoga, Y.; Takayanagi, J.; Matsumoto, I.; Nakayama, Y. Control of hop aroma impression of beer with blend-hopping using geraniol-rich hop and new hypothesis of synergy among hop-derived flavour compounds. Brew. Sci. 2016, 69, 85–93. [Google Scholar]
- Lafontaine, S.; Caffrey, A.; Dailey, J.; Varnum, S.; Hale, A.; Eichler, B.; Dennenlöhr, J.; Schubert, C.; Knoke, L.; Lerno, L.; et al. Evaluation of variety, maturity, and farm on the concentrations of monoterpene diglycosides and hop volatile/nonvolatile composition in five humulus lupulus cultivars. J. Agric. Food Chem. 2021, 69, 4356–4370. [Google Scholar] [CrossRef] [PubMed]
- Kollmannsberger, H.; Biendl, M.; Nitz, S. Occurrence of glycosidically bound flavor compounds in hops, hop products and beer. Monatsschr. Brauwiss. 2006, 5/6, 83–89. [Google Scholar]
- Biendl, M.; Kollmannsberger, H.; Nitz, S. Occurrence of glycosidically bound flavor compounds in different hop products. In Proceedings of the 29th Congress of the European Brewery Convention, Dublin, CD-ROM, Contribution 21, 2003; Fachverlag Hans Carl: Nürnberg, Germany, 2003; pp. 1–6. [Google Scholar]
- Colomer, M.S.; Funch, B.; Solodovnikova, N.; Hobley, T.J.; Förster, J. Biotransformation of hop derived compounds by Brettanomyces yeast strains. J. Inst. Brew. 2020, 126, 280–288. [Google Scholar] [CrossRef]
- Lloyd, N.D.R.; Capone, D.L.; Ugliano, M.; Taylor, D.K.; Skouroumounis, G.K.; Sefton, M.A.; Elsey, G.M. Formation of Damascenone under both Commercial and Model Fermentation Conditions. J. Agric. Food Chem. 2011, 59, 1338–1343. [Google Scholar] [CrossRef]
- Takoi, K.; Itoga, Y.; Koie, K.; Kosugi, T.; Shimase, M.; Katayama, Y.; Nakayama, Y.; Watari, J. The Contribution of Geraniol Metabolism to the Citrus Flavour of Beer: Synergy of Geraniol and β-Citronellol Under Coexistence with Excess Linalool. J. Inst. Brew. 2010, 116, 251–260. [Google Scholar] [CrossRef]
- Suggett, A.; Moir, M.; Seaton, J.C. The role of sulphur compounds in hop flavour. In Proceedings of the 17th European Brewery Convention Congress, Berlin, Germany, 1 December 1979; pp. 79–89. [Google Scholar]
- Lermusieau, G.; Collin, S. Volatile Sulfur Compounds in Hops and Residual Concentrations in Beer—A Review. J. Am. Soc. Brew. Chem. 2003, 61, 109–113. [Google Scholar] [CrossRef]
- Capone, D.L.; Ristic, R.; Pardon, K.H.; Jeffery, D.W. Simple Quantitative Determination of Potent Thiols at Ultratrace Levels in Wine by Derivatization and High-Performance Liquid Chromatography–Tandem Mass Spectrometry (HPLC-MS/MS) Analysis. Anal. Chem. 2015, 87, 1226–1231. [Google Scholar] [CrossRef]
- Dennenlöhr, J.; Thörner, S.; Rettberg, N. Analysis of Hop-Derived Thiols in Beer Using On-Fiber Derivatization in Combination with HS-SPME and GC-MS/MS. J. Agric. Food Chem. 2020, 68, 15036–15047. [Google Scholar] [CrossRef]
- Roland, A.; Viel, C.; Reillon, F.; Delpech, S.; Boivin, P.; Schneider, R.; Dagan, L. First identification and quantification of glutathionylated and cysteinylated precursors of 3-mercaptohexan-1-ol and 4-methyl-4-mercaptopentan-2-one in hops (Humulus lupulus). Flavour Fragr. J. 2016, 31, 455–463. [Google Scholar] [CrossRef]
- Chenot, C.; Haest, S.; Robiette, R.; Collin, S. Thiol S-Conjugate Profiles: A Comparative Investigation on Dual Hop and Grape Must with Focus on Sulfanylalkyl Aldehydes and Acetates Adducts. J. Am. Soc. Brew. Chem. 2022, 81, 23–32. [Google Scholar] [CrossRef]
- Howell, K.S.; Klein, M.; Swiegers, J.H.; Hayasaka, Y.; Elsey, G.M.; Fleet, G.H.; Høj, P.B.; Pretorius, I.S.; de Barros Lopes, M.A. Genetic Determinants of Volatile-Thiol Release by Saccharomyces cerevisiae during Wine Fermentation. Appl. Environ. Microbiol. 2005, 71, 5420–5426. [Google Scholar] [CrossRef] [Green Version]
- Thibon, C.; Marullo, P.; Claisse, O.; Cullin, C.; Dubourdieu, D.; Tominaga, T. Nitrogen catabolic repression controls the release of volatile thiols by Saccharomyces cerevisiae during wine fermentation. FEMS Yeast Res. 2008, 8, 1076–1086. [Google Scholar] [CrossRef] [Green Version]
- Roncoroni, M.; Santiago, M.; Hooks, D.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Cordente, A.G.; Williams, S.J.; Capone, D.L.; Jitjaroen, W.; Menz, I.R.; Curtin, C.; Anderson, P.A. Engineering Saccharomyces cerevisiae To Release 3-Mercaptohexan-1-ol during Fermentation through Overexpression of an S. cerevisiae Gene, STR3, for Improvement of Wine Aroma. Appl. Environ. Microbiol. 2011, 77, 3626–3632. [Google Scholar] [CrossRef] [Green Version]
- Subileau, M.; Schneider, R.; Salmon, J.-M.; Degryse, E. Nitrogen catabolite repression modulates the production of aromatic thiols characteristic of Sauvignon Blanc at the level of precursor transport. FEMS Yeast Res. 2008, 8, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Krogerus, K.; Fletcher, E.; Rettberg, N.; Gibson, B.; Preiss, R. Efficient breeding of industrial brewing yeast strains using CRISPR/Cas9-aided mating-type switching. Appl. Microbiol. Biotechnol. 2021, 105, 8359–8376. [Google Scholar] [CrossRef] [PubMed]
- Nizet, S.; Gros, J.; Peeters, F.; Chaumont, S.; Robiette, R.; Collin, S. First Evidence of the Production of Odorant Polyfunctional Thiols by Bottle Refermentation. J. Am. Soc. Brew. Chem. 2013, 71, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Michel, M.; Haslbeck, K.; Ampenberger, F.; Meier-Dörnberg, T.; Stretz, D.; Hutzler, M.; Coelhan, M.; Jacob, F.; Liu, Y. Screening of brewing yeast β-lyase activity and release of hop volatile thiols from precursors during fermentation. Brew. Sci. 2019, 72, 179–186. [Google Scholar]
- Darriet, P.; Tominaga, T.; Lavigne, V.; Boidron, J.-N.; Dubourdieu, D. Identification of a powerful aromatic component of Vitis vinifera L. var. sauvignon wines: 4-mercapto-4-methylpentan-2-one. Flavour Fragr. J. 1995, 10, 385–392. [Google Scholar] [CrossRef]
- Tominaga, T.; Furrer, A.; Henry, R.; Dubourdieu, D. Identification of new volatile thiols in the aroma of Vitis vinifera L. var. Sauvignon Blanc Wines. Flavour Fragr. J. 1998, 13, 159–162. [Google Scholar] [CrossRef]
- Kishimoto, T.; Wanikawa, A.; Kono, K.; Shibata, K. Comparison of the Odor-Active Compounds in Unhopped Beer and Beers Hopped with Different Hop Varieties. J. Agric. Food Chem. 2006, 54, 8855–8861. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, E.; Shinkaruk, S.; Tominaga, T.; Bennetau, B.; Frérot, E.; Dubourdieu, D. Odorous Impact of Volatile Thiols on the Aroma of Young Botrytized Sweet Wines: Identification and Quantification of New Sulfanyl Alcohols. J. Agric. Food Chem. 2007, 55, 1437–1444. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T.; Kobayashi, M.; Yako, N.; Iida, A.; Wanikawa, A. Comparison of 4-Mercapto-4-methylpentan-2-one Contents in Hop Cultivars from Different Growing Regions. J. Agric. Food Chem. 2008, 56, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Takoi, K.; Degueil, M.; Shinkaruk, S.; Thibon, C.; Maeda, K.; Ito, K.; Bennetau, B.; Dubourdieu, D.; Tominaga, T. Identification and characteristics of new volatile thiols derived from the hop (Humulus lupulus L.) cultivar Nelson Sauvin. J. Agric. Food Chem. 2009, 57, 2493–2502. [Google Scholar] [CrossRef]
- Iizuka-Furukawa, S.; Isogai, A.; Kusaka, K.; Fujii, T.; Wakai, Y. Identification of 4-mercapto-4-methylpentan-2-one as the characteristic aroma of sake made from low-glutelin rice. J. Biosci. Bioeng. 2017, 123, 209–215. [Google Scholar] [CrossRef]
- Cibaka, M.-L.K.; Ferreira, C.S.; Decourrière, L.; Lorenzo-Alonso, C.-J.; Bodart, E.; Collin, S. Dry Hopping with the Dual-Purpose Varieties Amarillo, Citra, Hallertau Blanc, Mosaic, and Sorachi Ace: Minor Contribution of Hop Terpenol Glucosides to Beer Flavors. J. Am. Soc. Brew. Chem. 2017, 75, 122–129. [Google Scholar] [CrossRef]
- Gijs, L.; Collin, S. Occurrence et voies de formation des arômes soufrés dans la bière 3. Les terpènes et hétérocycles soufrés. Cerevisia 2003, 28, 31–39. [Google Scholar]
- Zhang, P.; Zhang, R.; Sirisena, S.; Gan, R.; Fang, Z. Beta-glucosidase activity of wine yeasts and its impacts on wine volatiles and phenolics: A mini-review. Food Microbiol. 2021, 100, 103859. [Google Scholar] [CrossRef]
- Gamero, A.; Manzanares, P.; Querol, A.; Belloch, C. Monoterpene alcohols release and bioconversion by Saccharomyces species and hybrids. Int. J. Food Microbiol. 2011, 145, 92–97. [Google Scholar] [CrossRef]
- Masneuf-Pomarède, I.; Murat, M.-L.; Naumov, G.I.; Tominaga, T.; Dubourdieu, D. Hybrids Saccharomyces cerevisiae X Saccharomyces bayanus var. uvarum having a high liberating ability of some sulfur varietal aromas of Vitis vinifera Sauvignon blanc wines. J. Int. Sci. Vigne Vin. 2002, 36, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Fia, G.; Giovani, G.; Rosi, I. Study of β-glucosidase production by wine-related yeasts during alcoholic fermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 2005, 99, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Bonciani, T.; De Vero, L.; Giannuzzi, E.; Verspohl, A.; Giudici, P. Qualitative and quantitative screening of the β -glucosidase activity in Saccharomyces cerevisiae and Saccharomyces uvarum strains isolated from refrigerated must. Lett. Appl. Microbiol. 2018, 67, 72–78. [Google Scholar] [CrossRef]
- Escribano, R.; Gonza´lez-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; SantamarÍa, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Nebreda, A.R.; Villa, T.G.; Villanueva, J.R.; Delrey, F. Cloning of genes related to exo-beta-glucanase production in Saccharomyces cerevisiae—Characterization of an exo-beta-glucanase structural gene. Gene 1986, 47, 245–259. [Google Scholar] [CrossRef]
- Mateo, J.J.; DiStefano, R. Description of the betaglucosidase activity of wine yeasts. Food Microbiol. 1997, 14, 583–591. [Google Scholar] [CrossRef]
- Gil, J.V.; Manzanares, P.; Genovés, S.; Vallés, S.; González-Candelas, L. Over-production of the major exoglucanase of Saccharomyces cerevisiae leads to an increase in the aroma of wine. Int. J. Food Microbiol. 2005, 103, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Olivero, I.; Hernandez, L.M.; Larriba, G. Regulation of beta-exoglucanase activity production by Saccharomyces cerevisiae in batch and continuous culture. Arch. Microbiol. 1985, 143, 143–146. [Google Scholar] [CrossRef]
- Suzuki, K.; Yabe, T.; Maruyama, Y.; Abe, K.; Nakajima, T. Characterization of Recombinant Yeast Exo-β-1,3-Glucanase (Exg 1p) Expressed in Escherichia coli Cells. Biosci. Biotechnol. Biochem. 2001, 65, 1310–1314. [Google Scholar] [CrossRef]
- Cid, V.J.; Duran, A.; Delrey, F.; Snyder, M.P.; Nombela, C.; Sanchez, M. Molecular-basis of cell integrity and morphogenesis in Saccharomyces cerevisiae. Microbiol. Rev. 1995, 59, 345–386. [Google Scholar] [CrossRef]
- Gunata, Y.; Bayonove, C.; Baumes, R.; Cordonnier, R. The aroma of grapes I. Extraction and determination of free and glycosidically bound fractions of some grape aroma components. J. Chromatogr. A 1985, 331, 83–90. [Google Scholar] [CrossRef]
- Ugliano, M.; Bartowsky, E.J.; McCarthy, J.; Moio, L.; Henschke, P.A. Hydrolysis and Transformation of Grape Glycosidically Bound Volatile Compounds during Fermentation with Three Saccharomyces Yeast Strains. J. Agric. Food Chem. 2006, 54, 6322–6331. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Iserentant, D.; Malcorps, P.; Derdelinckx, G.; Van Dijck, P.; Winderickx, J.; Pretorius, I.S.; Thevelein, J.M.; Delvaux, F.R. Glucose and sucrose: Hazardous fast-food for industrial yeast? Trends Biotechnol. 2004, 22, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Verachtert, H.; Dawoud, E. Microbiology of lambic- type beers. J. Appl. Bacteriol. 1984, 57, R11–R12. [Google Scholar]
- Gondé, P.; Blondin, B.; Leclerc, M.; Ratomahenina, R.; Arnaud, A.; Galzy, P. Fermentation of Cellodextrins by Different Yeast Strains. Appl. Environ. Microbiol. 1984, 48, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spindler, D.D.; Wyman, C.E.; Grohmann, K.; Philippidis, G.P. Evaluation of the cellobiose-fermenting yeastBrettanomyces custersii in the simultaneous saccharification and fermentation of cellulose. Biotechnol. Lett. 1992, 14, 403–407. [Google Scholar] [CrossRef]
- Krogerus, K.; Rettberg, N.; Gibson, B. Increased volatile thiol release during beer fermentation using constructed interspecies yeast hybrids. Eur. Food Res. Technol. 2022, 249, 55–69. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Morrison-Whittle, P.; Goddard, M. Fungal diversity during fermentation correlates with thiol concentration in wine. Aust. J. Grape Wine Res. 2017, 24, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Dufour, M.; Zimmer, A.; Thibon, C.; Marullo, P. Enhancement of volatile thiol release of Saccharomyces cerevisiae strains using molecular breeding. Appl. Microbiol. Biotechnol. 2013, 97, 5893–5905. [Google Scholar] [CrossRef]
- Salmon, J.-M.; Barre, P. Improvement of Nitrogen Assimilation and Fermentation Kinetics under Enological Conditions by Derepression of Alternative Nitrogen-Assimilatory Pathways in an Industrial Saccharomyces cerevisiae Strain. Appl. Environ. Microbiol. 1998, 64, 3831–3837. [Google Scholar] [CrossRef] [Green Version]
- Pardo, E.; Rico, J.; Gil, J.V.; Orejas, M. De novo production of six key grape aroma monoterpenes by a geraniol synthase-engineered S. cerevisiae wine strain. Microb. Cell Factories 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.J.; Dickinson, J.R. Biotransformation of hop aroma terpenoids by ale and lager yeasts. FEMS Yeast Res. 2003, 3, 53–62. [Google Scholar] [CrossRef]
- Oswald, M.; Fischer, M.; Dirninger, N.; Karst, F. Monoterpenoid biosynthesis in Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maicas, S. The Role of Yeasts in Fermentation Processes. Microorganisms 2020, 8, 1142. [Google Scholar] [CrossRef]
- Methner, Y.; Hutzler, M.; Matoulková, D.; Jacob, F.; Michel, M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation 2019, 5, 101. [Google Scholar] [CrossRef] [Green Version]
- Ellis, D.J.; Kerr, E.D.; Schenk, G.; Schulz, B.L. Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages. Beverages 2022, 8, 41. [Google Scholar] [CrossRef]
- Michel, M.; Kopecka, J.; Meier-Dornberg, T.; Zarnkow, M.; Jacob, F.; Hutzler, M. Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast 2016, 33, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Tataridis, P.; Kanelis, A.; Logotetis, S.; Nerancis, E. Use of non-saccharomyces Torulaspora delbrueckii yeast strains in winemaking and brewing. Zb. Matic. Srp. Přír. Nauk. 2013, 113, 415–426. [Google Scholar] [CrossRef]
- Salvadó, Z.; Arroyo-López, F.N.; Guillamón, J.M.; Salazar, G.; Querol, A.; Barrio, E. Temperature adaptation markedly determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef] [Green Version]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovativem brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- King, A.; Richard Dickinson, J. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Postigo, V.; Sánchez, A.; Cabellos, J.M.; Arroyo, T. New Approaches for the Fermentation of Beer: Non-Saccharomyces Yeasts from Wine. Fermentation 2022, 8, 280. [Google Scholar] [CrossRef]
- Sharpe, F.R.; Laws, D.R.J. The essential oil of hops a review. J. Inst. Brew. 1981, 87, 96–107. [Google Scholar] [CrossRef]
- Eriksen, R.L.; Padgitt-Cobb, L.K.; Townsend, M.S.; Henning, J.A. Gene expression for secondary metabolite biosynthesis in hop (Humulus lupulus L.) leaf lupulin glands exposed to heat and low-water stress. Sci. Rep. 2021, 11, 5138. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K.; Garbe, D.; Surburg, H. Common Fragrance and Flavor Materials: Preparation and Uses, 2nd ed.; VCH Publishers: New York, NY, USA, 1990. [Google Scholar]
- Nickerson, G.B.; Van Engel, L. Hop aroma component profile and the aroma unit. J. Am. Soc. Brew. Chem. 1992, 50, 77–81. [Google Scholar] [CrossRef]
- Eisenreich, W.; Sagner, S.; Zenk, M.H.; Bacher, A. Monoterpenoid essential oils are not of mevalanoid origin. Tetrahedron Lett. 1997, 38, 3889–3892. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Prediction of flavor differences between beers from their chemical composition. J. Agric. Food Chem. 1982, 30, 1009–1017. [Google Scholar] [CrossRef]
- Meilgaard, M.C.; Peppard, T.L. The flavor of beer. In Food Flavors Part B: The Flavor of Beverages; Morton, I.D., Macleod, A.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; pp. 99–170. [Google Scholar]
- Siebert, K.J. Sensory analysis of hop oil-derived compounds in beer: Flavor effects of individual compounds. Quality control. EBC Monograph 22. In Proceedings of the Symposium on Flavor Hops, Zoeterwoude, The Netherlands, 1994; Fachverlag Hans Carl: Nürnberg, Germany, 1994; pp. 198–212. [Google Scholar]
- Mitter, W.; Biendl, M.; Kalner, D. Behavior of hop-derived aroma substances during wort boiling. In EBC Monograph 31, Proceedings of the EBC Symposium on Flavor and Flavor Stability, Nancy, France, 2001; Fachverlag Hans Carl: Nürnberg, Germany, 2001; pp. 1–12, Contribution 31. [Google Scholar]
- Yang, X.; Lederer, C.; McDaniel, M.; Deinzer, M. Chemical analysis and sensory evaluation of hydrolysis products of humulene epoxides II and III. J. Agric. Food Chem. 1993, 41, 1300–1304. [Google Scholar] [CrossRef]
- Yang, X.; Lederer, C.; McDaniel, M.; Deinzer, M. Hydrolysis products of caryophyllene oxide in hops and beer. J. Agric. Food Chem. 1993, 41, 2082–2085. [Google Scholar] [CrossRef]
- Takoi, K.; Itoga, Y.; Koie, K.; Takayanagi, J.; Kaneko, T.; Watanabe, T.; Matsumoto, I.; Nomura, M. Systematic Analysis of Behaviour of Hop-Derived Monoterpene Alcohols during Fermentation and New Classification of Geraniol-Rich Flavour Hops. BrewingScience 2017, 70, 177–186. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, M.; Kishimoto, T.; Kobayashi, M.; Yako, N.; Iida, A.; Wanikawa, A.; Kitagawa, Y. Effects of Bordeaux Mixture (Copper Sulfate) Treatment on Blackcurrant/Muscat-Like Odors in Hops and Beer. J. Am. Soc. Brew. Chem. 2010, 68, 30–33. [Google Scholar] [CrossRef]
- Praet, T.; Van Opstaele, F.; Jaskula-Goiris, B.; Aerts, G.; De Cooman, L. Biotransformations of hop-derived aroma compounds by Saccharomyces cerevisiae upon fermentation. Cerevisia 2012, 36, 125–132. [Google Scholar] [CrossRef]
- Hauser, D.G.; Shellhammer, T.H. An overview of sustainability challenges in beer production, and the carbon footprint of hops production. MBAA Technical. Q. 2019, 56, 2–6. [Google Scholar]
- Gomes, F.D.O.; Guimarães, B.P.; Ceola, D.; Ghesti, G.F. Advances in dry hopping for industrial brewing: A review. Food Sci. Technol. 2022, 42, 1–8. [Google Scholar] [CrossRef]
- Kaltner, D.; Mitter, W. Changes in hop derived compounds during beer production and ageing. In Hop Flavor and Aroma- Proceedings of the 1st International Brewers Symposium; Shelhammer, T.H., Ed.; Master Brewers Association of the Americas: St. Paul, MN, USA, 2009; pp. 37–46. [Google Scholar]
- Lermusieau, G.; Bulens, M.; Collin, S. Use of GC−Olfactometry to Identify the Hop Aromatic Compounds in Beer. J. Agric. Food Chem. 2001, 49, 3867–3874. [Google Scholar] [CrossRef]
- Kaltner, D.; Thum, B.; Forster, C.; Back, W. Untersuchen zum Hopfenaroma in Pilsner Bieren bei Variation technologischer Parameter. Monatsschr. Brauwiss. 2001, 9–10, 199–205. [Google Scholar]
- Takoi, K.; Itoga, Y.; Takayanagi, J.; Kosugi, T.; Shioi, T.; Nakamura, T.; Watari, J. Screening of geraniol-rich flavor hop and interesting behavior of β-citronellol during fermentation under various hop-addition timings. J. Am. Soc. Brew. Chem. 2014, 72, 22–29. [Google Scholar] [CrossRef]
- Chenot, C.; de Chanvalon, E.T.; Janssens, P.; Collin, S. Modulation of the Sulfanylalkyl Acetate/Alcohol Ratio and Free Thiol Release from Cysteinylated and/or Glutathionylated Sulfanylalkyl Alcohols in Beer under Different Fermentation Conditions. J. Agric. Food Chem. 2021, 69, 6005–6012. [Google Scholar] [CrossRef] [PubMed]
- Koslitz, S.; Renaud, L.; Kohler, M.; Wüst, M. Stereoselective Formation of the Varietal Aroma Compound Rose Oxide during Alcoholic Fermentation. J. Agric. Food Chem. 2008, 56, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Moir, M. Hop aromatic compounds. In EBC Monograph 22, Proceedings of the Symposium on Hops, Zoeterwoude, The Netherlands, 1994; Fachverlag Hans Carl: Nürnberg, Germany, 1994; pp. 165–180. [Google Scholar]
- Nielsen, T.P. Character impact hop aroma compounds in ale. In Proceedings of the 1st International Brewers Symposium Hop Flavor and Aroma; Shelhammer, T.H., Ed.; Master Brewers Association of the Americas: St. Paul, MN, USA, 2009; pp. 59–77. [Google Scholar]
- Gramatica, P.; Manitto, P.; Ranzi, B.M.; Delbianco, A.; Francavilla, M. Stereospecific reduction of geraniol to (R)-(+)-citronellol bySaccharomyces cerevisiae. Cell Mol. Life Sci. 1982, 38, 775–776. [Google Scholar] [CrossRef]
- Dickinson, J.R. Carbon metabolism. In The Metabolism and Molecular Physiology of Saccharomyces Cerevisiae; Dickinson, J.R., Schweizer, M., Eds.; Taylor and Francis: London, UK; Philadelphia, PA, USA, 1999; pp. 23–55. [Google Scholar]
- Schopfer, L.M.; Massey, V. Old yellow enzyme. In A Study of Enzymes; Kuby, S.A., Ed.; CRC Press: Boston, MA, USA, 1991; pp. 247–269. [Google Scholar]
- Yuan, T.-T.; Chen, Q.-Q.; Zhao, P.-J.; Zeng, Y.; Liu, X.-Z.; Lu, S. Identification of enzymes responsible for the reduction of geraniol to citronellol. Nat. Prod. Bioprospecting 2011, 1, 108–111. [Google Scholar] [CrossRef] [Green Version]
- Tressl, R.; Kossa, M.; Köppler, H. Changes of aroma components during processing of hops. In EBC Monograph 16, Proceedings of the EBC Symposium on Hops, Freisin/Weihenstephan, Germany, 1987; Fachverlag Hans Carl: Nuremberg, Germany, 1987; pp. 116–119. [Google Scholar]
- Hughes, P. Beer flavor. In Beer—A Quality Perspective; Bamforth, C., Russell, I., Stewart, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 68–71. [Google Scholar]
- Khor, G.K.; Uzir, M.H. Saccharomyces cerevisiae: A potential stereospecific reduction tool for biotransformation of mono- and sesquiterpenoids. Yeast 2010, 28, 93–107. [Google Scholar] [CrossRef]
- Takoi, K.; Koie, K.; Itoga, Y.; Katayama, Y.; Shimase, M.; Nakayama, Y.; Watari, J. Biotransformation of Hop-Derived Monoterpene Alcohols by Lager Yeast and Their Contribution to the Flavor of Hopped Beer. J. Agric. Food Chem. 2010, 58, 5050–5058. [Google Scholar] [CrossRef]
- Peacock, V.E.; Deinzer, M.L. Chemistry of Hop Aroma in Beer. J. Am. Soc. Brew. Chem. 1981, 39, 136–141. [Google Scholar] [CrossRef]
- Forster, A.; Gahr, A.; Van Opstaele, F. On the transfer rate of geraniol with dry hopping. Brew. Sci. 2014, 67, 60–62. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
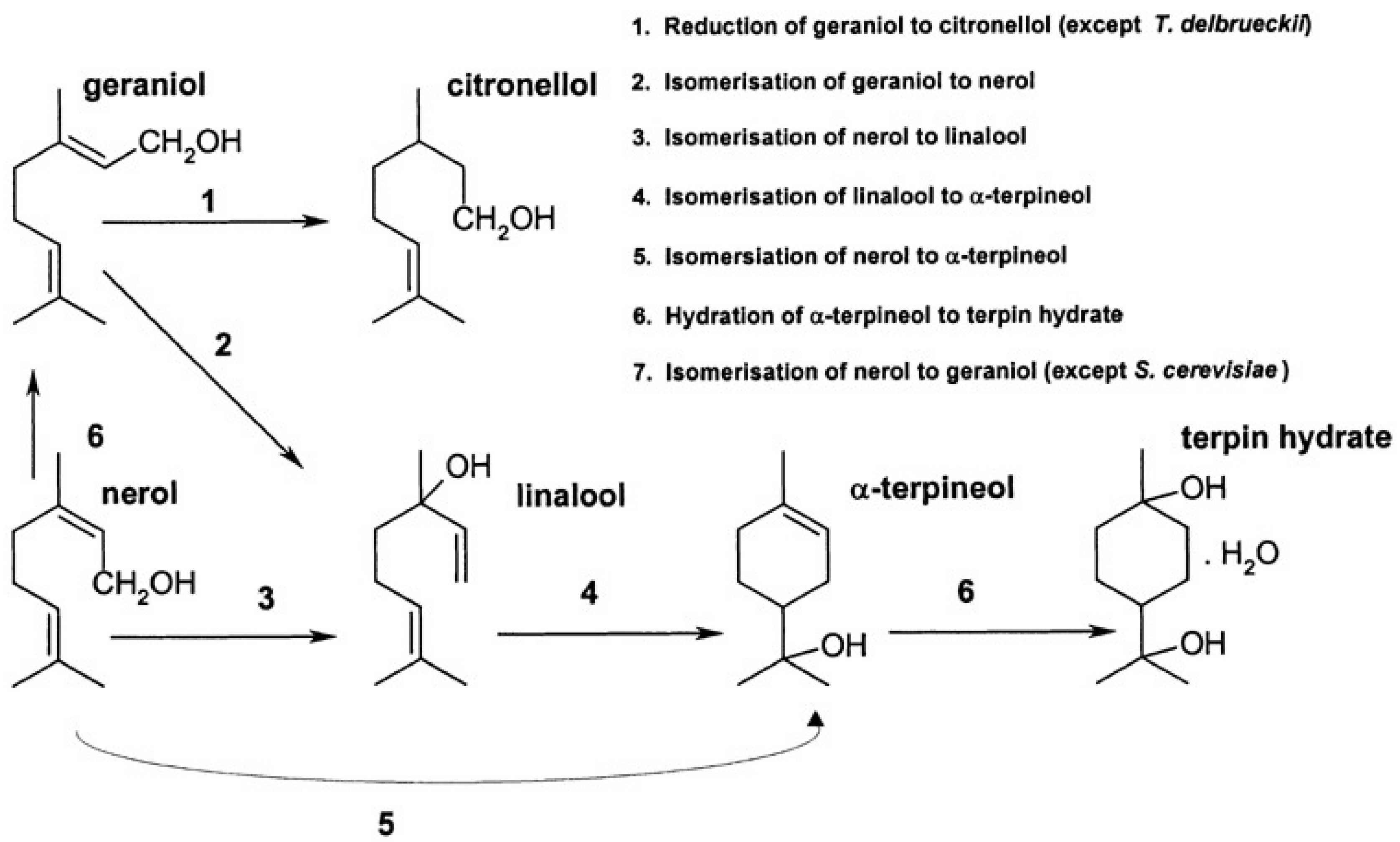{kind=link}
| Yeasts | β-Glucosidase Activity | Reference | β-Lyase Activity | Reference |
|---|---|---|---|---|
| Saccharomyces cerevisiae | - or +++ | [45] | + or ++ | [35] |
| Saccharomyces bayanus | + | [46] | ++ | [47] |
| Brettanomyces spp. | ++ or ++++ | [48] | - | [48] |
| Saccharomyces uvarum | ++ | [49] | ++ | [47] |
| Saccharomyces pastorianus | - | [13] | +++ | [35] |
| Torulaspora delbrueckii | - | [50] | ++ | [1,35] |
| Metschnikowia spp. | +++ | [50] | ++ | [1] |
| Lachancea | + | [50] | ++ | [1] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buiatti, S.; Tat, L.; Natolino, A.; Passaghe, P. Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review. Fermentation 2023, 9, 327. https://doi.org/10.3390/fermentation9040327
Buiatti S, Tat L, Natolino A, Passaghe P. Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review. Fermentation. 2023; 9(4):327. https://doi.org/10.3390/fermentation9040327
Chicago/Turabian StyleBuiatti, Stefano, Lara Tat, Andrea Natolino, and Paolo Passaghe. 2023. "Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review" Fermentation 9, no. 4: 327. https://doi.org/10.3390/fermentation9040327
APA StyleBuiatti, S., Tat, L., Natolino, A., & Passaghe, P. (2023). Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review. Fermentation, 9(4), 327. https://doi.org/10.3390/fermentation9040327