
Microbial Diversity Using a Metataxonomic Approach, Associated with Coffee Fermentation Processes in the Department of Quindío, Colombia

Abstract
:1. Introduction
2. Materials and Methods
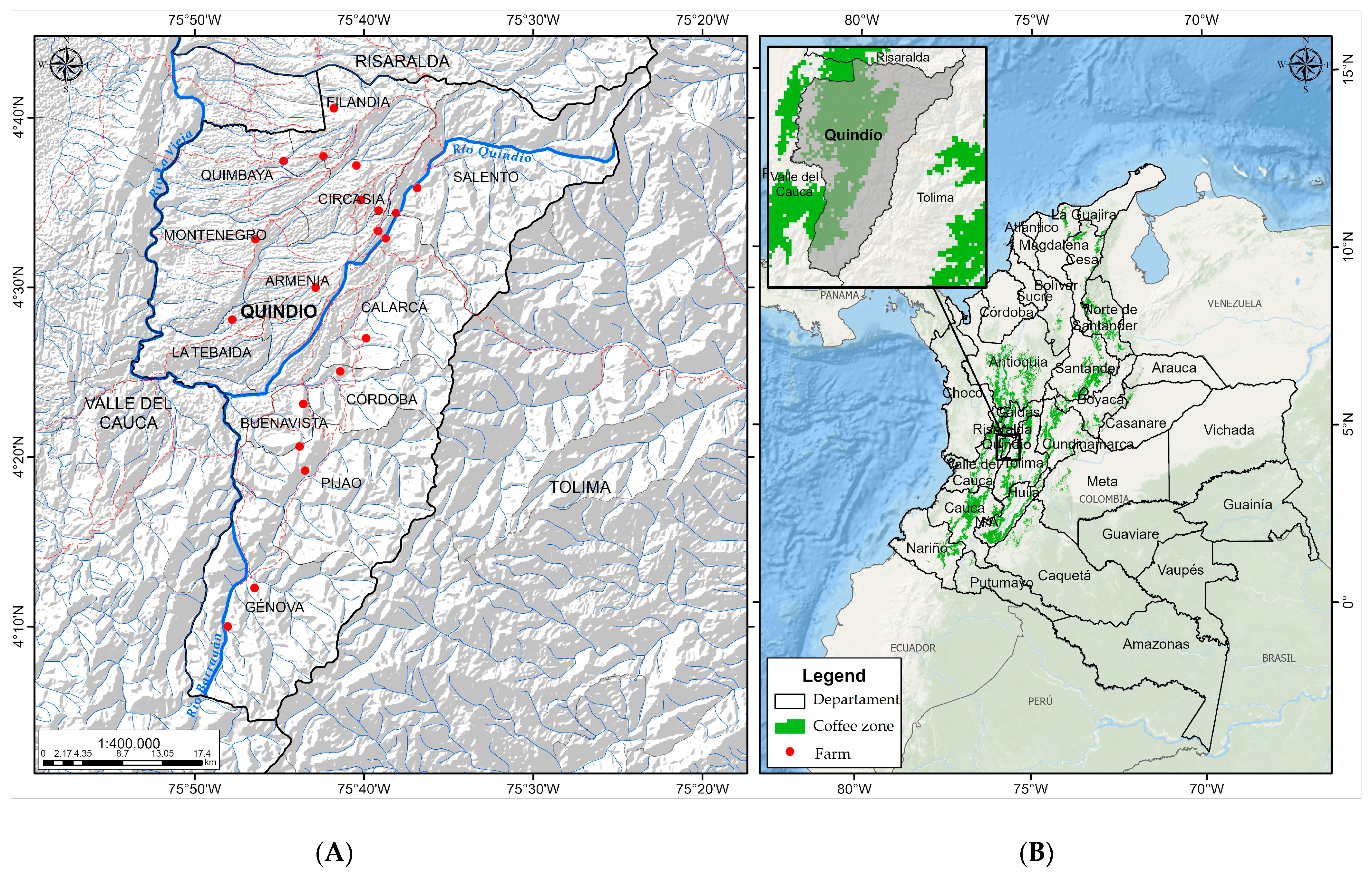
2.1. On-Farm Coffee Fermentation and Sampling
2.2. pH and Temperature Measurement
2.3. Coffee Beverage Sensory Quality
2.4. Volatile Organic Compound Determination
2.5. DNA Extraction and Quantification
2.6. Illumina MiSeq Amplicon Sequencing of Bacterial 16S rRNA Gene and of Fungal ITS Region
2.7. Microbiome and Statistical Analysis
3. Results
3.1. Coffee Fermentation Processes and Sample Characteristics
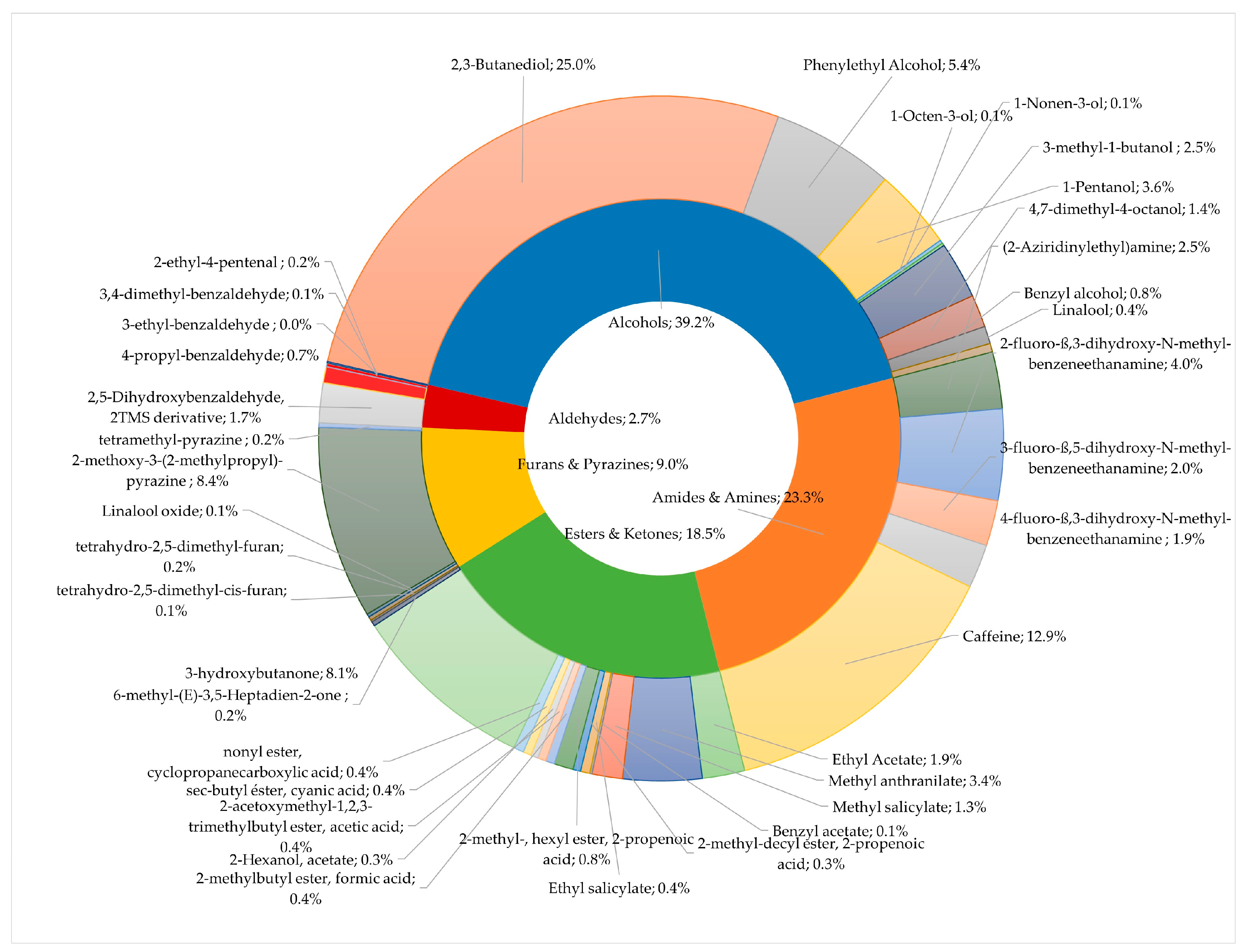
3.2. Coffee Sensory Quality and Volatile Organic Compounds in Green Coffee Beans
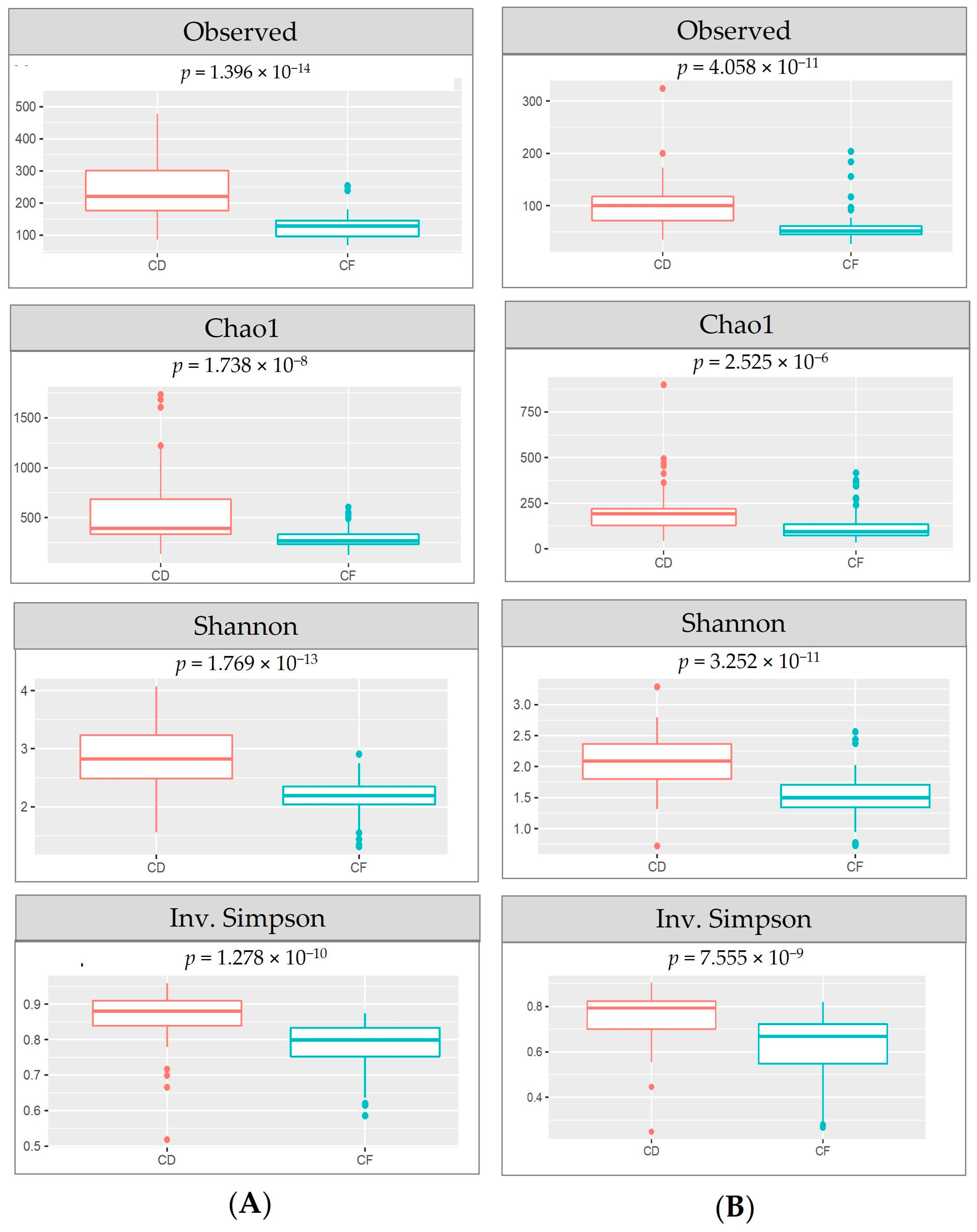
3.3. Microbial Richness and Diversity Using Illumina-Based Amplicon Sequencing
3.4. Taxonomic Composition and Relative Abundance of the Microbial Community in Coffee Fermentation Samples
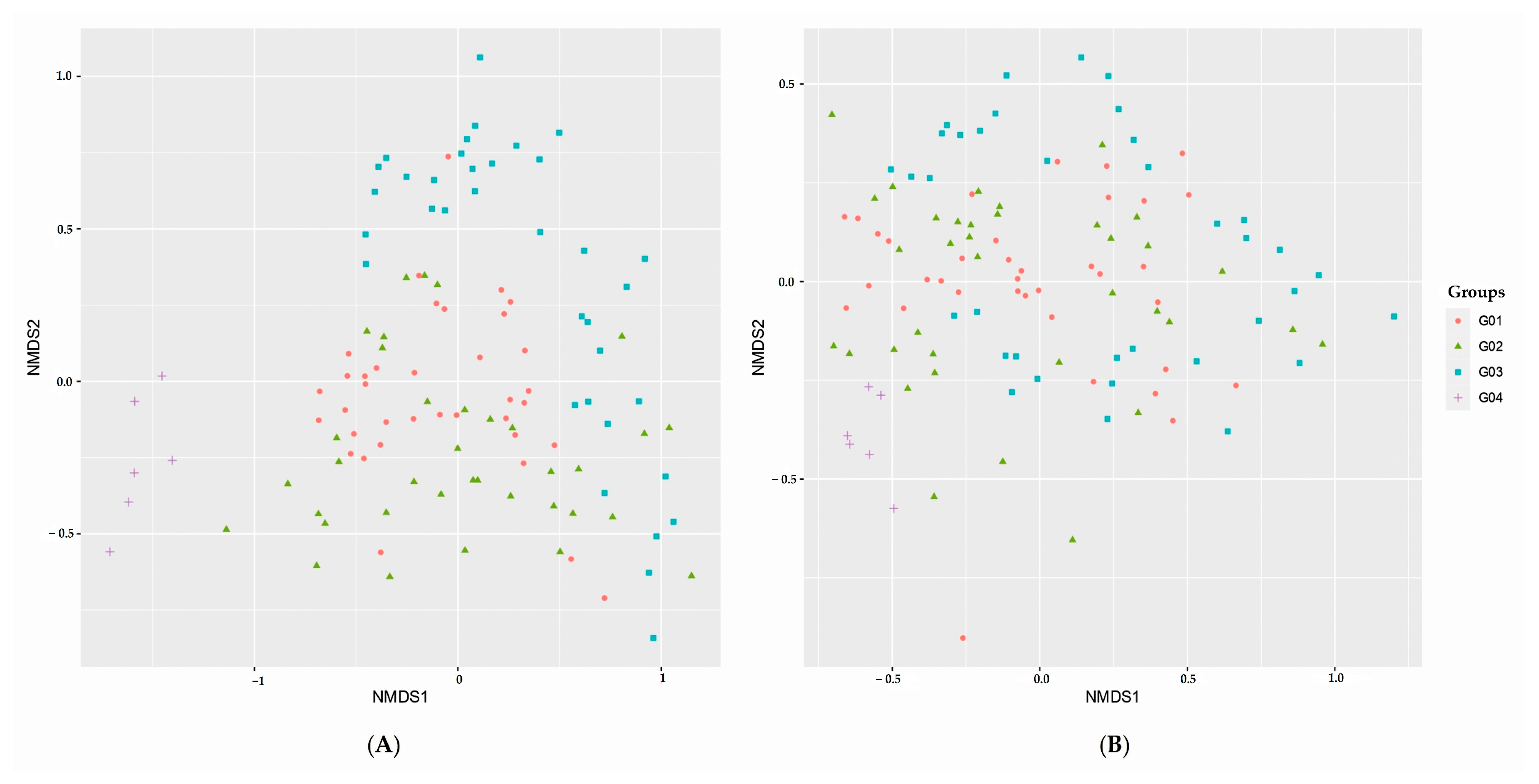
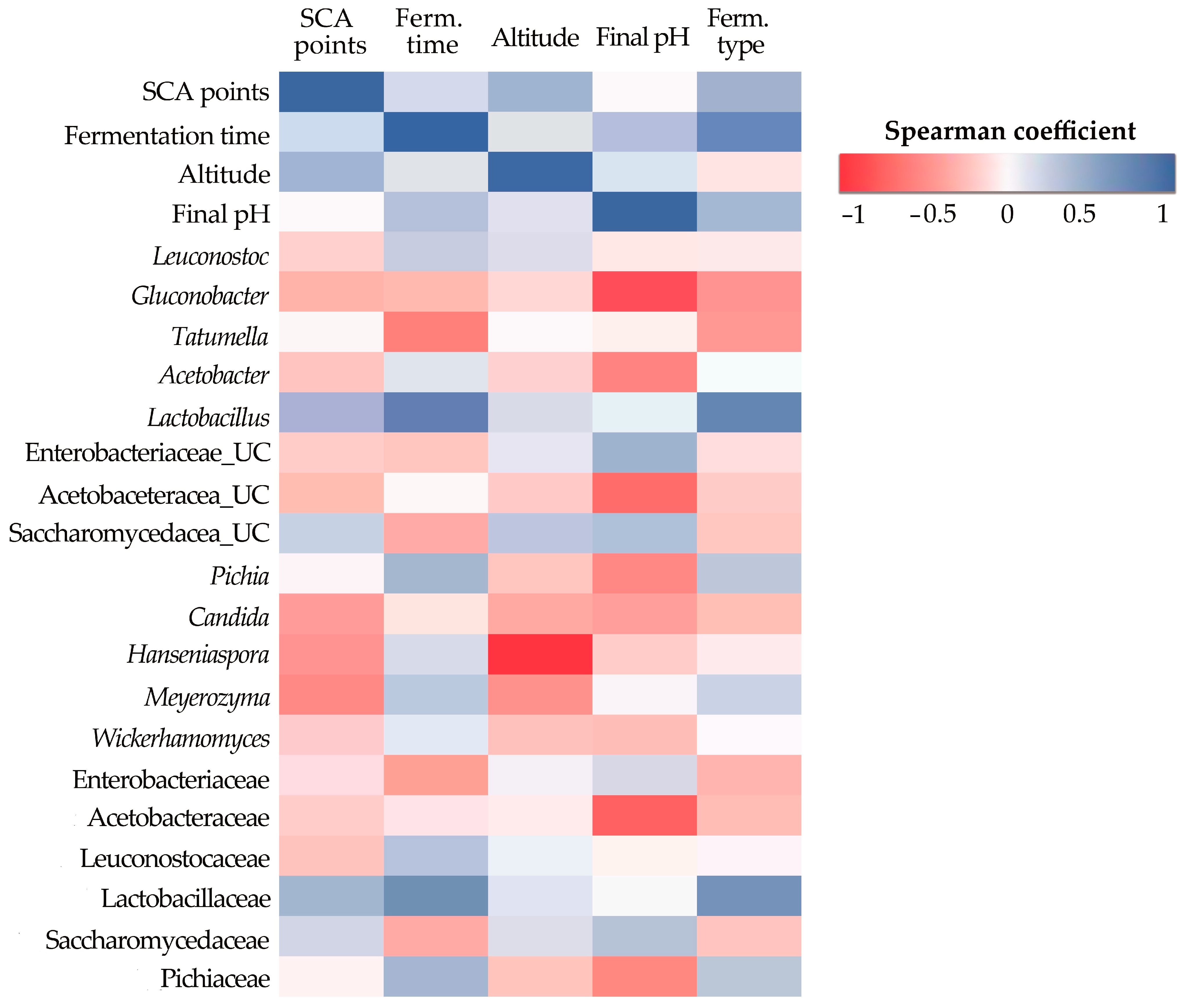
3.5. Microbial Beta Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Coffee Organization (ICO). Available online: https://www.icocoffee.org/documents/cy2022-23/cmr-0223-e.pdf (accessed on 24 March 2023).
- Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.fao.org/markets-and-trade/commodities/coffee/en/ (accessed on 30 January 2023).
- De Oliveira Junqueira, A.C.; de Melo Pereira, G.V.; Coral Medina, J.D.; Alvear, M.C.R.; Rosero, R.; de Carvalho Neto, D.P.; Enríquez, H.G.; Soccol, C.R. First Description of Bacterial and Fungal Communities in Colombian Coffee Beans Fermentation Analysed Using Illumina-Based Amplicon Sequencing. Sci. Rep. 2019, 9, 8794. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Uribe, J.R.; Yusianto; Menon, S.N.; Peñuela, A.; Oliveros, C.; Husson, J.; Brando, C.; Rodriguez, A. Chapter 3-Postharvest Processing—Revealing the Green Bean. In The Craft and Science of Coffee; Folmer, B., Ed.; Academic Press: London, UK, 2017; pp. 51–79. ISBN 978-0-12-803520-7. [Google Scholar]
- Elhalis, H.; Cox, J.; Zhao, J. Coffee Fermentation: Expedition from Traditional to Controlled Process and Perspectives for Industrialization. Appl. Food Res. 2023, 3, 100253. [Google Scholar] [CrossRef]
- Haile, M.; Kang, W.H. Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation. Microorganisms 2019, 7, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peñuela-Martínez, A.E.; Zapata-Zapata, A.D.; Durango-Restrepo, D.L. Performance of different fermentation methods and the effect on coffee quality (Coffea arabica L.). Coffee Sci. 2018, 13, 465–476. [Google Scholar] [CrossRef]
- Lee, L.W.; Cheong, M.W.; Curran, P.; Yu, B.; Liu, S.Q. Coffee Fermentation and Flavor—An Intricate and Delicate Relationship. Food Chem. 2015, 185, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Contreras, G.F.; Cai, Z.; Moccand, C.; Weckx, S.; De Vuyst, L. Influence of Various Processing Parameters on the Microbial Community Dynamics, Metabolomic Profiles, and Cup Quality During Wet Coffee Processing. Front. Microbiol. 2019, 10, 2621. [Google Scholar] [CrossRef]
- Peñuela-Martínez, A.-E.; Tibaduiza-Vianchá, C.-A.; Morcillo, C.-A.; Restrepo-Rivera, M.-V. Enzymatic Mucilage Degradation from Coffea arabica L., for Washed Coffee Production. Biotecnol. Sect. Agropecu. Agroind. 2021, 19, 170–183. [Google Scholar] [CrossRef]
- De Bari, I.; Giuliano, A.; Petrone, M.T.; Stoppiello, G.; Fatta, V.; Giardi, C.; Razza, F.; Novelli, A. From Cardoon Lignocellulosic Biomass to Bio-1,4 Butanediol: An Integrated Biorefinery Model. Processes 2020, 8, 1585. [Google Scholar] [CrossRef]
- Peñuela, A.E.; Romero-Tabarez, M.; Zapata-Zapata, A.D. Functional Diversity of Microbial Communities Associated with Fermentation Processes in Coffee (Coffea arabica L.). Coffee Sci. 2021, 16, e161825. [Google Scholar] [CrossRef]
- Pothakos, V.; De Vuyst, L.; Zhang, S.J.; De Bruyn, F.; Verce, M.; Torres, J.; Callanan, M.; Moccand, C.; Weckx, S. Temporal Shotgun Metagenomics of an Ecuadorian Coffee Fermentation Process Highlights the Predominance of Lactic Acid Bacteria. Curr. Res. Biotechnol. 2020, 2, 1–15. [Google Scholar] [CrossRef]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Torres, J.; Falconi, C.; Moccand, C.; Weckx, S.; De Vuyst, L. Following Coffee Production from Cherries to Cup: Microbiological and Metabolomic Analysis of Wet Processing of Coffea arabica. Appl. Environ. Microbiol. 2019, 85, 6. [Google Scholar] [CrossRef] [Green Version]
- Barooah, M.; Joshi, S.R.; Bahar, B. Editorial: Genomics and Metabolomics of Microbes in Fermented Food. Front. Microbiol. 2022, 13, 892726. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, M.; O’Sullivan, O.; Cotter, P.D.; Sinderen, D.v.; Kenny, J.G. The Application of Metagenomics to Study Microbial Communities and Develop Desirable Traits in Fermented Foods. Foods 2022, 11, 3297. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Ercolini, D. Recent Past, Present, and Future of the Food Microbiome. Annu. Rev. Food Sci. Technol. 2018, 9, 589–608. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Applications of Next Generation Sequencing Technologies as Applied to Food-Related Microbiome Studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feranchuk, S.; Belkova, N.; Potapova, U.; Kuzmin, D.; Belikov, S. Evaluating the Use of Diversity Indices to Distinguish between Microbial Communities with Different Traits. Res. Microbiol. 2018, 169, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Cruz-O’Byrne, R.; Piraneque-Gambasica, N.; Aguirre-Forero, S. Microbial Diversity Associated with Spontaneous Coffee Bean Fermentation Process and Specialty Coffee Production in Northern Colombia. Int. J. Food Microbiol. 2021, 354, 109282. [Google Scholar] [CrossRef]
- De Bruyn, F.; Zhang, S.J.; Pothakos, V.; Torres, J.; Lambot, C.; Moroni, A.V.; Callanan, M.; Sybesma, W.; Weckx, S.; De Vuyst, L. Exploring the Impacts of Postharvest Processing on the Microbiota and Metabolite Profiles during Green Coffee Bean Production. Appl. Environ. Microbiol. 2016, 83, e02398-16. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Dong, H.; Yang, P.; Yang, R.; Lu, J.; Lv, J.; Sheng, J. Culture-Dependent and -Independent Methods to Investigate the Predominant Microorganisms Associated with Wet Processed Coffee. Curr. Microbiol. 2016, 73, 190–195. [Google Scholar] [CrossRef]
- Evangelista, S.R.; Miguel, M.G.d.C.P.; Silva, C.F.; Pinheiro, A.C.M.; Schwan, R.F. Microbiological Diversity Associated with the Spontaneous Wet Method of Coffee Fermentation. Int. J. Food Microbiol. 2015, 210, 102–112. [Google Scholar] [CrossRef]
- De Carvalho Neto, D.P.; Vinícius de Melo, G.; Pereira; César de Carvalho, J.; Soccol, V.T.; Soccol, C.R. High-Throughput rRNA Gene Sequencing Reveals High and Complex Bacterial Diversity Associated with Brazilian Coffee Bean Fermentation. Food Technol. Biotechnol. 2017, 56, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Pregolini, V.B.; de Melo Pereira, G.V.; da Silva Vale, A.; de Carvalho Neto, D.P.; Soccol, C.R. Influence of Environmental Microbiota on the Activity and Metabolism of Starter Cultures Used in Coffee Beans Fermentation. Fermentation 2021, 7, 278. [Google Scholar] [CrossRef]
- De Carvalho Neto, D.P.; De Melo Pereira, G.V.; Tanobe, V.O.A.; Thomaz Soccol, V.G.; da Silva, B.J.; Rodrigues, C.; Soccol, C.R. Yeast Diversity and Physicochemical Characteristics Associated with Coffee Bean Fermentation from the Brazilian Cerrado Mineiro Region. Fermentation 2017, 3, 11. [Google Scholar] [CrossRef]
- Elhalis, H.; Cox, J.; Zhao, J. Ecological Diversity, Evolution and Metabolism of Microbial Communities in the Wet Fermentation of Australian Coffee Beans. Int. J. Food Microbiol. 2020, 321, 108544. [Google Scholar] [CrossRef]
- Da Silva Vale, A.; Balla, G.; Rodrigues, L.R.S.; de Carvalho Neto, D.P.; Soccol, C.R.; de Melo Pereira, G.V. Understanding the Effects of Self-Induced Anaerobic Fermentation on Coffee Beans Quality: Microbiological, Metabolic, and Sensory Studies. Foods 2023, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Vale, A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Sorto, R.D.; Goés-Neto, A.; Kato, R.; Soccol, C.R. Facility-Specific ‘House’ Microbiome Ensures the Maintenance of Functional Microbial Communities into Coffee Beans Fermentation: Implications for Source Tracking. Environ. Microbiol. Rep. 2021, 13, 470–481. [Google Scholar] [CrossRef]
- Plataforma Agroclimática Cafetera—Agroclima. Available online: https://agroclima.cenicafe.org/caracterizacion-agroclimatica (accessed on 24 February 2023).
- Specialty Coffee Association Available (SCA). Available online: https://sca.coffee/research/protocols-best-practices (accessed on 24 February 2023).
- Coffee Quality Institute (CQI). Available online: https://www.coffeeinstitute.org/ (accessed on 24 February 2023).
- Mass Spectral Databases. Available online: https://sciencesolutions.wiley.com/mass-spectral-databases/ (accessed on 15 February 2023).
- DNeasy PowerLyzer PowerSoil Kit Handbook—QIAGEN. Available online: https://www.qiagen.com/at/resources/resourcedetail?id=329362e4-03e6-4ae1-9e4e-bbce41abe4b7&lang=en (accessed on 24 February 2023).
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Põlme, S.; Riit, T.; Liiv, I.; Kõljalg, U.; Kisand, V.; Nilsson, H.; Hildebrand, F.; et al. Shotgun Metagenomes and Multiple Primer Pair-Barcode Combinations of Amplicons Reveal Biases in Metabarcoding Analyses of Fungi. MycoKeys 2015, 10, 1–43. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 2016, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahti, L.; Shetty, S. Microbiome R Package; Bioconductor: Cambridge, UK, 2017. [Google Scholar]
- Chao, A.; Lee, S.-M. Estimating the Number of Classes via Sample Coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Holland, S.M. Non-Metric Multidimensional Scaling (NMDS); R Documentation; Department of Geology, University of Georgia: Athens, Greece, 2008; Available online: https://strata.uga.edu/software/pdf/mdsTutorial.pdf (accessed on 26 January 2023).
- Simmer, M.M.B.; Soares da Silva, M.d.C.; Pereira, L.L.; Moreira, T.R.; Guarçoni, R.C.; Veloso, T.G.R.; da Silva, I.M.R.; Entringer, T.L.; Kasuya, M.C.M.; da Luz, J.M.R.; et al. Edaphoclimatic Conditions and the Soil and Fruit Microbiota Influence on the Chemical and Sensory Quality of the Coffee Beverage. Eur. Food Res. Technol. 2022, 248, 2941–2953. [Google Scholar] [CrossRef]
- Pereira, T.S.; Batista, N.N.; Santos Pimenta, L.P.; Martinez, S.J.; Ribeiro, L.S.; Oliveira Naves, J.A.; Schwan, R.F. Self-Induced Anaerobiosis Coffee Fermentation: Impact on Microbial Communities, Chemical Composition and Sensory Quality of Coffee. Food Microbiol. 2022, 103, 103962. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.J.; Batista, N.N.; Bressani, A.P.P.; Dias, D.R.; Schwan, R.F. Molecular, Chemical, and Sensory Attributes Fingerprinting of Self-Induced Anaerobic Fermented Coffees from Different Altitudes and Processing Methods. Foods 2022, 11, 3945. [Google Scholar] [CrossRef]
- Verce, M.; Schoonejans, J.; Hernandez Aguirre, C.; Molina-Bravo, R.; De Vuyst, L.; Weckx, S. A Combined Metagenomics and Metatranscriptomics Approach to Unravel Costa Rican Cocoa Box Fermentation Processes Reveals Yet Unreported Microbial Species and Functionalities. Front. Microbiol. 2021, 12, 641185. [Google Scholar] [CrossRef] [PubMed]
- Duong, B.; Marraccini, P.; Maeght, J.-L.; Vaast, P.; Lebrun, M.; Duponnois, R. Coffee Microbiota and Its Potential Use in Sustainable Crop Management. A Review. Front. Sustain. Food Syst. 2020, 4, 607935. [Google Scholar] [CrossRef]
- López, C.I.; Bautista, E.; Moreno, E.; Dentan, E. Factors Related to the Formation of “overfermented Coffee Beans” during the Wet Processing Method and Storage of Coffee. In Proceedings of the 13th International Scientific Colloquium on Coffee, Paipa, Colombia, 21–25 August 1989; Association Scientifique Internationale du Café: Paris, France, 1990; pp. 373–384. [Google Scholar]
- Wang, D.; He, M.; Zhang, M.; Yang, H.; Huang, J.; Zhou, R.; Jin, Y.; Wu, C. Food Yeasts: Occurrence, Functions, and Stress Tolerance in the Brewing of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Masoud, W.; Kaltoft, C.H. The Effects of Yeasts Involved in the Fermentation of Coffea arabica in East Africa on Growth and ochratoxin A (OTA) Production by Aspergillus ochraceus. Int. J. Food Microbiol. 2006, 106, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Krajangsang, S.; Seephin, P.; Tantayotai, P.; Mahingsapun, R.; Meeampun, Y.; Panyachanakul, T.; Samosorn, S.; Dolsophon, K.; Jiamjariyatam, R.; Lorliam, W.; et al. New Approach for Screening of Microorganisms from Arabica Coffee Processing for Their Ability to Improve Arabica Coffee Flavor. 3 Biotech 2022, 12, 143. [Google Scholar] [CrossRef]
- Da Silva Vale, A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Rodrigues, C.; Pagnoncelli, M.G.B.; Soccol, C.R. Effect of Co-Inoculation with Pichia Fermentans and Pediococcus Acidilactici on Metabolite Produced During Fermentation and Volatile Composition of Coffee Beans. Fermentation 2019, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. The Crucial Role of Yeasts in the Wet Fermentation of Coffee Beans and Quality. Int. J. Food Microbiol. 2020, 333, 108796. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; de Carvalho Neto, D.P.; Magalhães Júnior, A.I.; Vásquez, Z.S.; Medeiros, A.B.P.; Vandenberghe, L.P.S.; Soccol, C.R. Exploring the Impacts of Postharvest Processing on the Aroma Formation of Coffee Beans—A Review. Food Chem. 2019, 272, 441–452. [Google Scholar] [CrossRef]
- Perry, D.M.; Byrnes, N.K.; Heymann, H.; Hayes, J.E. Rejection of Labrusca-Type Aromas in Wine Differs by Wine Expertise and Geographic Region. Food Qual. Prefer. 2019, 74, 147–154. [Google Scholar] [CrossRef]
- Tsegay, G.; Redi-Abshiro, M.; Chandravanshi, B.S.; Ele, E.; Mohammed, A.M.; Mamo, H. Volatile Profile of Green Coffee Beans from Coffea arabica L. Plants Grown at Different Altitudes in Ethiopia. Bull. Chem. Soc. Ethiop. 2019, 33, 401–413. [Google Scholar] [CrossRef]
- Martinez, S.J.; Bressani, A.P.P.; Dias, D.R.; Simão, J.B.P.; Schwan, R.F. Effect of Bacterial and Yeast Starters on the Formation of Volatile and Organic Acid Compounds in Coffee Beans and Selection of Flavors Markers Precursors During Wet Fermentation. Front. Microbiol. 2019, 10, 1287. [Google Scholar] [CrossRef]
- Batista, N.N.; Ramos, C.L.; Dias, D.R.; Pinheiro, A.C.M.; Schwan, R.F. The Impact of Yeast Starter Cultures on the Microbial Communities and Volatile Compounds in Cocoa Fermentation and the Resulting Sensory Attributes of Chocolate. J. Food Sci. Technol. 2016, 53, 1101–1110. [Google Scholar] [CrossRef] [Green Version]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. Microbiological and Chemical Characteristics of Wet Coffee Fermentation Inoculated With Hansinaspora uvarum and Pichia kudriavzevii and Their Impact on Coffee Sensory Quality. Front. Microbiol. 2021, 12, 713969. [Google Scholar] [CrossRef] [PubMed]
- Bressani, A.P.P.; Martinez, S.J.; Sarmento, A.B.I.; Borém, F.M.; Schwan, R.F. Influence of Yeast Inoculation on the Quality of Fermented Coffee (Coffea arabica Var. Mundo Novo) Processed by Natural and Pulped Natural Processes. Int. J. Food Microbiol. 2021, 343, 109107. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Park, S.-E.; Kim, E.-J.; Seo, S.-H.; Son, H.-S. Investigation of Metabolite Differences in Green Coffee Beans Fermented with Various Microbes. LWT 2022, 172, 114202. [Google Scholar] [CrossRef]
- Ribeiro, L.S.; Evangelista, S.R.; da Cruz Pedrozo Miguel, M.G.; van Mullem, J.; Silva, C.F.; Schwan, R.F. Microbiological and Chemical-Sensory Characteristics of Three Coffee Varieties Processed by Wet Fermentation. Ann. Microbiol. 2018, 68, 705–716. [Google Scholar] [CrossRef]
- Cassimiro, D.M.d.J.; Batista, N.N.; Fonseca, H.C.; Naves, J.A.O.; Dias, D.R.; Schwan, R.F. Coinoculation of Lactic Acid Bacteria and Yeasts Increases the Quality of Wet Fermented Arabica Coffee. Int. J. Food Microbiol. 2022, 369, 109627. [Google Scholar] [CrossRef]
- Gonzalez-Rios, O.; Suarez-Quiroz, M.L.; Boulanger, R.; Barel, M.; Guyot, B.; Guiraud, J.-P.; Schorr-Galindo, S. Impact of “Ecological” Post-Harvest Processing on the Volatile Fraction of Coffee Beans: I. Green Coffee. J. Food Compos. Anal. 2007, 20, 289–296. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample-ID | Municipality | Altitude (m) | Temperature Mean Annual Average. (°C) | Relative Humidity Mean Annual Average. (%) |
|---|---|---|---|---|
| F01 | La Tebaida | 1179 | 22.0 | 76 |
| F02 | Salento | 1642 | 19.3 | 81 |
| F03 | Filandia | 1623 | 19.3 | 80 |
| F04 | Calarcá | 1550 | 20.4 | 79 |
| F05 | Córdoba | 1610 | 19.4 | 80 |
| F06 | Buenavista | 1695 | 18.7 | 81 |
| F07 | Génova | 1593 | 19.8 | 80 |
| F08 | Circasia | 1646 | 19.1 | 81 |
| F09 | Quimbaya | 1356 | 21.0 | 77 |
| F10 | Armenia | 1599 | 19.5 | 80 |
| F11 | Armenia | 1317 | 21.2 | 77 |
| F12 | Génova | 1832 | 18.1 | 83 |
| F13 | Circasia | 1600 | 19.5 | 80 |
| F14 | Filandia | 1558 | 19.9 | 80 |
| F15 | Montenegro | 1288 | 21.4 | 77 |
| F16 | Armenia | 1551 | 19.9 | 80 |
| F17 | Pijao | 1721 | 18.5 | 82 |
| F18 | Salento | 1561 | 19.8 | 80 |
| F19 | Calarcá | 1467 | 21.1 | 78 |
| F20 | Buenavista | 1329 | 21.1 | 77 |
| Fermentation Type | Description | Time (h) | Frequency (%) |
|---|---|---|---|
| Traditional | Fermentation occurs spontaneously until the mucilage is broken down into water-soluble compounds, to be removed from the grains by washing. It usually happens over one night. | 12–16 | 55 |
| Prolonged | After mucilage is degraded, fermentation is terminated after an additional time, arbitrarily defined by the coffee farmer. | 24–72 | 10 |
| Mix of de-pulped coffee batches | Fermentation is carried out with batches of de-pulped coffee from different days of harvest. The amounts of mixed coffee are not controlled. | 48–72 | 15 |
| Underwater | This corresponds to traditional fermentation with the addition of water until the de-pulped coffee mass is completely covered. The relationship between the volume of water (L) and the amount of coffee (kg) is not controlled. | 14–24 | 15 |
| Induced Anaerobic | Coffee fruits or berries are placed in closed containers to avoid contact with air. Generated lixiviates are collected and periodically added to the coffee mass. | 72–96 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peñuela-Martínez, A.E.; Velasquez-Emiliani, A.V.; Angel, C.A. Microbial Diversity Using a Metataxonomic Approach, Associated with Coffee Fermentation Processes in the Department of Quindío, Colombia. Fermentation 2023, 9, 343. https://doi.org/10.3390/fermentation9040343
Peñuela-Martínez AE, Velasquez-Emiliani AV, Angel CA. Microbial Diversity Using a Metataxonomic Approach, Associated with Coffee Fermentation Processes in the Department of Quindío, Colombia. Fermentation. 2023; 9(4):343. https://doi.org/10.3390/fermentation9040343
Chicago/Turabian StylePeñuela-Martínez, Aida Esther, Anyela Vanessa Velasquez-Emiliani, and Carlos A. Angel. 2023. "Microbial Diversity Using a Metataxonomic Approach, Associated with Coffee Fermentation Processes in the Department of Quindío, Colombia" Fermentation 9, no. 4: 343. https://doi.org/10.3390/fermentation9040343
APA StylePeñuela-Martínez, A. E., Velasquez-Emiliani, A. V., & Angel, C. A. (2023). Microbial Diversity Using a Metataxonomic Approach, Associated with Coffee Fermentation Processes in the Department of Quindío, Colombia. Fermentation, 9(4), 343. https://doi.org/10.3390/fermentation9040343