
Dynamics of Microbial Communities in Nitrite-Free and Nutritionally Improved Dry Fermented Sausages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dry Fermented Sausage Manufacturing
2.2. Microbial Counts, pH, aw and Weight Loss
2.3. Monitoring of Starter Culture Implantation
2.4. DNA Purification, qPCR and High Throughput Sequencing
2.5. Bioinformatic Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Characteristics of DFS
3.2. Culture-Dependent Microbial Dynamics
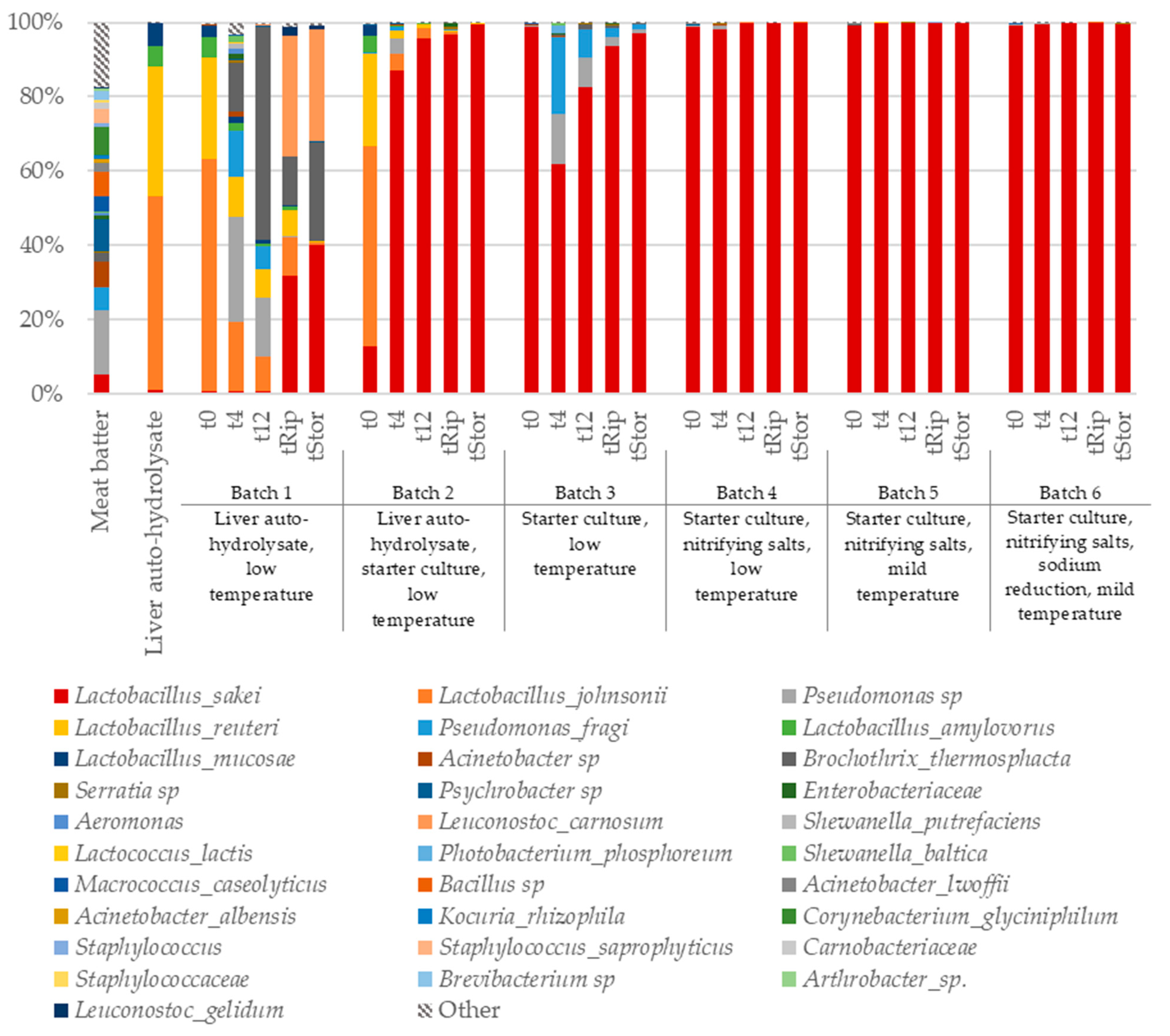
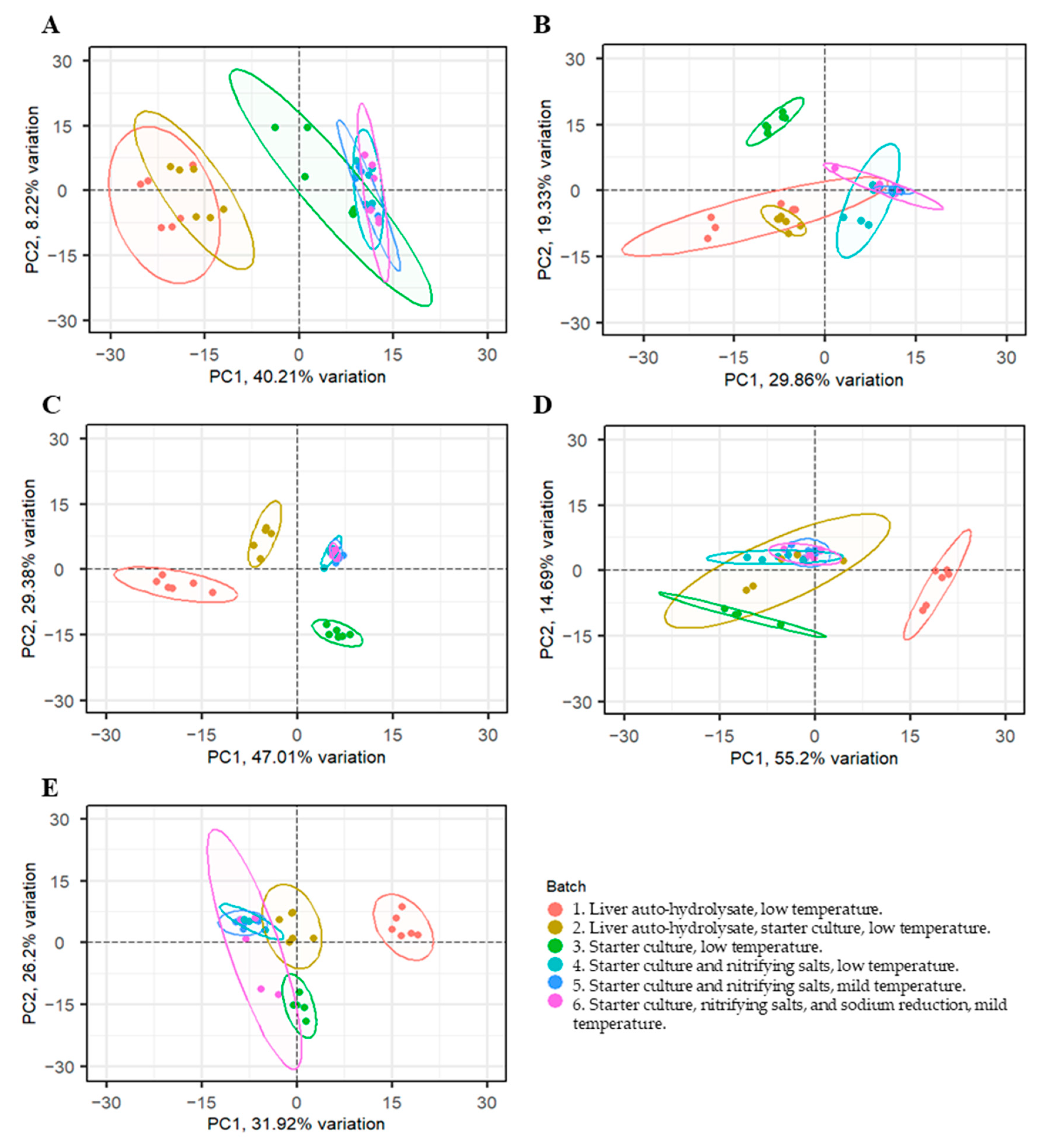
3.3. Diversity and Taxonomic Composition of the Bacterial Communities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garriga, M.; Aymerich, T. The microbiology of fermentation and ripening. In Handbook of Fermented Meat and Poultry; Toldrá, F., Hui, Y.H., Astiasarán, I., Sebranek, J.G., Talon, R., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 107–115. [Google Scholar]
- Austrich-Comas, A.; Jofré, A.; Gou, P.; Bover-Cid, S. Assessing the impact of different technological strategies on the fate of Salmonella in chicken dry-fermented sausages by means of challenge testing and predictive models. Microorganisms 2023, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Rubio, R.; Jofré, A.; Aymerich, T.; Guàrdia, M.D.; Garriga, M. Nutritionally enhanced fermented sausages as a vehicle for potential probiotic lactobacilli delivery. Meat Sci. 2014, 96, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Toldrá, F. Microbial enzymatic activities for improved fermented meats. Trends Food Sci. Technol. 2011, 22, 81–90. [Google Scholar] [CrossRef]
- Martín, B.; Garriga, M.; Hugas, M.; Bover-Cid, S.; Veciana-Nogués, M.T.; Aymerich, T. Molecular, technological and safety characterization of Gram-positive catalase-positive cocci from slightly fermented sausages. Int. J. Food Microbiol. 2006, 107, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Nychas, G.J.E.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Leistner, L.; Gorris, L.G.M. Food preservation by hurdle technology. Trends Food Sci. Technol. 1995, 6, 41–46. [Google Scholar] [CrossRef]
- De Araújo, P.D.; Araújo, W.M.C.; Patarata, L.; Fraqueza, M.J. Understanding the main factors that influence consumer quality perception and attitude towards meat and processed meat products. Meat Sci. 2022, 193, 108952. [Google Scholar] [CrossRef]
- Marco, A.; Navarro, J.L.; Flores, M. The influence of nitrite and nitrate on microbial, chemical and sensory parameters of slow dry fermented sausage. Meat Sci. 2006, 73, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Tabanelli, G.; Barbieri, F.; Soglia, F.; Magnani, R.; Gardini, G.; Petracci, M.; Gardini, F.; Montanari, C. Safety and technological issues of dry fermented sausages produced without nitrate and nitrite. Food Res. Int. 2022, 160, 111685. [Google Scholar] [CrossRef]
- De Mey, E. N-Nitrosamines in Dry Fermented Sausages: Occurrence and Formation of n-Nitrosopiperidine; KU Leuven: Leuven, Belgium, 2014; ISBN 978-90-8826-353-8. [Google Scholar]
- Abril, B.; Sanchez-Torres, E.A.; Bou, R.; Benedito, J.; Garcia-Perez, J.V. Influence of pork liver drying on ferrochelatase activity for zinc protoporphyrin formation. LWT 2022, 171, 114128. [Google Scholar] [CrossRef]
- Wakamatsu, J.I.; Murakami, N.; Nishimura, T. A comparative study of zinc protoporphyrin ix-forming properties of animal by-products as sources for improving the color of meat products. Anim. Sci. J. 2015, 86, 547–552. [Google Scholar] [CrossRef] [PubMed]
- EFSA NDA Panel (EFSA Panel on Nutrition Novel Foods and Food Allergens); Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Knutsen, H.K.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; et al. Dietary Reference Values for Sodium. EFSA J. 2019, 17, e05778. [Google Scholar] [CrossRef] [PubMed]
- Stollewerk, K.; Jofré, A.; Comaposada, J.; Arnau, J.; Garriga, M. The impact of fast drying (QDS Process®) and high pressure on food safety of NaCl-free processed dry fermented sausages. Innov. Food Sci. Emerg. Technol. 2012, 16, 89–95. [Google Scholar] [CrossRef]
- Teixeira, A.; Domínguez, R.; Ferreira, I.; Pereira, E.; Estevinho, L.; Rodrigues, S.; Lorenzo, J.M. Effect of NaCl replacement by other salts on the quality of bísaro pork sausages (Pgi Chouriça de Vinhais). Foods 2021, 10, 961. [Google Scholar] [CrossRef]
- Majou, D.; Christieans, S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Sci. 2018, 145, 273–284. [Google Scholar] [CrossRef]
- Franciosa, I.; Alessandria, V.; Dolci, P.; Rantsiou, K.; Cocolin, L. Sausage fermentation and starter cultures in the era of molecular biology methods. Int. J. Food Microbiol. 2018, 279, 26–32. [Google Scholar] [CrossRef]
- Belleggia, L.; Ferrocino, I.; Reale, A.; Haouet, M.N.; Corvaglia, M.R.; Milanović, V.; Boscaino, F.; Di Renzo, T.; Di Bella, S.; Borghi, M.; et al. Unravelling microbial populations and volatile organic compounds of artisan fermented liver sausages manufactured in central Italy. Food Res. Int. 2022, 154, 111019. [Google Scholar] [CrossRef]
- Van Reckem, E.; Claeys, E.; Charmpi, C.; Sosa Fajardo, A.; Van der Veken, D.; Maes, D.; Weckx, S.; De Vuyst, L.; Leroy, F. High-throughput amplicon sequencing to assess the impact of processing factors on the development of microbial communities during spontaneous meat fermentation. Int. J. Food Microbiol. 2021, 354, 109322. [Google Scholar] [CrossRef]
- Bou, R.; Llauger, M.; Albano, M.; Tenorio, E. Optimitzation of ZnPP-forming conditions in porcine livers using response surface methodology (RSM). In Proceedings of the 67th International Congress of Meat Science and Technology, Kraków, Poland, 23–27 August 2021. [Google Scholar]
- Hugas, M.; Garriga, M.; Aymerich, M.T.; Monfort, J.M. Inhibition of Listeria in dry fermented sausages by the bacteriocinogenic Lactobacillus Sake CTC494. J. Appl. Bacteriol. 1995, 79, 322–330. [Google Scholar] [CrossRef]
- ANSES. Fiche de Description de Danger Biologique Transmissible Par Les Aliments: Clostridium Botulinum, Clostridium Neurotoxinogènes; ANSES: Maisons-Alfort, France, 2019. [Google Scholar]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards). Opinion of the scientific panel on biological hazards on a request from the commission related to Clostridium spp. in foodstuffs. EFSA J. 2005, 3, 1–65. [Google Scholar] [CrossRef]
- Serra-Castelló, C.; Bover-Cid, S.; Garriga, M.; Hansen, T.B.; Gunvig, A.; Jofré, A. Risk management tool to define a corrective storage to enhance Salmonella inactivation in dry fermented sausages. Int. J. Food Microbiol. 2021, 346, 109160. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R. A Language and Environment for Statistical Computing 2019; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://r-project.org/ (accessed on 30 March 2023).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Gloor, G.B.; Reid, G. Compositional analysis: A valid approach to analyze microbiome high-throughput sequencing data. Can. J. Microbiol. 2016, 62, 692–703. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [PubMed]
- Palarea-Albaladejo, J.; Martín-Fernández, J.A. ZCompositions—R Package for multivariate imputation of left-censored data under a compositional approach. Chemom. Intell. Lab. Syst. 2015, 143, 85–96. [Google Scholar] [CrossRef]
- Martín-Fernández, J.-A.; Hron, K.; Templ, M.; Filzmoser, P.; Palarea-Albaladejo, J. Bayesian-multiplicative treatment of count zeros in compositional data sets. Stat. Model. 2014, 15, 134–158. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Fernandes, A.D.; Macklaim, J.M.; Linn, T.G.; Reid, G.; Gloor, G.B. ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-Seq. PLoS ONE 2013, 8, e67019. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; Mcmurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef]
- Aymerich, T.; Martín, B.; Garriga, M.; Hugas, M. Microbial quality and direct PCR identification of lactic acid bacteria and nonpathogenic staphylococci from artisanal low-acid sausages. Appl. Environ. Microbiol. 2003, 69, 4583–4594. [Google Scholar] [CrossRef]
- Ordöñez, J.A.; de la Hoz, L. Mediterranean products. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 333–347. [Google Scholar]
- Peck, M.W.; Stringer, S.C.; Carter, A.T. Clostridium botulinum in the post-genomic era. Food Microbiol. 2011, 28, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Zagorec, M.; Champomier-Vergès, M.C. Lactobacillus sakei: A starter for sausage fermentation, a protective culture for meat products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Ravyts, F.; Steen, L.; Goemaere, O.; Paelinck, H.; De Vuyst, L.; Leroy, F. The application of staphylococci with flavour-generating potential is affected by acidification in fermented dry sausages. Food Microbiol. 2010, 27, 945–954. [Google Scholar] [CrossRef]
- Barbieri, F.; Tabanelli, G.; Montanari, C.; Dall’osso, N.; Šimat, V.; Možina, S.S.; Baños, A.; Özogul, F.; Bassi, D.; Fontana, C.; et al. Mediterranean spontaneously fermented sausages: Spotlight on microbiological and quality features to exploit their bacterial biodiversity. Foods 2021, 10, 2691. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.J.E. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef]
- Ferrer-Bustins, N.; Martín, B.; Bover-Cid, S.; Jofré, A. Caracterización de La Diversidad Microbiana En Productos Cárnicos Porcinos y Superficies de Instalaciones Con Métodos Dependientes e Independientes de Cultivo. In Proceedings of the XXVIII Congreso Sociedad Española de Microbiología, Online, 28 June–2 July 2021; p. 398. [Google Scholar]
- Ercolini, D.; Casaburi, A.; Nasi, A.; Ferrocino, I.; Di Monaco, R.; Ferranti, P.; Mauriello, G.; Villani, F. Different molecular types of Pseudomonas fragi have the same overall behaviour as meat spoilers. Int. J. Food Microbiol. 2010, 142, 120–131. [Google Scholar] [CrossRef]
- Danilo, E.; Federica, R.; Elena, T.; Paolo, M.; Francesco, V. Changes in the spoilage-related microbiota of beef during refrigerated storage under different packaging conditions. Appl. Environ. Microbiol. 2006, 72, 4663–4671. [Google Scholar] [CrossRef]
- Cardinali, F.; Milanović, V.; Osimani, A.; Aquilanti, L.; Taccari, M.; Garofalo, C.; Polverigiani, S.; Clementi, F.; Franciosi, E.; Tuohy, K.; et al. Microbial dynamics of model Fabriano-like fermented sausages as affected by starter cultures, nitrates and nitrites. Int. J. Food Microbiol. 2018, 278, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Työppönen, S.; Petäjä, E.; Mattila-Sandholm, T. Bioprotectives and probiotics for dry sausages. Int. J. Food Microbiol. 2003, 83, 233–244. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; de Cesare, A.; Hilbert, F.; et al. Statement on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 10: Suitability of taxonomic units notified to EFSA until March 2019. EFSA J. 2019, 17, 1–79. [Google Scholar] [CrossRef]
- Luong, N.D.M.; Coroller, L.; Zagorec, M.; Membré, J.M.; Guillou, S. Spoilage of chilled fresh meat products during storage: A quantitative analysis of literature data. Microorganisms 2020, 8, 1198. [Google Scholar] [CrossRef] [PubMed]
- Casas, I.A.; Dobrogosz, W.J. Validation of the probiotic concept: Lactobacillus reuteri confers broad-spectrum protection against disease in humans and animals. Microb. Ecol. Health Dis. 2000, 12, 247–285. [Google Scholar] [CrossRef]
- Raimondi, S.; Nappi, M.R.; Sirangelo, T.M.; Leonardi, A.; Amaretti, A.; Ulrici, A.; Magnani, R.; Montanari, C.; Tabanelli, G.; Gardini, F.; et al. Bacterial community of industrial raw sausage packaged in modified atmosphere throughout the shelf life. Int. J. Food Microbiol. 2018, 280, 78–86. [Google Scholar] [CrossRef]
- Muthukumarasamy, P.; Holley, R.A. Microbiological and sensory quality of dry fermented sausages containing alginate-microencapsulated Lactobacillus reuteri. Int. J. Food Microbiol. 2006, 111, 164–169. [Google Scholar] [CrossRef]
- Wójciak, K.M.; Dolatowski, Z.J.; Kołożyn-Krajewska, D. Use of acid whey and probiotic strains to improve microbiological quality and sensory acceptance of organic fermented sausage. J. Food Process Preserv. 2015, 39, 539–547. [Google Scholar] [CrossRef]
- Cocconcelli, P.S. Starter cultures: Bacteria. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; Blackwell Publishers: Hoboken, NJ, USA, 2007; pp. 137–145. [Google Scholar]
- Hernández-Macias, S.; Ferrer-Bustins, N.; Comas-Basté, O.; Jofré, A.; Latorre-Moratalla, M.; Bover-Cid, S.; Vidal-Carou, M.D.C. Revalorization of cava lees to improve the safety of fermented sausages. Foods 2021, 10, 1916. [Google Scholar] [CrossRef]
- Rocchetti, G.; Falasconi, I.; Dallolio, G.; Lorenzo, J.M.; Lucini, L.; Rebecchi, A. Impact of hurdle technologies and low temperatures during ripening on the production of nitrate-free pork salami: A microbiological and metabolomic comparison. LWT 2021, 141, 110939. [Google Scholar] [CrossRef]
- Charmpi, C.; Van der Veken, D.; Van Reckem, E.; De Vuyst, L.; Leroy, F. Raw meat quality and salt levels affect the bacterial species diversity and community dynamics during the fermentation of pork mince. Food Microbiol. 2020, 89, 103434. [Google Scholar] [CrossRef] [PubMed]
- Palman, Y.; De Leo, R.; Pulvirenti, A.; Green, S.J.; Hayouka, Z. Antimicrobial peptide cocktail activity in minced turkey meat. Food Microbiol. 2020, 92, 103580. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Specific Additives/Ingredients (g/kg) | Process Conditions 3 | ||||||
|---|---|---|---|---|---|---|---|
| Batch | KNO3/NaNO2 | Liver Auto-Hydrolysate/GDL | NaCl/ KCl | Starter Culture 2 | Days | Temperature (°C) | Relative Humidity (%) |
| 1 | na | 300/3 | 20/0 | na | 20 2 18 | 3.4 ± 0.3 7.8 ± 2.1 12.5 ± 0.4 | 87.7 ± 8.6 90.0 ± 8.3 76.0 ± 8.6 |
| 2 | na | 300/3 | 20/0 | + | |||
| 3 | na | na 1 | 20/0 | + | |||
| 4 | 0.10/ 0.15 | na 1 | 20/0 | + | |||
| 5 | 0.10/ 0.15 | na 1 | 20/0 | + | 27 | 12.4 ± 0.6 | 86.2 ± 7.6 |
| 6 | 0.10/ 0.15 | na 1 | 10/ 12.76 | + | |||
| Time | Batch 1 | Weight Loss | aw | pH | LAB | GCC+ |
|---|---|---|---|---|---|---|
| t0 | 1 | NA | 0.983 ± 0.001 AB | 5.96 ± 0.04 A | 5.01 ± 0.19 B | 4.13 ± 0.19 AB |
| 2 | NA | 0.982 ± 0.000 B | 5.93 ± 0.03 A | 6.19 ± 0.00 A | 4.38 ± 0.03 A | |
| 3 | NA | 0.985 ± 0.001 AB | 5.87 ± 0.11 A | 6.26 ± 0.13 A | 4.03 ± 0.01 AB | |
| 4 | NA | 0.985 ± 0.001 AB | 5.83 ± 0.12 A | 6.08 ± 0.04 A | 3.76 ± 0.09 B | |
| 5 | NA | 0.985 ± 0.001 AB | 5.84 ± 0.08 A | 6.11 ± 0.04 A | 4.04 ± 0.33 AB | |
| 6 | NA | 0.985 ± 0.000 A | 5.90 ± 0.05 A | 6.06 ± 0.02 A | 4.03 ± 0.15 AB | |
| t4 | 1 | 2.49 ± 0.85 C | 0.982 ± 0.000 B | 5.94 ± 0.11 A | 5.17 ± 0.03 C | 3.84 ± 0.03 A |
| 2 | 2.70 ± 0.92 C | 0.982 ± 0.000 B | 5.95 ± 0.13 A | 8.11 ± 0.16 B | 4.13 ± 0.15 A | |
| 3 | 3.68 ± 0.58 C | 0.985 ± 0.001 A | 5.90 ± 0.12 A | 8.24 ± 0.17 B | 3.97 ± 0.16 A | |
| 4 | 12.45 ± 1.23 A | 0.983 ± 0.001 B | 5.90 ± 0.13 A | 8.09 ± 0.28 B | 3.70 ± 0.05 A | |
| 5 | 6.37 ± 0.87 B | 0.984 ± 0.000 A | 5.11 ± 0.08 B | 8.90 ± 0.11 A | 3.40 ± 0.00 A | |
| 6 | 6.00 ± 0.71 B | 0.984 ± 0.000 A | 5.09 ± 0.09 B | 8.85 ± 0.03 A | 3.40 ± 0.00 A | |
| t7 | 1 | 6.87 ± 1.03 C | 0.980 ± 0.000 B | 5.95 ± 0.08 A | 5.27 ± 0.07 C | 4.14 ± 0.27 A |
| 2 | 8.05 ± 2.15 C | 0.980 ± 0.001 B | 5.74 ± 0.04 B | 8.43 ± 0.16 B | 4.38 ± 0.09 A | |
| 3 | 9.74 ± 0.65 BC | 0.983 ± 0.001 A | 5.58 ± 0.02 C | 8.47 ± 0.06 B | 4.07 ± 0.26 A | |
| 4 | 12.16 ± 1.27 AB | 0.983 ± 0.000 AB | 5.66 ± 0.14 BC | 8.40 ± 0.02 B | 3.95 ± 0.30 A | |
| 5 | 13.21 ± 2.59 AB | 0.983 ± 0.002 AB | 4.81 ± 0.03 D | 8.92 ± 0.04 A | 3.52 ± 0.11 B | |
| 6 | 13.46 ± 0.73 A | 0.982 ± 0.001 AB | 4.83 ± 0.02 D | 8.97 ± 0.04 A | 3.51 ± 0.12 B | |
| t12 | 1 | 23.24 ± 2.06 B | 0.976 ± 0.000 A | 5.94 ± 0.09 A | 5.48 ± 0.25 D | 3.77 ± 0.29 A |
| 2 | 26.85 ± 3.47 B | 0.976 ± 0.001 A | 5.30 ± 0.01 B | 8.77 ± 0.04 ABC | 3.55 ± 0.01 A | |
| 3 | 30.85 ± 1.86 B | 0.979 ± 0.000 A | 5.27 ± 0.02 B | 8.54 ± 0.04 C | 3.58 ± 0.21 A | |
| 4 | 24.17 ± 1.70 B | 0.979 ± 0.000 A | 5.31 ± 0.06 B | 8.63 ± 0.09 BC | 3.40 ± 0.05 A | |
| 5 | 44.45 ± 3.82 A | 0.971 ± 0.003 B | 4.85 ± 0.04 C | 9.00 ± 0.16 A | 2.76 ± 0.02 B | |
| 6 | 46.36 ± 3.72 A | 0.970 ± 0.003 B | 4.90 ± 0.06 C | 8.88 ± 0.05 AB | 2.75 ± 0.37 B | |
| t20 | 1 | 37.61 ± 2.36 CD | 0.968 ± 0.004 A | 5.85 ± 0.02 A | ND | ND |
| 2 | 41.38 ± 3.44 CD | 0.967 ± 0.001 A | 5.11 ± 0.04 A | ND | ND | |
| 3 | 45.20 ± 2.10 B | 0.967 ± 0.000 A | 5.09 ± 0.02 A | ND | ND | |
| 4 | 37.01 ± 2.28 D | 0.970 ± 0.004 A | 5.13 ± 0.01 A | ND | ND | |
| 5 | 59.19 ± 2.22 A | 0.939 ± 0.017 AB | 5.30 ± 0.32 A | 8.62 ± 0.10 | 2.73 ± 0.45 | |
| 6 | 59.86 ± 2.13 A | 0.928 ± 0.007 B | 5.59 ± 0.37 A | 8.61 ± 0.07 | 2.53 ± 0.24 | |
| tRip | 1 | 61.41 ± 1.81 B | 0.866 ± 0.016 A | 5.82 ± 0.00 A | 8.25 ± 0.11 B | 1.98 ± 0.73 A |
| 2 | 61.52 ± 1.37 B | 0.846 ± 0.009 A | 5.20 ± 0.05 C | 8.34 ± 0.00 AB | 1.27 ± 0.01 A | |
| 3 | 64.75 ± 0.90 A | 0.840 ± 0.031 A | 5.20 ± 0.02 C | 8.18 ± 0.00 AB | 1.40 ± 0.12 A | |
| 4 | 63.54 ± 1.25 A | 0.857 ± 0.027 A | 5.19 ± 0.01 C | 8.17 ± 0.00 B | 1.90 ± 0.08 A | |
| 5 | 63.95 ± 1.47 A | 0.865 ± 0.012 A | 5.49 ± 0.17 B | 8.32 ± 0.03 AB | 1.90 ± 0.44 A | |
| 6 | 64.24 ± 1.32 A | 0.858 ± 0.013 A | 5.54 ± 0.08 B | 8.53 ± 0.00 A | 2.88 ± 0.92 A | |
| tStor | 1 | ND | ND | 5.83 ± 0.00 A | 8.02 ± 0.06 B | 2.83 ± 0.16 A |
| 2 | ND | ND | 5.35 ± 0.01 C | 8.21 ± 0.00 AB | 2.31 ± 0.23 AB | |
| 3 | ND | ND | 5.23 ± 0.01 C | 8.22 ± 0.12 AB | 2.24 ± 0.11 AB | |
| 4 | ND | ND | 5.23 ± 0.08 C | 8.20 ± 0.08 AB | 1.97 ± 0.01 B | |
| 5 | ND | ND | 5.32 ± 0.04 C | 8.29 ± 0.00 A | 2.25 ± 0.05 AB | |
| 6 | ND | ND | 5.52 ± 0.12 B | 8.30 ± 0.03 A | 2.68 ± 0.24 AB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer-Bustins, N.; Martín, B.; Llauger, M.; Bou, R.; Bover-Cid, S.; Jofré, A. Dynamics of Microbial Communities in Nitrite-Free and Nutritionally Improved Dry Fermented Sausages. Fermentation 2023, 9, 403. https://doi.org/10.3390/fermentation9040403
Ferrer-Bustins N, Martín B, Llauger M, Bou R, Bover-Cid S, Jofré A. Dynamics of Microbial Communities in Nitrite-Free and Nutritionally Improved Dry Fermented Sausages. Fermentation. 2023; 9(4):403. https://doi.org/10.3390/fermentation9040403
Chicago/Turabian StyleFerrer-Bustins, Núria, Belén Martín, Mar Llauger, Ricard Bou, Sara Bover-Cid, and Anna Jofré. 2023. "Dynamics of Microbial Communities in Nitrite-Free and Nutritionally Improved Dry Fermented Sausages" Fermentation 9, no. 4: 403. https://doi.org/10.3390/fermentation9040403
APA StyleFerrer-Bustins, N., Martín, B., Llauger, M., Bou, R., Bover-Cid, S., & Jofré, A. (2023). Dynamics of Microbial Communities in Nitrite-Free and Nutritionally Improved Dry Fermented Sausages. Fermentation, 9(4), 403. https://doi.org/10.3390/fermentation9040403