
Current Status, Challenges, and Prospects for the Biological Production of Vanillin


 and
and
Abstract
:1. Introduction
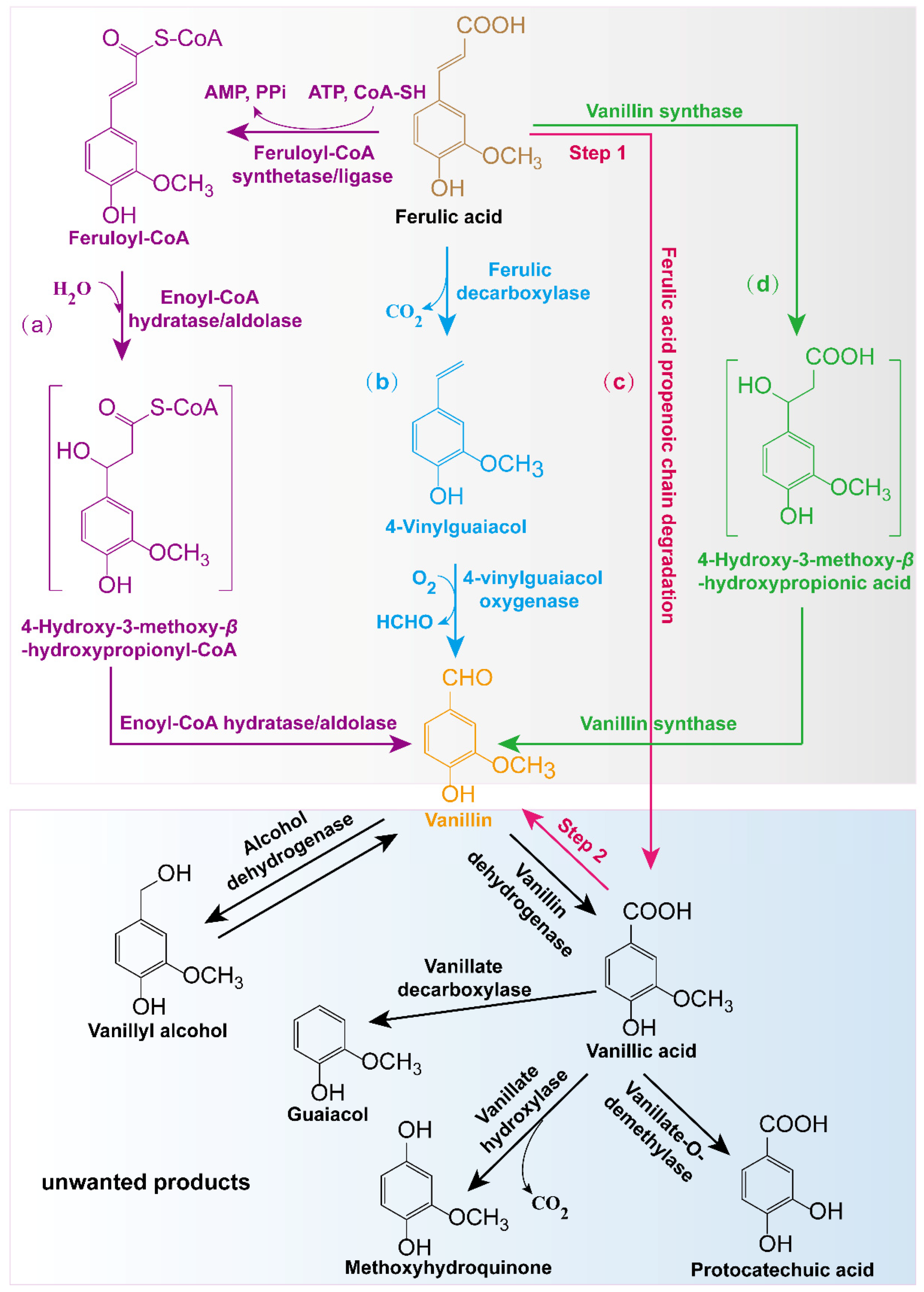
2. Biological Production of Vanillin from Ferulic Acid
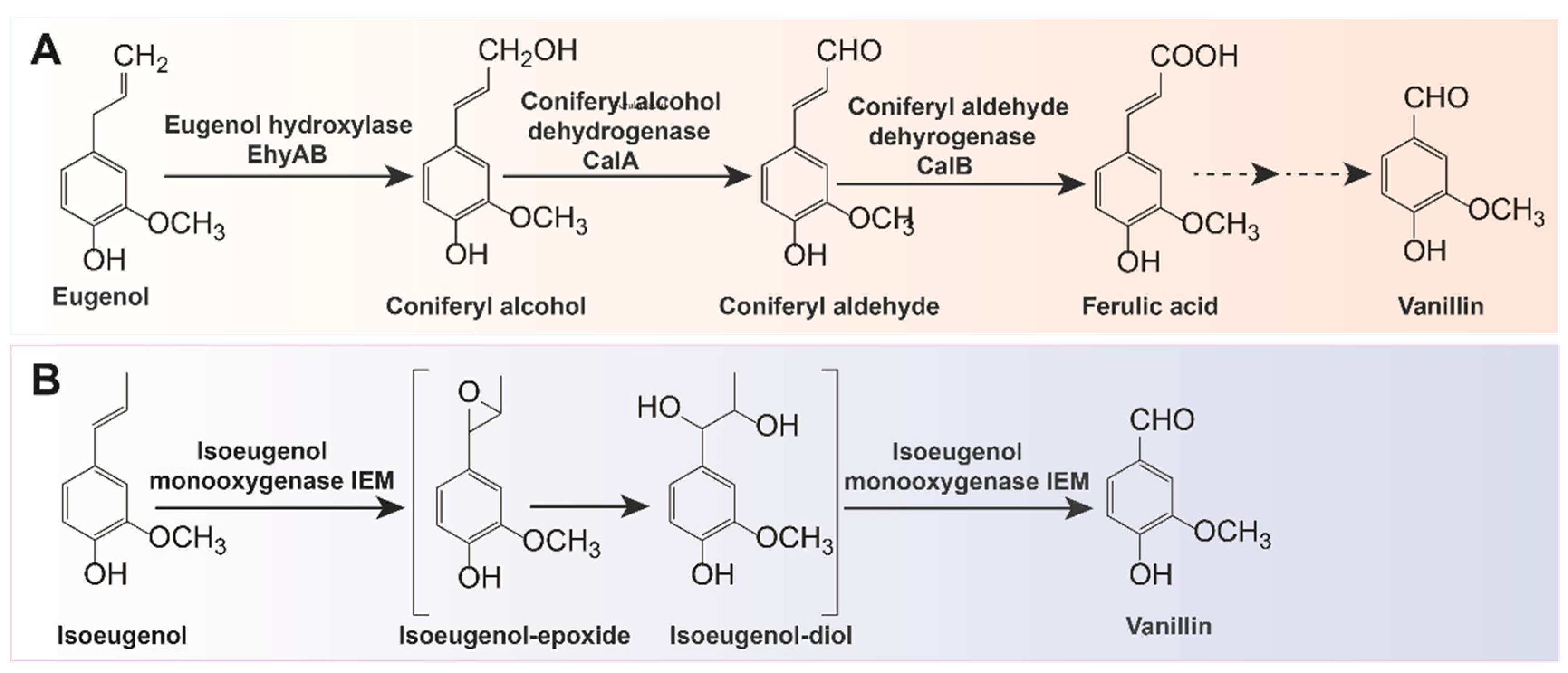
2.1. Biotransformation of Ferulic Acid into Vanillin Using Native Microbial Strains
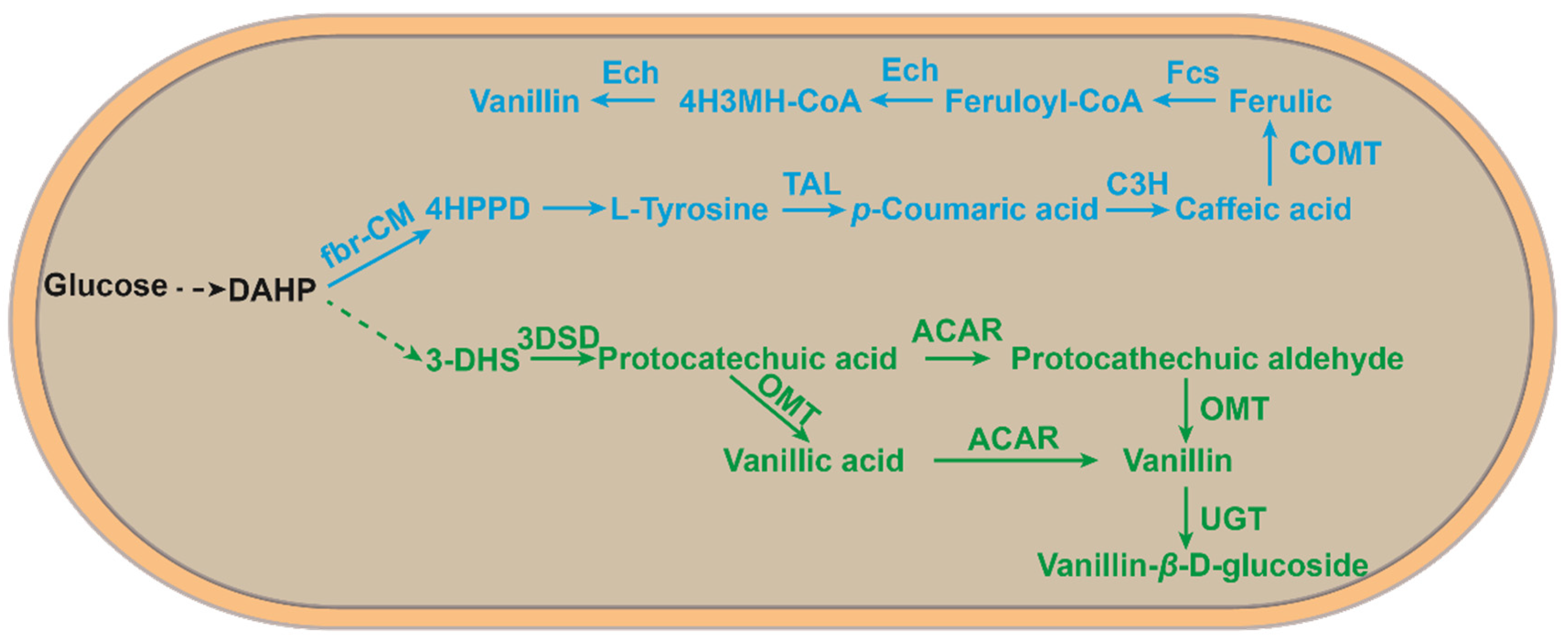
2.2. Biotransformation of Ferulic Acid into Vanillin Using Engineered Microbes
2.3. Synthesis of Vanillin from Ferulic Acid Using Plants
3. Vanillin Bioproduction from Lignin
4. Vanillin Production Using Other Substrates
5. Process Optimization
6. Downstream Engineering: Vanillin Recovery
7. Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banerjee, G.; Chattopadhyay, P. Vanillin biotechnology: The perspectives and future. J. Sci. Food Agric. 2019, 99, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Priefert, H.; Rabenhorst, J.; Steinbüchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef]
- Gallage, N.J.; Møller, B.L. Vanillin-bioconversion and bioengineering of the most popular plant flavor and its de novo biosynthesis in the vanilla orchid. Mol. Plant 2015, 8, 40–57. [Google Scholar] [CrossRef]
- Martău, G.A.; Călinoiu, L.F.; Vodnar, D.C. Bio-vanillin: Towards a sustainable industrial production. Trends Food Sci. Technol. 2021, 109, 579–592. [Google Scholar] [CrossRef]
- Walton, N.J.; Mayer, M.J.; Arjan, N. Molecules of interest vanillin. Phytochesmistry 2003, 63, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Fache, M.; Boutevin, B.; Caillol, S. Vanillin production from lignin and its use as a renewable chemical. ACS Sustain. Chem. Eng. 2016, 4, 35–46. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Cheng, Y.; Sun, H.; Bai, S.; Li, C. Identifying environmental hotspots and improvement strategies of vanillin production with life cycle assessment. Sci. Total Environ. 2021, 769, 144771. [Google Scholar] [CrossRef]
- Ciriminna, R.; Fidalgo, A.; Meneguzzo, F.; Parrino, F.; Ilharco, L.M.; Pagliaro, M. Vanillin: The case for greener production driven by sustainability megatrend. ChemistryOpen 2019, 8, 660–667. [Google Scholar] [CrossRef]
- Galadima, A.I.; Salleh, M.M.; Hussin, H.; Chong, C.S.; Yahya, A.; Mohamad, S.E.; Abd-Aziz, S.; Yusof, N.N.M.; Naser, M.A.; Al-Junid, A.F.M. Biovanillin: Production concepts and prevention of side product formation. Biomass Convers. Bior. 2020, 10, 589–609. [Google Scholar] [CrossRef]
- Furuya, T.; Kuroiwa, M.; Kino, K. Biotechnological production of vanillin using immobilized enzymes. J. Biotechnol. 2017, 243, 25–28. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Delattre, M.; Haon, M.; Thibault, J.F.; Ceccaldi, B.C.; Brunerie, P.; Asther, M. A two-step bioconversion process for vanillin production from ferulic acid combining Aspergillus niger and Pycnoporus cinnabarinus. J. Biotechnol. 1996, 50, 107–113. [Google Scholar] [CrossRef]
- Gallage, N.J.; Hansen, E.H.; Kannangara, R.; Olsen, C.E.; Motawia, M.S.; Jørgensen, K.; Holme, I.; Hebelstrup, K.; Grisoni, M.; Møller, B.L. Vanillin formation from ferulic acid in Vanilla planifolia is catalysed by a single enzyme. Nat. Commun. 2014, 5, 4037. [Google Scholar] [CrossRef] [PubMed]
- Overhage, J.; Priefert, H.; Steinbüchel, A. Biochemical and genetic analyses of ferulic acid catabolism in Pseudomonas sp. strain HR199. Appl. Environ. Microbiol. 1999, 65, 4837–4847. [Google Scholar] [CrossRef]
- Narbad, A.; Gasson, M.J. Metabolism of ferulic acid via vanillin using a novel CoA-dependent pathway in a newly-isolated strain of Pseudomonas fluorescens. Microbiology 1998, 144, 1397–1405. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Haon, M.; Delattre, M.; Thibault, J.F.; Ceccaldi, B.C.; Asther, M. An attempt to channel the transformation of vanillic acid into vanillin by controlling methoxyhydroquinone formation in Pycnoporus cinnabarinus with cellobiose. Appl. Microbiol. Biotechnol. 1997, 47, 393–397. [Google Scholar] [CrossRef]
- Ding, W.; Si, M.; Zhang, W.; Zhang, Y.; Chen, C.; Zhang, L.; Lu, Z.; Chen, S.; Shen, X. Functional characterization of a vanillin dehydrogenase in Corynebacterium glutamicum. Sci. Rep. 2015, 5, 8044. [Google Scholar] [CrossRef]
- Varman, A.M.; He, L.; Follenfant, R.; Wu, W.; Wemmer, S.; Wrobel, S.A.; Tang, Y.J.; Singh, S. Decoding how a soil bacterium extracts building blocks and metabolic energy from ligninolysis provides road map for lignin valorization. Proc. Natl. Acad. Sci. USA 2016, 113, E5802–E5811. [Google Scholar] [CrossRef]
- Bloem, A.; Bertrand, A.; Lonvaud-Funel, A.; de Revel, G. Vanillin production from simple phenols by wine-associated lactic acid bacteria. Lett. Appl. Microbiol. 2007, 44, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Paz, A.; Carballo, J.; Pérez, M.J.; Domínguez, J.M. Bacillus aryabhattai BA03: A novel approach to the production of natural value-added compounds. World J. Microbiol. Biotechnol. 2016, 32, 159. [Google Scholar] [CrossRef] [PubMed]
- Fleige, C.; Hansen, G.; Kroll, J.; Steinbüchel, A. Investigation of the Amycolatopsis sp. strain ATCC 39116 vanillin dehydrogenase and its impact on the biotechnical production of vanillin. Appl. Environ. Microbiol. 2013, 79, 81–90. [Google Scholar] [CrossRef]
- Fleige, C.; Meyer, F.; Steinbüchel, A. Metabolic engineering of the actinomycete Amycolatopsis sp. strain ATCC 39116 towards enhanced production of natural vanillin. Appl. Environ. Microbiol. 2016, 82, 3410–3419. [Google Scholar] [CrossRef]
- Valério, R.; Bernardino, A.R.; Torres, C.A.; Brazinha, C.; Tavares, M.L.; Crespo, J.G.; Reis, M.A. Feeding strategies to optimize vanillin production by Amycolatopsis sp. ATCC 39116. Bioproc. Biosyst. Eng. 2021, 44, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Plaggenborg, R.; Overhage, J.; Steinbüchel, A.; Priefert, H. Functional analyses of genes involved in the metabolism of ferulic acid in Pseudomonas putida KT2440. Appl. Microbiol. Biotechnol. 2003, 61, 528–535. [Google Scholar] [CrossRef]
- Graf, N.; Altenbuchner, J. Genetic engineering of Pseudomonas putida KT2440 for rapid and high-yield production of vanillin from ferulic acid. Appl. Microbiol. Biotechnol. 2014, 98, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Hua, D.; Ma, C.; Song, L.; Lin, S.; Zhang, Z.; Deng, Z.; Xu, P. Enhanced vanillin production from ferulic acid using adsorbent resin. Appl. Microbiol. Biotechnol. 2007, 74, 783–790. [Google Scholar] [CrossRef]
- Yang, W.; Tang, H.; Ni, J.; Wu, Q.; Hua, D.; Tao, F.; Xu, P. Characterization of two Streptomyces enzymes that convert ferulic acid to vanillin. PLoS ONE 2013, 8, e67339. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, D.; Luziatelli, F.; Negroni, A.; Ficca, A.G.; Fava, F.; Ruzzi, M. Metabolic engineering of Pseudomonas fluorescens for the production of vanillin from ferulic acid. J. Biotechnol. 2011, 156, 309–316. [Google Scholar] [CrossRef]
- Chen, H.P.; Chow, M.; Liu, C.C.; Lau, A.; Liu, J.; Eltis, L.D. Vanillin catabolism in Rhodococcus jostii RHA1. Appl. Environ. Microbiol. 2012, 78, 586–588. [Google Scholar] [CrossRef]
- Meyer, F.; Pupkes, H.; Steinbüchel, A. Development of an improved system for the generation of knockout mutants of Amycolatopsis sp. strain ATCC 39116. Appl. Environ. Microbiol. 2017, 83, e02660-e16. [Google Scholar] [CrossRef]
- Simon, O.; Klaiber, I.; Huber, A.; Pfannstiel, J. Comprehensive proteome analysis of the response of Pseudomonas putida KT2440 to the flavor compound vanillin. J. Proteom. 2014, 109, 212–227. [Google Scholar] [CrossRef]
- Chakraborty, D.; Gupta, G.; Kaur, B. Metabolic engineering of E. coli top 10 for production of vanillin through FA catabolic pathway and bioprocess optimization using RSM. Protein Expr. Purification 2016, 128, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.H.; Xie, D.T.; Qiang, S.; Hu, C.Y.; Meng, Y.H. Developing efficient vanillin biosynthesis system by regulating feruloyl-CoA synthetase and enoyl-CoA hydratase enzymes. Appl. Microbiol. Biotechnol. 2022, 106, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, J.W.; Jayaraman, S.S.; Tan, S.G.; Jayaraman, P.; Holowko, M.B.; Zhang, J.; Kang, C.W.; Leo, H.L.; Poh, C.L. A model-driven approach towards rational microbial bioprocess optimization. Biotechnol. Bioeng. 2021, 118, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Li, C.; Kim, J.E.; Lee, S.H.; Yoon, J.Y.; Choi, M.S.; Seo, W.T.; Yang, J.K.; Kim, J.Y.; Kim, S.W. Production of vanillin by metabolically engineered Escherichia coli. Biotechnol. Lett. 2005, 27, 1829–1832. [Google Scholar] [CrossRef]
- Lee, E.G.; Yoon, S.H.; Das, A.; Lee, S.H.; Li, C.; Kim, J.Y.; Choi, M.S.; Oh, D.K.; Kim, S.W. Directing vanillin production from ferulic acid by increased acetyl-CoA consumption in recombinant. Escherichia coli. Biotechnol. Bioeng. 2009, 102, 200–208. [Google Scholar] [CrossRef]
- Overhage, J.; Steinbüchel, A.; Priefert, H. Highly efficient biotransformation of eugenol to ferulic acid and further conversion to vanillin in recombinant strains of Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 6569–6576. [Google Scholar] [CrossRef]
- Ni, J.; Tao, F.; Du, H.; Xu, P. Mimicking a natural pathway for de novo biosynthesis: Natural vanillin production from accessible carbon sources. Sci. Rep. 2015, 5, 13670. [Google Scholar] [CrossRef]
- Brochado, A.R.; Matos, C.; Møller, B.L.; Hansen, J.; Mortensen, U.H.; Patil, K.R. Improved vanillin production in baker’s yeast through in silico design. Microb. Cell Fact. 2010, 9, 1–15. [Google Scholar] [CrossRef]
- Hansen, E.H.; Møller, B.L.; Kock, G.R.; Bünner, C.M.; Kristensen, C.; Jensen, O.R.; Okkels, F.T.; Olsen, C.E.; Motawia, M.S.; Hansen, J. De novo biosynthesis of vanillin in fission yeast (Schizosaccharomyces pombe) and baker’s yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, X.; Lu, X.; He, Y.; Ma, B.; Xu, Y. Efficient biosynthesis of vanillin from isoeugenol by recombinant isoeugenol monooxygenase from Pseudomonas nitroreducens Jin1. Appl. Biochem. Biotechnol. 2021, 193, 1116–1128. [Google Scholar] [CrossRef]
- Zhao, L.; Xie, Y.; Chen, L.; Xu, X.; Zhao, C.X.; Cheng, F. Efficient biotransformation of isoeugenol to vanillin in recombinant strains of Escherichia coli by using engineered isoeugenol monooxygenase and sol-gel chitosan membrane. Process Biochem. 2018, 71, 76–81. [Google Scholar] [CrossRef]
- Yamada, M.; Okada, Y.; Yoshida, T.; Nagasawa, T. Vanillin production using Escherichia coli cells over-expressing isoeugenol monooxygenase of Pseudomonas putida. Biotechnol. Lett. 2008, 30, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Lv, Y.; Yu, H.; Cao, H.; Wang, L.; Wen, B.; Gu, T.; Wang, F.; Sun, L.; Xin, F. Site-directed mutagenesis of coenzyme-independent carotenoid oxygenase CSO2 to enhance the enzymatic synthesis of vanillin. Appl. Microbiol. Biotechnol. 2020, 104, 3897–3907. [Google Scholar] [CrossRef] [PubMed]
- Gallage, N.J.; Jørgensen, K.; Janfelt, C.; Nielsen, A.J.Z.; Naake, T.; Dunski, E.; Dalsten, L.; Grisoni, M.; Møller, B.L. The intracellular localization of the vanillin biosynthetic machinery in pods of Vanilla planifolia. Plant Cell Physiol. 2018, 59, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Barros-Rios, J.; Kourteva, G.; Rao, X.; Chen, F.; Shen, H.; Liu, C.; Podstolski, A.; Belanger, F.; Havkin-Frenkel, D.; et al. A re-evaluation of the final step of vanillin biosynthesis in the orchid Vanilla planifolia. Phytochemistry. 2017, 139, 33–46. [Google Scholar] [CrossRef]
- Chee, M.J.Y.; Lycett, G.W.; Khoo, T.J.; Chin, C.F. Bioengineering of the plant culture of Capsicum frutescens with vanillin synthase gene for the production of vanillin. Mol. Biotechnol. 2017, 59, 1–8. [Google Scholar] [CrossRef]
- Arya, S.S.; Mahto, B.K.; Sengar, M.S.; Rookes, J.E.; Cahill, D.M.; Lenka, S.K. Metabolic engineering of rice cells with vanillin synthase gene (VpVAN) to produce vanillin. Mol. Biotechnol. 2022, 64, 861–872. [Google Scholar] [CrossRef]
- Jiang, W.; Gao, H.; Sun, J.; Yang, X.; Jiang, Y.; Zhang, W.; Jiang, M.; Xin, F. Current status, challenges and prospects for lignin valorization by using Rhodococcus sp. Biotechnol. Adv. 2022, 60, 108004. [Google Scholar] [CrossRef]
- Brown, M.E.; Chang, M.C. Exploring bacterial lignin degradation. Curr. Opin. Chem. Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef]
- Lundell, T.K.; Mäkelä, M.R.; Hildén, K. Lignin-modifying enzymes in filamentous basidiomycetes-ecological, functional and phylogenetic review. J. Basic Microbiol. 2010, 50, 5–20. [Google Scholar] [CrossRef]
- Karode, B.; Patil, U.; Jobanputra, A. Biotransformation of low cost lignocellulosic substrate into vanillin by white rot fungus, Phanerochaete chrysosporium NCIM 1197. Indian J. Biotechnol. 2013, 12, 281–283. [Google Scholar]
- Baghel, S.; Anandkumar, J. Biodepolymerization of kraft lignin for production and optimization of vanillin using mixed bacterial culture. Bioresour. Technol. Rep. 2019, 8, 100335. [Google Scholar] [CrossRef]
- Zuo, K.; Li, H.; Chen, J.; Ran, Q.; Huang, M.; Cui, X.; He, L.; Liu, J.; Jiang, Z. Effective biotransformation of variety of guaiacyl lignin monomers into vanillin by Bacillus pumilus. Front. Microbiol. 2022, 13, 901690. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, P.D.; Hardiman, E.M.; Ahmad, M.; Otani, H.; Seghezzi, N.; Eltis, L.D.; Bugg, T.D. Breaking down lignin to high-value chemicals: The conversion of lignocellulose to vanillin in a gene deletion mutant of Rhodococcus jostii RHA1. ACS Chem. Biol. 2013, 8, 2151–2156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Xu, L.; Sethupathy, S.; Si, H.; Ahmad, F.; Zhang, R.; Zhang, W.; Yang, B.; Sun, J. Decoding lignin valorization pathways in the extremophilic Bacillus ligniniphilus L1 for vanillin biosynthesis. Green Chem. 2021, 23, 9554–9570. [Google Scholar] [CrossRef]
- Xu, Z.; Lei, P.; Zhai, R.; Wen, Z.; Jin, M. Recent advances in lignin valorization with bacterial cultures: Microorganisms, metabolic pathways, and bio-products. Biotechnol. Biofuels 2019, 12, 32. [Google Scholar] [CrossRef]
- Zamzuri, N.A.; Abd-Aziz, S. Biovanillin from agro wastes as an alternative food flavour. J. Sci. Food. Agric. 2013, 93, 429–438. [Google Scholar] [CrossRef]
- Harshvardhan, K.; Suri, M.; Goswami, A.; Goswami, T. Biological approach for the production of vanillin from lignocellulosic biomass (Bambusa tulda). J. Clean. Prod. 2017, 149, 485–490. [Google Scholar] [CrossRef]
- Chattopadhyay, P.; Banerjee, G.; Sen, S.K. Cleaner production of vanillin through biotransformation of ferulic acid esters from agroresidue by Streptomyces sannanensis. J. Clean. Prod. 2018, 182, 272–279. [Google Scholar] [CrossRef]
- Chakraborty, D.; Selvam, A.; Kaur, B.; Wong, J.W.C.; Karthikeyan, O.P. Application of recombinant Pediococcus acidilactici BD16 (fcs+/ech+) for bioconversion of agrowaste to vanillin. Appl. Microbiol. Biotechnol. 2017, 101, 5615–5626. [Google Scholar] [CrossRef]
- Zheng, L.; Zheng, P.; Sun, Z.; Bai, Y.; Wang, J.; Guo, X. Production of vanillin from waste residue of rice bran oil by Aspergillus niger and Pycnoporus cinnabarinus. Bioresour. Technol. 2007, 98, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Barghini, P.; Di Gioia, D.; Fava, F.; Ruzzi, M. Vanillin production using metabolically engineered Escherichia coli under non-growing conditions. Microb. Cell Fact. 2007, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lesage-Meessen, L.; Lomascolo, A.; Bonnin, E.; Thibault, J.F.; Buleon, A.; Roller, M.; Asther, M.; Record, E.; Ceccaldi, B.C.; Asther, M. A biotechnological process involving filamentous fungi to produce natural crystalline vanillin from maize bran. Appl. Biochem. Biotechnol. 2002, 102, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Lesage-Meessen, L.; Stentelaire, C.; Lomascolo, A.; Couteau, D.; Asther, M.; Moukha, S.; Moukha, S.; Record, E.; Sigoillot, J.; Asther, M. Fungal transformation of ferulic acid from sugar beet pulp to natural vanillin. J. Sci. Food Agric. 1999, 79, 487–490. [Google Scholar] [CrossRef]
- Rabenhorst, J. Production of methoxyphenol-type natural aroma chemicals by biotransformation of eugenol with a new Pseudomonas sp. Appl. Microbiol. Biotechnol. 1996, 46, 470–474. [Google Scholar] [CrossRef]
- Plaggenborg, R.; Overhage, J.; Loos, A.; Archer, J.A.; Lessard, P.; Sinskey, A.J.; Steinbüchel, A.; Priefert, H. Potential of Rhodococcus strains for biotechnological vanillin production from ferulic acid and eugenol. Appl. Microbiol. Biotechnol. 2006, 72, 745–755. [Google Scholar] [CrossRef]
- Khanafari, A.; Olia, M.S.J.; Sharifnia, F. Bioconversion of essential oil from plants with eugenol bases to vanillin by Serratia marcescens. J. Essent. Oil Bear. Plants 2011, 14, 229–240. [Google Scholar] [CrossRef]
- Singh, A.; Mukhopadhyay, K.; Ghosh Sachan, S. Enhanced vanillin production from eugenol by Bacillus cereus NCIM-5727. Bioproc. Biosyst. Eng. 2022, 45, 1811–1824. [Google Scholar] [CrossRef]
- Hua, D.; Ma, C.; Lin, S.; Song, L.; Deng, Z.; Maomy, Z.; Zhang, Z.; Yu, B.; Xu, P. Biotransformation of isoeugenol to vanillin by a newly isolated Bacillus pumilus strain: Identification of major metabolites. J. Biotechnol. 2007, 130, 463–470. [Google Scholar] [CrossRef]
- Paz, A.; Costa-Trigo, I.; Tugores, F.; Míguez, M.; de la Montaña, J.; Domínguez, J.M. Biotransformation of phenolic compounds by Bacillus aryabhattai. Bioproc. Biosyst. Eng. 2019, 42, 1671–1679. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, P.; Han, S.; Yan, H.; Ma, C. Metabolism of isoeugenol via isoeugenol-diol by a newly isolated strain of Bacillus subtilis HS8. Appl. Microbiol. Biotechnol. 2006, 73, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Ashengroph, M.; Nahvi, I.; Zarkesh-Esfahani, H.; Momenbeik, F. Use of growing cells of Pseudomonas aeruginosa for synthesis of the natural vanillin via conversion of isoeugenol. Iran. J. Pharm. Res. 2011, 10, 749–757. [Google Scholar] [PubMed]
- Haridoss, M.; Kamatchi, C.; Rafiq, Z.; Vaidyanathan, R. Biotransformation of isoeugenol to vanillin by beneficial bacteria isolated from the soil of aromatic plants. J. Chem. Pharm. Res. 2015, 7, 274–280. [Google Scholar]
- Unno, T.; Kim, S.J.; Kanaly, R.A.; Ahn, J.H.; Kang, S.I.; Hur, H.G. Metabolic characterization of newly isolated Pseudomonas nitroreducens Jin1 growing on eugenol and isoeugenol. J. Agric. Food. Chem. 2007, 55, 8556–8561. [Google Scholar] [CrossRef]
- Yamada, M.; Okada, Y.; Yoshida, T.; Nagasawa, T. Biotransformation of isoeugenol to vanillin by Pseudomonas putida IE27 cells. Appl. Microbiol. Biotechnol. 2007, 73, 1025–1030. [Google Scholar] [CrossRef]
- Li, Y.H.; Sun, Z.H.; Zhao, L.Q.; Xu, Y. Bioconversion of isoeugenol into vanillin by crude enzyme extracted from soybean. Appl. Biochem. Biotechnol. 2005, 125, 1–10. [Google Scholar] [CrossRef]
- Ryu, J.Y.; Seo, J.; Park, S.; Ahn, J.H.; Chong, Y.; Sadowsky, M.J.; Hur, H.G. Characterization of an isoeugenol monooxygenase (Iem) from Pseudomonas nitroreducens Jin1 that transforms isoeugenol to vanillin. Biosci. Biotechnol. Biochem. 2013, 77, 289–294. [Google Scholar] [CrossRef]
- Yamada, M.; Okada, Y.; Yoshida, T.; Nagasawa, T. Purification, characterization and gene cloning of isoeugenol-degrading enzyme from Pseudomonas putida IE27. Arch. Microbiol. 2007, 187, 511–517. [Google Scholar] [CrossRef]
- Zhao, L.; Jiang, Y.; Fang, H.; Zhang, H.; Cheng, S.; Rajoka, M.S.R.; Wu, Y. Biotransformation of isoeugenol into vanillin using immobilized recombinant cells containing isoeugenol monooxygenase active aggregates. Appl. Biochem. Biotechnol. 2019, 189, 448–458. [Google Scholar] [CrossRef]
- Lu, X.Y.; Wu, X.M.; Ma, B.D.; Xu, Y. Enhanced thermostability of Pseudomonas nitroreducens isoeugenol monooxygenase by the combinatorial strategy of surface residue replacement and consensus mutagenesis. Catalysts 2021, 11, 1199. [Google Scholar] [CrossRef]
- Fitzgerald, D.J.; Stratford, M.; Gasson, M.J.; Narbad, A. Structure-function analysis of the vanillin molecule and its antifungal properties. J. Agric. Food Chem. 2005, 53, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.R.; Lee, J.H.; Moon, Y.M.; Choi, T.R.; Yang, S.Y.; Song, H.S.; Park, J.Y.; Park, Y.L.; Bhatia, S.K.; Gurav, R.; et al. Increased tolerance to furfural by introduction of polyhydroxybutyrate synthetic genes to Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.R.; Song, H.S.; Han, Y.H.; Park, Y.L.; Park, J.Y.; Yang, S.Y.; Bhatia, S.K.; Gurav, R.; Kim, H.J.; Lee, Y.K.; et al. Enhanced tolerance to inhibitors of Escherichia coli by heterologous expression of cyclopropane-fatty acid-acyl-phospholipid synthase (cfa) from Halomonas socia. Bioprocess Biosyst. Eng. 2020, 43, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liang, Z.; Hou, J.; Bao, X.; Shen, Y. Identification and functional evaluation of the reductases and dehydrogenases from Saccharomyces cerevisiae involved in vanillin resistance. BMC. Biotechnol. 2016, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Ashengroph, M.; Amini, J. Bioconversion of isoeugenol to vanillin and vanillic acid using the resting cells of Trichosporon asahii. 3 Biotech. 2017, 7, 358. [Google Scholar] [CrossRef]
- Valério, R.; Brazinha, C.; Crespo, J.G. Comparative analysis of bio-vanillin recovery from bioconversion media using pervaporation and vacuum distillation. Membranes 2022, 12, 801. [Google Scholar] [CrossRef]
- Zabkova, M.; da Silva, E.B.; Rodrigues, A.E. Recovery of vanillin from Kraft lignin oxidation by ion-exchange with neutralization. Sep. Purif. Technol. 2007, 55, 56–68. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Jiang, Z.T.; Gao, H.J.; Li, R. Recovery of vanillin from aqueous solutions using macroporous adsorption resins. Eur. Food Res. Technol. 2008, 226, 377–383. [Google Scholar] [CrossRef]
- Khwanjaisakun, N.; Amornraksa, S.; Simasatitkul, L.; Charoensuppanimit, P.; Assabumrungrat, S. Techno-economic analysis of vanillin production from Kraft lignin: Feasibility study of lignin valorization. Bioresour. Technol. 2020, 299, 122559. [Google Scholar] [CrossRef]
- Smink, D.; Kersten, S.R.; Schuur, B. Recovery of lignin from deep eutectic solvents by liquid-liquid extraction. Sep. Purif. Technol. 2020, 235, 116127. [Google Scholar] [CrossRef]
- Brazinha, C.; Barbosa, D.S.; Crespo, J.G. Sustainable recovery of pure natural vanillin from fermentation media in a single pervaporation step. Green Chem. 2011, 13, 2197–2203. [Google Scholar] [CrossRef]
- Pavón, S.; Blaesing, L.; Jahn, A.; Aubel, I.; Bertau, M. Liquid membranes for efficient recovery of phenolic compounds such as vanillin and catechol. Membranes 2020, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Dong, W.; Xin, F.; Jiang, M. Designing synthetic microbial consortia for biofuel production. Trends Biotechnol. 2020, 38, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Bharagava, R.N. Bacterial degradation of synthetic and kraft lignin by axenic and mixed culture and their metabolic products. J. Environ. Biol. 2013, 34, 991. [Google Scholar] [PubMed]
- Cai, T.; Sun, H.; Qiao, J.; Zhu, L.; Zhang, F.; Zhang, J.; Tang, Z.; Wei, X.; Yang, J.; Yuan, Q.; et al. Cell-free chemoenzymatic starch synthesis from carbon dioxide. Science 2021, 373, 1523–1527. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Strain | Main Strategies | Production | References |
|---|---|---|---|---|
| Eugenol | Recombinant Escherichia coli | Coexpression of vaoA, calA, calB, fcs, and ech genes. | 0.3 g/L | [36] |
| Ferulic acid | Recombinant Escherichia coli JM109-FE-F | Construction of an efficient cell-free catalytic system with FCS-Str and ECH-Str combination at 1:1; using resting cells | 2.3 g/L | [32] |
| Recombinant Escherichia coli FR13 | Integration of Fcs and Ech onto chromosomes; using resting cells; fed-batch fermentation; using a two-phase (solid–liquid) system | 4.3 g/L | [4] | |
| Recombinant Escherichia coli top 10 | Introduction of the cloned vanillin biosynthetic gene cassette in the pCCIBAC expression vector | 0. 068 g/L | [32] | |
| Recombinant Escherichia coli | Using a model-driven approach to fine-tune nutrients | 0.91 g/L | [33] | |
| Recombinant Escherichia coli | Using a two-pot bioprocess; introduction of fdc and cso2 genes; designing the cultivation medium | 7.9 g/L | [10] | |
| Recombinant Escherichia coli NTG-VR1 | Production of vanillin plasmid pTAHEF containing fcs and ech genes; using NTG mutagenesis; employing 50% (w/v) of XAD-2 resin | 2.9 g/L | [34] | |
| Recombinant Escherichia coli | Coexpression of gltA, icdA, fcs, and ech genes. | 5.1 g/L | [35] | |
| L-tyrosine, glucose, xylose, glycerol | Recombinant Escherichia coli | Mimicking the construction of the phenylpropanoid pathway in microorganisms and inducing five enzymes | 0.097 g/L, 0.019 g/L, 0.013 g/L, 0.024 g/L | [37] |
| Glucose | Recombinant Saccharomyces cerevisiae | Using an in silico strategy based on the strain S. cerevisiae | 0.500 g/L | [38] |
| Recombinant Saccharomyces cerevisiae | Coexpression of four genes: 3DSD, ACAR, OMT, and UGT | 0.045 g/L | [39] | |
| Isoeugenol | Recombinant Escherichia coli | Overexpression of isoeugenol monooxygenase; employing the magnetic chitosan membrane | 38.3 g/L | [40] |
| Recombinant Escherichia coli | Expression of the IEM720 gene; employing the sol–gel chitosan membrane | 4.5 g/L | [41] | |
| Recombinant Escherichia coli | Introduction of a plasmid with the isoeugenol monooxygenase gene | 28.3 g/L | [42] |
| Microorganism | Substrate | Time | Yield | References |
|---|---|---|---|---|
| Pediococcus acidilactici BD16 (fcs+/ech+) | Rice bran containing 0.257 mM FA | 24 h | 4.0 g/L | [60] |
| Aspergillus niger CGMCC0774 and Pycnoporus cinnabarinus CGMCC1115 | Rice bran oil | 72 h | 2.8 g/L | [61] |
| Streptomyces sannanensis MTCC 6637 | Wheat bran | 5 d | 0.71 g/L | [59] |
| E. coli strain JM109(pBB1) | Wheat bran containing ferulic acid | - | 2.5 g/L | [62] |
| 14 natural bacterial consortium | Bamboo chips from Bambusa tulda (ligno-cellulosic biomass) | 8 d | 0.9 g/L | [58] |
| Aspergillus niger I -1472 and Pycnoporus cinnabarinus MUCL 39532 | Sugar beet pulp | 8 d | 0.11 g/L | [63] |
| A. niger I-1472 and P. cinnabarinus MUCL39533 | Maize bran | 7–8 d | 0.77 g/L | [64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Chen, X.; Feng, Y.; Sun, J.; Jiang, Y.; Zhang, W.; Xin, F.; Jiang, M. Current Status, Challenges, and Prospects for the Biological Production of Vanillin. Fermentation 2023, 9, 389. https://doi.org/10.3390/fermentation9040389
Jiang W, Chen X, Feng Y, Sun J, Jiang Y, Zhang W, Xin F, Jiang M. Current Status, Challenges, and Prospects for the Biological Production of Vanillin. Fermentation. 2023; 9(4):389. https://doi.org/10.3390/fermentation9040389
Chicago/Turabian StyleJiang, Wankui, Xiaoyue Chen, Yifan Feng, Jingxiang Sun, Yujia Jiang, Wenming Zhang, Fengxue Xin, and Min Jiang. 2023. "Current Status, Challenges, and Prospects for the Biological Production of Vanillin" Fermentation 9, no. 4: 389. https://doi.org/10.3390/fermentation9040389
APA StyleJiang, W., Chen, X., Feng, Y., Sun, J., Jiang, Y., Zhang, W., Xin, F., & Jiang, M. (2023). Current Status, Challenges, and Prospects for the Biological Production of Vanillin. Fermentation, 9(4), 389. https://doi.org/10.3390/fermentation9040389