
Improving Pulping Performance as Well as Reducing Consumption and Increasing Efficiency via Microbial Consortium Pretreating Bamboo
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Process
2.2.1. Microbiological Pretreatment of Bamboo Chips
2.2.2. Conventional Kraft Cooking, Hand Paper-Making
2.2.3. Modified Kraft Cooking
2.3. Chemical Components Determination of Pulp
2.4. Fiber Morphology and Properties of Hand Papermaking
2.5. XRD Characterization
2.6. 16S rRNA Gene Sequencing and Functional Prediction
2.6.1. 16S rRNA Gene Sequencing
2.6.2. Enzyme Functional Prediction
2.6.3. Bioinformatics Analysis
3. Results and Discussion
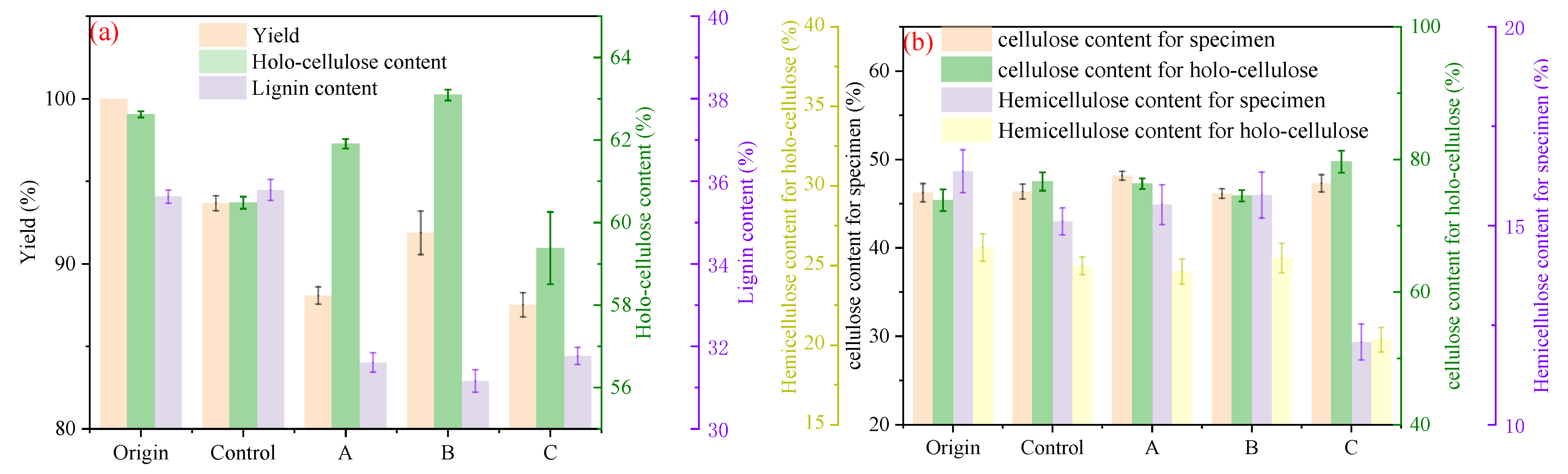
3.1. Chemical Components and Physical Structure of Bamboo via Microbiological Pretreatment
3.2. 16S rRNA Sequencing and Analysis
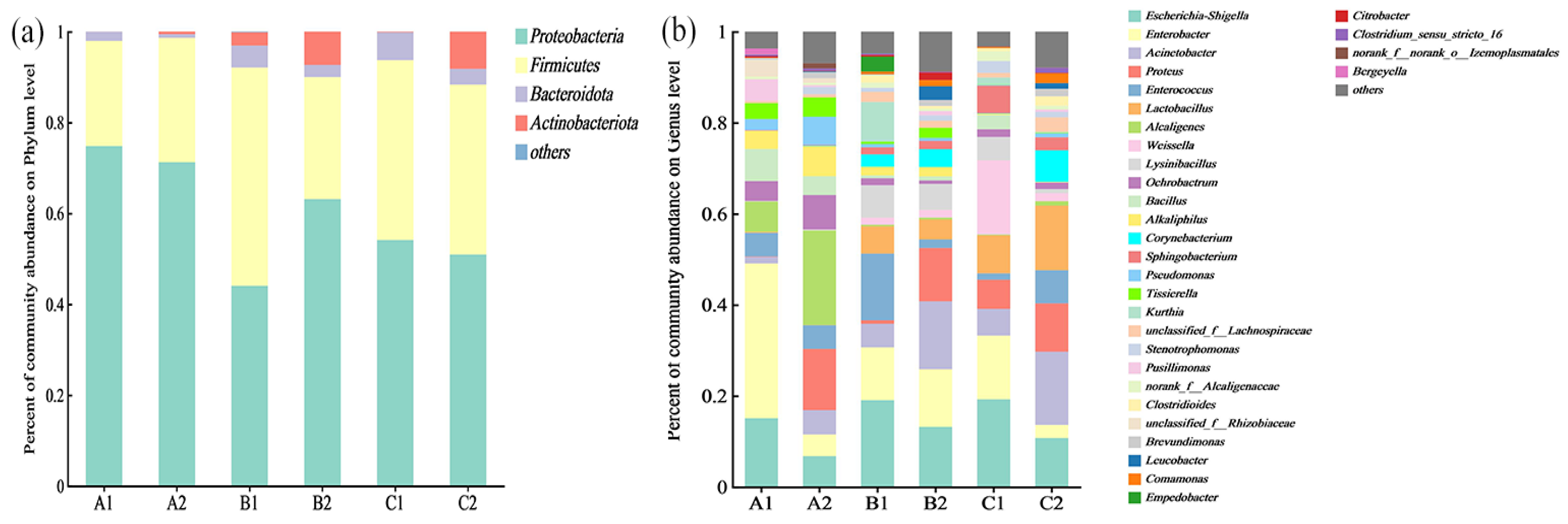
3.2.1. Differences in Microbial Consortium Composition and Species Distribution
3.2.2. Enzyme Annotation Analysis Related to Lignocellulose Degradation
3.3. Effect of Microbial Pretreatment on Bamboo Bio-Refining
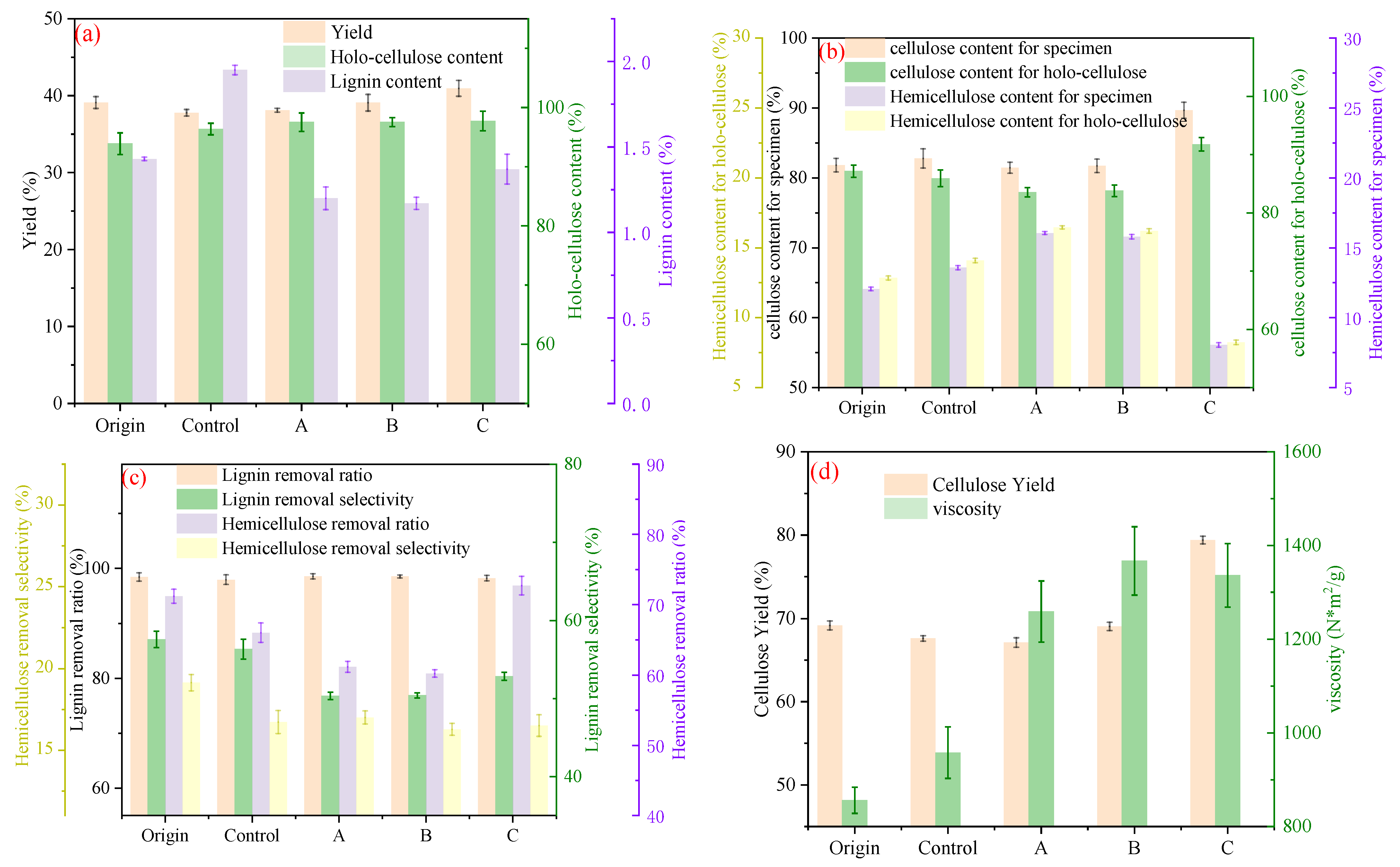
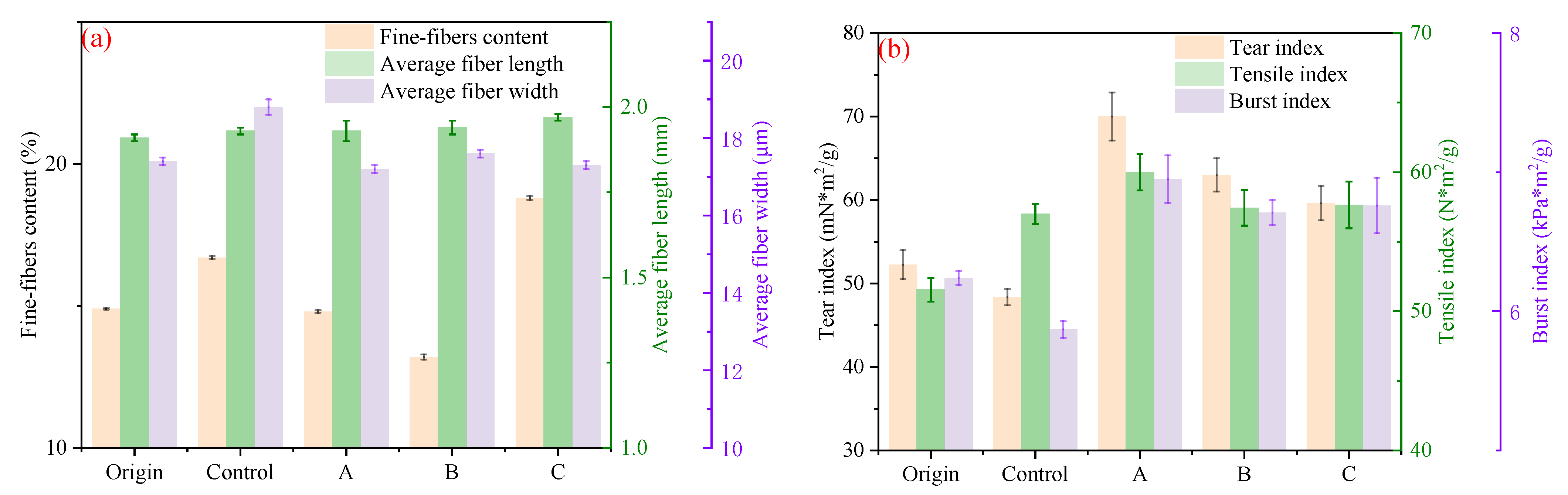
3.3.1. Effect from the Perspective of Pulping Product Characteristics
3.3.2. Effect of Pulping Process on the Potential for Reducing Consumption and Increasing Efficiency
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hendriks, A.T.W.M.; Zeeman, G. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol. 2009, 100, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, L.; Liu, D. Biomass recalcitrance. Part I: The chemical compositions and physical structures affecting the enzymatic hydrolysis of lignocellulose. Biofuels Bioprod. Biorefin. 2012, 6, 465–482. [Google Scholar] [CrossRef]
- Gautam, A.; Kumar, A.; Dutt, D. Effects of Ethanol Addition and Biological Pretreatment on Soda Pulping of Eulaliopsis binata. J. Biomater. Nanobiotechnol. 2016, 7, 78. [Google Scholar] [CrossRef]
- Singh, P.; Sulaiman, O.; Hashim, R.; Rupani, P.F.; Peng, L.C. Biopulping of lignocellulosic material using different fungal species: A review. Rev. Environ. Sci. Bio/Technol. 2010, 9, 141–151. [Google Scholar] [CrossRef]
- Adaskaveg, J.E.; Gilbertson, R.L.; Dunlap, M.R. Effects of Incubation Time and Temperature on In Vitro Selective Delignification of Silver Leaf Oak by Ganoderma colossum. Appl. Environ. Microbiol. 1995, 61, 138–144. [Google Scholar] [CrossRef]
- Behrendt, C.; Blanchette, R. Biological Processing of Pine Logs for Pulp and Paper Production with Phlebiopsis gigantea. Appl. Environ. Microbiol. 1997, 63, 1995–2000. [Google Scholar] [CrossRef]
- Gupta, R.; Saini, V.K.; Bhatt, R.P.; Thapliyal, B.; Naithani, S. Influence of mechanical operation on the biodelignification of eucalyptus tereticornis by trametes versicolor. Cellul. Chem. Technol. 2013, 47, 759–764. [Google Scholar]
- Helmy, S.M.; El-Meligi, M. Biopulping and Biobleaching by white rot fungi. J. Sci. Ind. Res. 2002, 61, 376–381. [Google Scholar]
- Sahin, H.T. Base-catalyzed organosolv pulping of jute. J. Chem. Technol. Biotechnol. 2003, 78, 1267–1273. [Google Scholar] [CrossRef]
- Singh, D.; Shulin, C. The White-Rot Fungus Phanerochaete chrysosporium: Conditions for the Production of Lignin-Degrading Enzymes. Appl. Microbiol. Biotechnol. 2008, 81, 399–417. [Google Scholar] [CrossRef]
- Lei, W.; Wangui, W.; Xiang, J.; Cai, L. Biodegradation of lignin by the white rot fungus Polyporus varius and its promising potential for biopulping. In Proceedings of the 2011 International Conference on Materials for Renewable Energy and Environment—ICMREE2011, Shanghai, China, 20–22 May 2011; Volume 1. [Google Scholar]
- Saini, V.K.; Naithani, S.; Thapliyal, B.P.; Gupta, R. Increased delignification rate of Dendrocalamus strictus (Roxburgh) nees by Schizophyllum commune Fr.; Fr. to reduce chemical consumption during pulping process. Songklanakarin J. Sci. Technol. 2013, 35, 415–420. [Google Scholar]
- Hatakka, A. Lignin-modifying enzymes from selected white-rot fungi: Production and role from in lignin degradation. FEMS Microbiol. Rev. 1994, 13, 125–135. [Google Scholar] [CrossRef]
- Dai, Y.Y.; Zhong, W.H. Research Process on Degradation of Lignocellulose by bacteria. Chem. Bioeng. 2016, 33, 11–16. [Google Scholar]
- Xu, R.; Zhang, K.; Liu, p.; Han, H.; Zhao, S.; Kakade, A.; Khan, A.; Du, D.; Li, X. Lignin depolymerization and utilization by bacteria. Bioresour. Technol. 2018, 269. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass—An overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef]
- Passos, F.; Uggetti, E.; Carrere, H.; Ferrer, I. Pretreatment of microalgae to improve biogas production: A review. Bioresour. Technol. 2014, 172, 403–412. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.; Liu, J.; Yang, X.; Ren, Y.; Miao, H.; Pan, Y.; Lv, J.; Yan, G.; Ding, L.; et al. Exploring the mechanisms of organic matter degradation and methane emission during sewage sludge composting with added vesuvianite: Insights into the prediction of microbial metabolic function and enzymatic activity. Bioresour. Technol. 2019, 286, 121397. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Yan, L.; Gao, Y.; Wang, Y.; Wang, W. A novel lignin degradation bacterial consortium for efficient pulping. Bioresour. Technol. 2013, 139, 113–119. [Google Scholar] [CrossRef]
- Han, H.M. Construction and Application of Bacillus licheniformis with Selective Degradation of the Bamboo Component; Qilu University of Technology: Jinan, China, 2017. [Google Scholar]
- Chen, Z.; Zhang, H.; He, Z.; Zhang, L.; Yue, X. Bamboo as an emerging resource for worldwide pulping and papermaking. BioResources 2018, 14, 3–5. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Duan, C.; Liu, Y.; Ni, Y. Enhancing hemicelluloses removal from a softwood sulfite pulp. Bioresour. Technol. 2015, 192, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.D.; Creely, J.J.; Martin, A.E.; Conrad, C.M. An Empirical Method for Estimating the Degree of Crystallinity of Native Cellulose Using the X-Ray Diffractometer. Text. Res. J. 1959, 29, 786–794. [Google Scholar] [CrossRef]
- French, A. Idealized powder diffraction patterns for cellulose polymorphs. Cellulose 2014, 21. [Google Scholar] [CrossRef]
- Ma, S.; Sun, X.; Fang, C.; He, X.; Han, L.; Huang, G. Exploring the mechanisms of decreased methane during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane. Waste Manag. 2018, 78, 393–400. [Google Scholar] [CrossRef]
- Lan, Y.; Wang, Q.; Cole, J.R.; Rosen, G.L. Using the RDP Classifier to Predict Tanonomic Novelty and Reduce the Search Space for Fingding Novel Organisms. PLoS ONE 2012, 7, e32491. [Google Scholar] [CrossRef]
- Kong, Z.Y.; Wu, Z.J.; Glick, B.R.; He, S.Y.; Huang, C.; Wu, L. Co-occurrence patterns of microbial communities affected by inoculants of plant growth-promoting bacteria during phytoremediation of heavy metal-contaminated soils. Ecotoxicol. Environ. Saf. 2019, 183, 1–10. [Google Scholar] [CrossRef]
- Fan, L.F.; Wang, Z.L.; Chen, M.S.; Qu, Y.X.; Li, J.Y.; Zhou, A.G.; Xie, S.L.; Zeng, F.; Zou, J.X. Microbiota comparison of Pacific white shrimp intestine and sediment at freshwater and marine cultured environment. Sci. Total Environ. 2019, 657, 1197–1204. [Google Scholar] [CrossRef]
- Chandra, R.P.; Bura, R.; Mabee, W.; Berlin, A.K.; Pan, X.; Saddler, J.N. Substrate pretreatment: The key to effective enzymatic hydrolysis of lignocellulosics? Adv. Biochem. Eng./Biotechnol. 2007, 108, 67–93. [Google Scholar]
- Yang, B.; Wyman, C. Pretreatment: The key to unlocking low-cost cellulosic ethanol. Biofuels Bioprod. Biorefin. 2008, 2, 26–40. [Google Scholar] [CrossRef]
- Hall, M.; Bansal, P.; Lee, J.; Realff, M.; Bommarius, A. Cellulose crystallinity—A key predictor of the enzymatic hydrolysis rate. FEBS J. 2010, 277, 1571–1582. [Google Scholar] [CrossRef]
- Zhou, Z.; Qiao, W.; Xing, C.; An, Y.; Shen, X.; Ren, W.; Jiang, L.-M.; Wang, L. Microbial community structure of anoxic–oxic-settling-anaerobic sludge reduction process revealed by 454-pyrosequencing. Chem. Eng. J. 2015, 266, 249–257. [Google Scholar] [CrossRef]
- Ma, S.-J.; Ding, L.-L.; Huang, H.; Geng, J.-J.; Xu, K.; Zhang, Y.; Ren, H. Effects of DO levels on surface force, cell membrane properties and microbial community dynamics of activated sludge. Bioresour. Technol. 2016, 214, 645–652. [Google Scholar] [CrossRef]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Singh, R. The emerging role for bacteria in lignin degradation and bio-product formation. Curr. Opin. Biotechnol. 2010, 22, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Rui, J.; Peng, J.; Lu, Y. Succession of Bacterial Populations during Plant Residue Decomposition in Rice Field Soil. Appl. Environ. Microbiol. 2009, 75, 4879–4886. [Google Scholar] [CrossRef] [PubMed]
- Blanc, M.; Marilley, L.; Beffa, T.; Aragno, M. Thermophilic Bacterial Communities in Hot Composts as Revealed by Most Probable Number Counts and Molecular (16S rDNA) Methods. FEMS Microbiol. Ecol. 1999, 28, 141–149. [Google Scholar] [CrossRef]
- Singh, Y.-S.; Singh, J.; Singh, G.; Govinder, S.; Ladha, J. Placement effects on rice residue decomposition and nutrient dynamics on two soil types during wheat cropping in rice–wheat system in northwestern India. Nutr. Cycl. Agroecosyst. 2010, 88, 471–480. [Google Scholar] [CrossRef]
- Patel, A.; Falck, P.; Shah, N.; Immerzeel, P.; Adlercreutz, P.; Stålbrand, H.; Prajapati, J.B.; Holst, O.; Nordberg Karlsson, E. Evidence for xylooligosaccharide utilization in Weissella strains isolated from Indian fermented foods and vegetables. FEMS Microbiol. Lett. 2013, 346, 20–28. [Google Scholar] [CrossRef]
- Cheng, J.F.; Guo, J.X.; Bian, Y.N.; Chen, Z.L.; Li, C.L.; Li, X.D.; Li, Y.H. Sphingobacterium athyrii sp. nov., a cellulose- and xylan-degrading bacterium isolated from a decaying fern (Athyrium wallichianum Ching). Int. J. Syst. Evol. Microbiol. 2019, 69, 752–760. [Google Scholar] [CrossRef]
- Ding, W.; Si, M.; Zhang, W.; Zhang, Y.; Chen, C.; Zhang, L.; Lu, Z.; Chen, S.; Shen, X. Functional characterization of a vanillin dehydrogenase in Corynebacterium glutamicum. Sci. Rep. 2015, 5, 8044. [Google Scholar] [CrossRef]
- Granja, R.; Wilkinson, R.; Persinoti, G.; Squina, F.; Fülöp, V.; Bugg, T. Structural and Functional Characterisation of Multi-copper Oxidase CueO from Lignin-Degrading Bacterium Ochrobactrum sp. Reveal its Activity towards Lignin Model Compounds and Lignosulfonate. FEBS J. 2018, 285, 1684–1700. [Google Scholar] [CrossRef]
- Ahmed, A.A.Q.; Babalola, O.; McKay, T. Cellulase- and Xylanase-Producing Bacterial Isolates with the Ability to Saccharify Wheat Straw and Their Potential Use in the Production of Pharmaceuticals and Chemicals from Lignocellulosic Materials. Waste Biomass Valorization 2018, 9, 765–775. [Google Scholar] [CrossRef]
- Fischer, W.; Mayr, M.; Spirk, S.; Reishofer, D.; Jagiello, L.; Schmiedt, R.; Colson, J.; Zankel, A.; Bauer, W. Pulp Fines—Characterization, Sheet Formation, and Comparison to Microfibrillated Cellulose. Polymers 2017, 9, 366. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Li, Z.; Guan, X.; Xu, Y.; Huang, N.; Liu, K. Improving Pulping Performance as Well as Reducing Consumption and Increasing Efficiency via Microbial Consortium Pretreating Bamboo. Fermentation 2023, 9, 400. https://doi.org/10.3390/fermentation9040400
Liu C, Li Z, Guan X, Xu Y, Huang N, Liu K. Improving Pulping Performance as Well as Reducing Consumption and Increasing Efficiency via Microbial Consortium Pretreating Bamboo. Fermentation. 2023; 9(4):400. https://doi.org/10.3390/fermentation9040400
Chicago/Turabian StyleLiu, Chun, Zhijian Li, Xiuqiong Guan, Yang Xu, Nan Huang, and Kui Liu. 2023. "Improving Pulping Performance as Well as Reducing Consumption and Increasing Efficiency via Microbial Consortium Pretreating Bamboo" Fermentation 9, no. 4: 400. https://doi.org/10.3390/fermentation9040400
APA StyleLiu, C., Li, Z., Guan, X., Xu, Y., Huang, N., & Liu, K. (2023). Improving Pulping Performance as Well as Reducing Consumption and Increasing Efficiency via Microbial Consortium Pretreating Bamboo. Fermentation, 9(4), 400. https://doi.org/10.3390/fermentation9040400