
Potential Capacity of Candida wangnamkhiaoensis to Produce Oleic Acid

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Preparation of the Inoculum
2.3. Kinetics of Cell Growth, Substrate Consumption, and Lipid Production
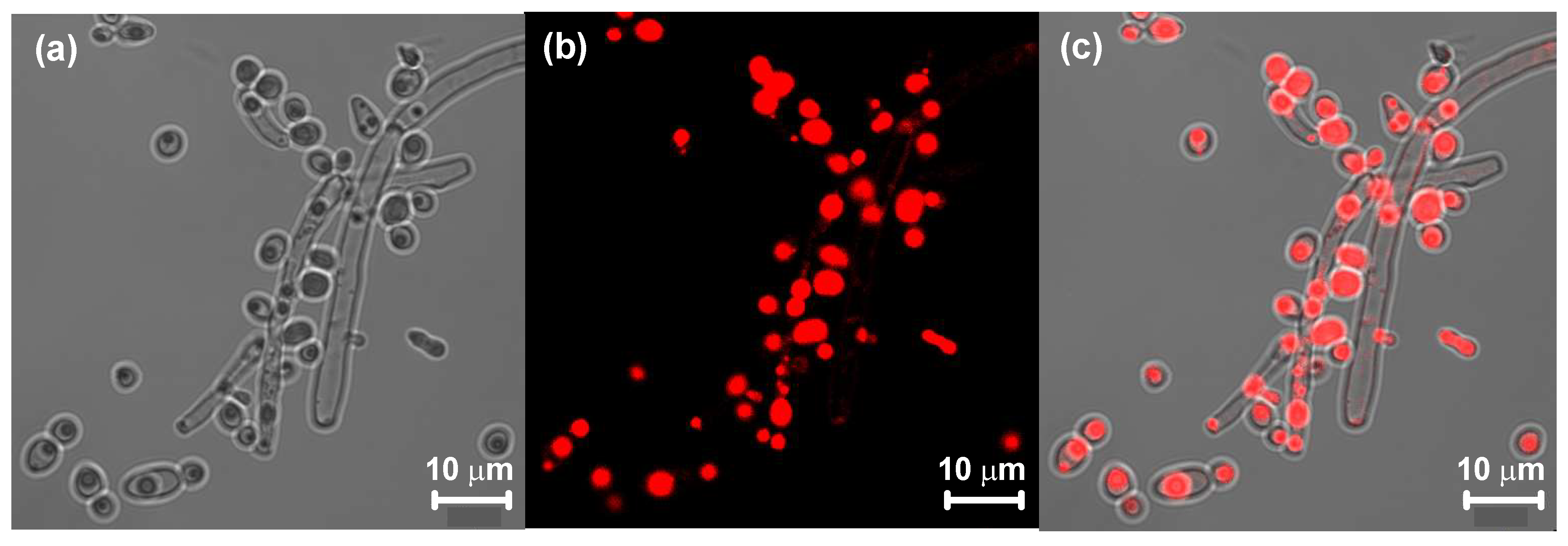
2.4. Detection of Lipids by Confocal Laser Scanning Microscopy (CLSM)
2.5. Quantification of Biomass, Glucose, Glycerol, and Lipids
2.6. Determination of the Profile of Fatty Acids and the Quantity of Each One
2.7. Kinetic Parameters
2.8. Statistical Analysis
3. Results and Discussion
3.1. Detection of Lipids by Confocal Laser Scanning Microscopy (CLSM)
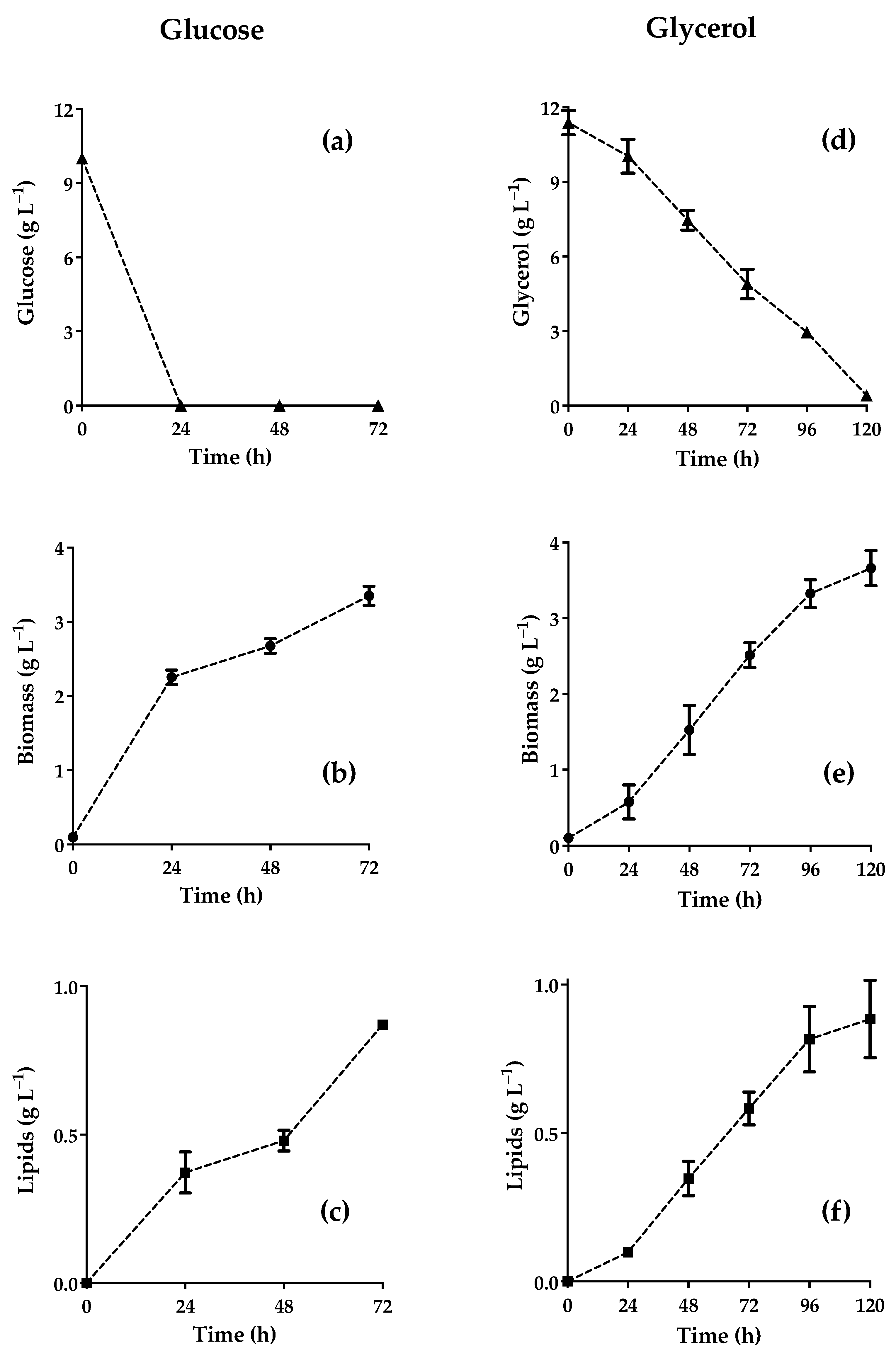
3.2. Kinetics of Cell Growth, Substrate Consumption, and Lipid Production by CW
3.3. Kinetic Parameters of Cellular Growth, Substrate Consumption, and Lipid Production
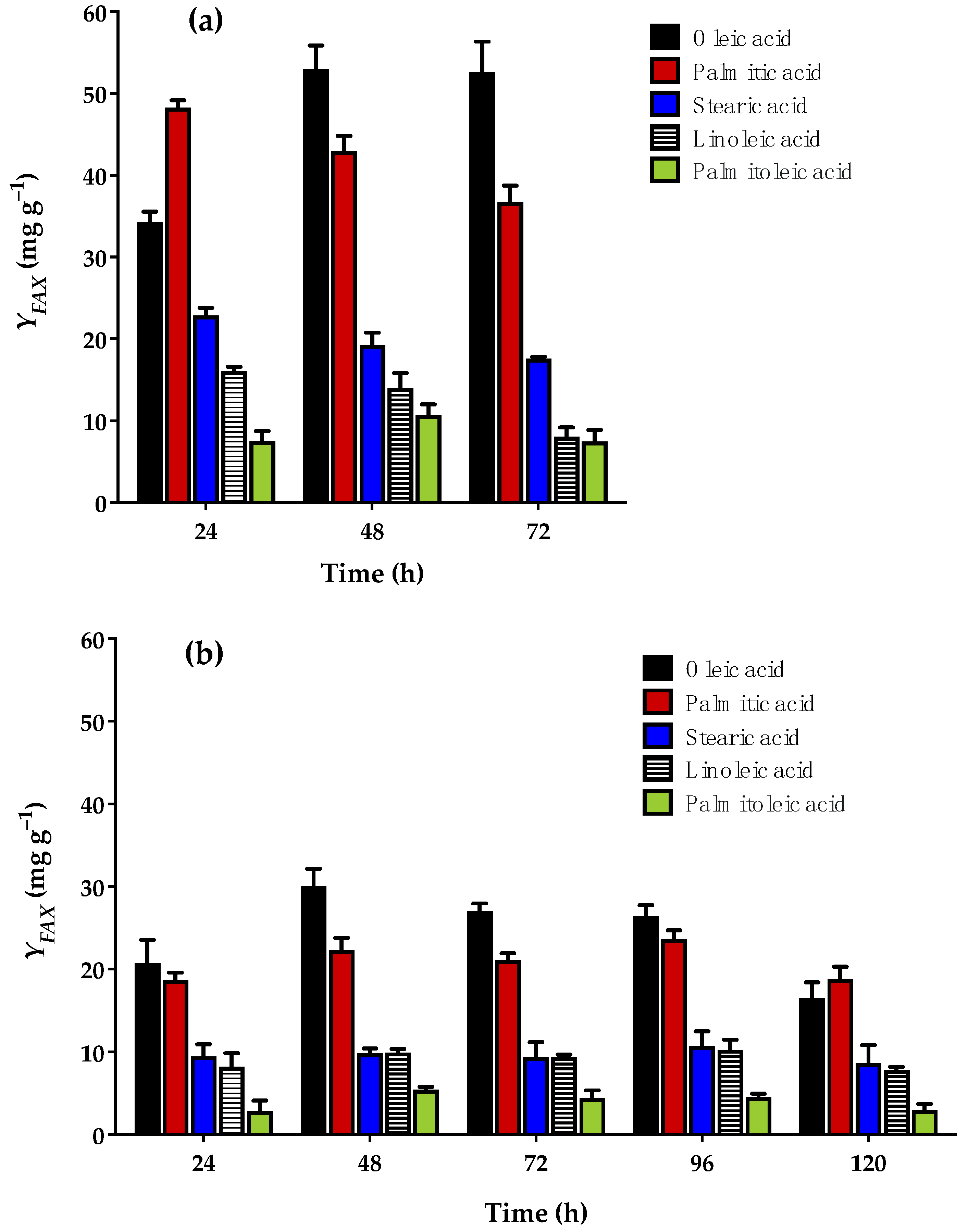
3.4. Fatty Acid Profile
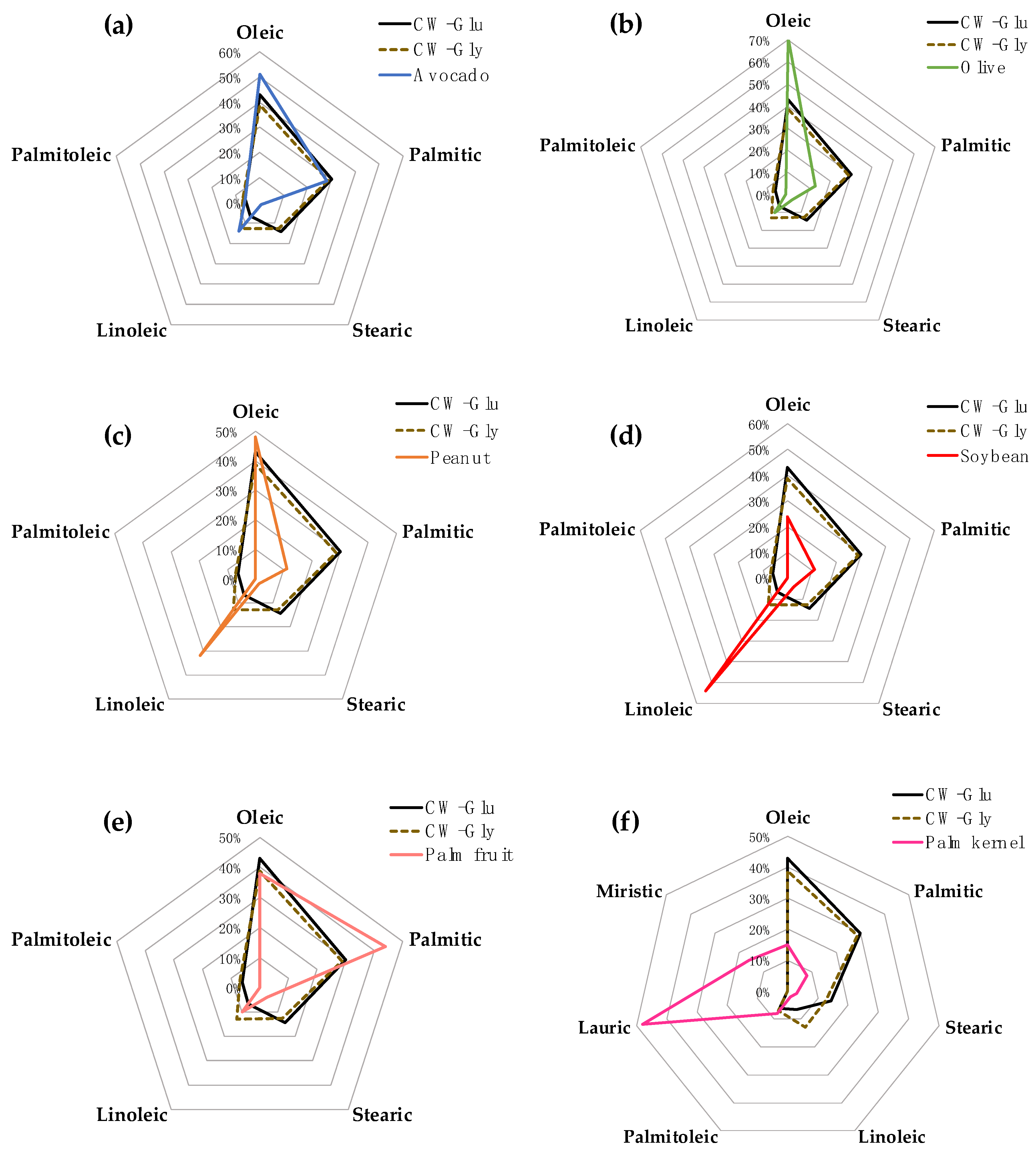
3.5. Comparison of the Composition of the Fatty Acids between CW and Some Vegetable Oils
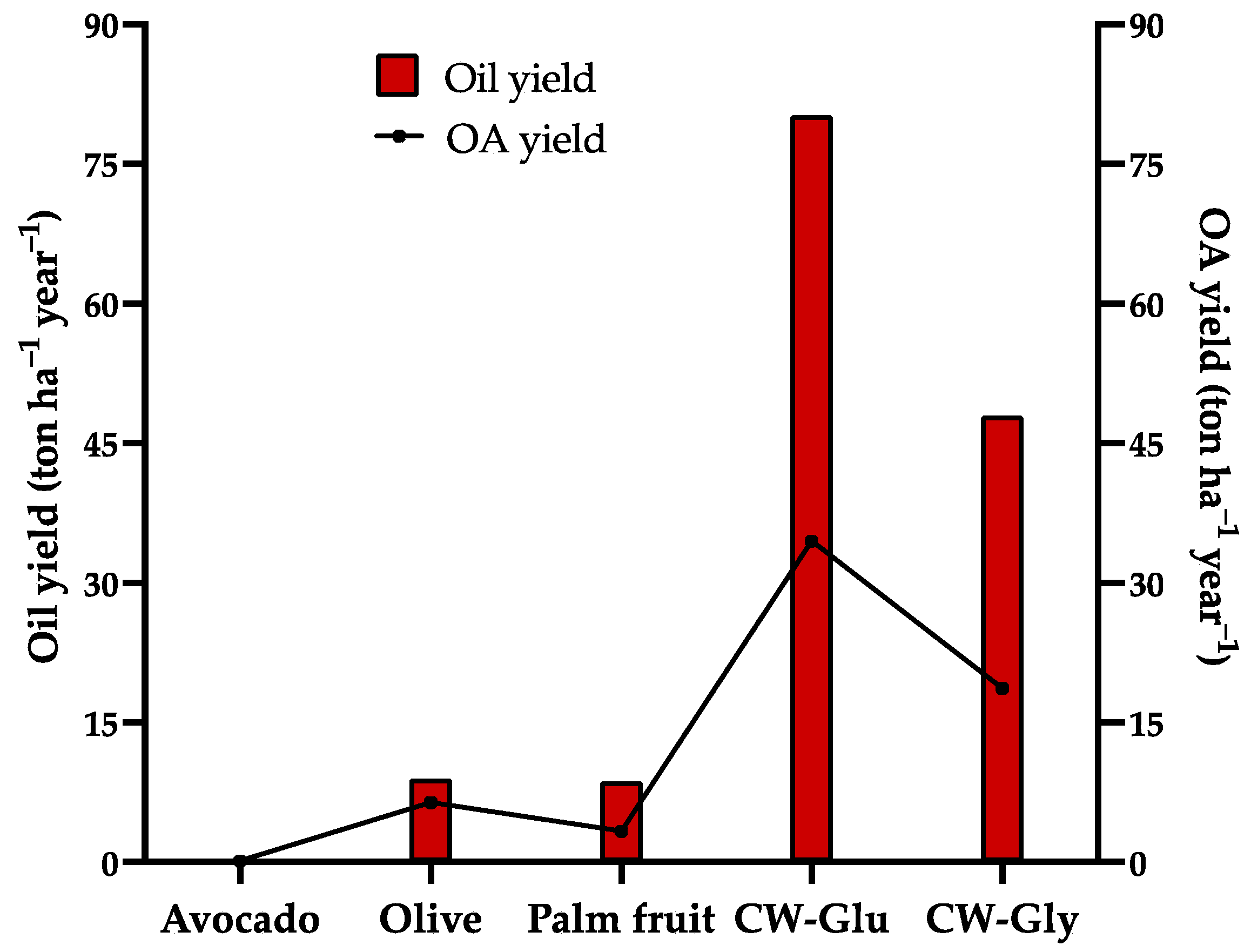
3.6. Plausible Uses of the Oil in CW Based on Its Similarity to Some Vegetable Oils
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghazani, S.M.; Marangoni, A.G. Microbial Lipids for Foods. Trends Food Sci. Technol. 2022, 119, 593–607. [Google Scholar] [CrossRef]
- Orsavova, J.; Misurcova, L.; Vavra Ambrozova, J.; Vicha, R.; Mlcek, J. Fatty Acids Composition of Vegetable Oils and Its Contribution to Dietary Energy Intake and Dependence of Cardiovascular Mortality on Dietary Intake of Fatty Acids. Int. J. Mol. Sci. 2015, 16, 12871–12890. [Google Scholar] [CrossRef] [PubMed]
- Sales-Campos, H.; Reis de Souza, P.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An Overview of the Modulatory Effects of Oleic Acid in Health and Disease. Mini Rev. Med. Chem. 2013, 13, 201–210. [Google Scholar] [PubMed]
- Abdullah, M.M.H.; Jew, S.; Jones, P.J.H. Health Benefits and Evaluation of Healthcare Cost Savings If Oils Rich in Monounsaturated Fatty Acids Were Substituted for Conventional Dietary Oils in the United States. Nutr. Rev. 2017, 75, 163–174. [Google Scholar] [CrossRef]
- Flores, M.; Saravia, C.; Vergara, C.E.; Avila, F.; Valdés, H.; Ortiz-Viedma, J. Avocado Oil: Characteristics, Properties, and Applications. Molecules 2019, 24, 2172. [Google Scholar] [CrossRef]
- Giulitti, F.; Petrungaro, S.; Mandatori, S.; Tomaipitinca, L.; de Franchis, V.; D’Amore, A.; Filippini, A.; Gaudio, E.; Ziparo, E.; Giampietri, C. Anti-Tumor Effect of Oleic Acid in Hepatocellular Carcinoma Cell Lines via Autophagy Reduction. Front. Cell Dev. Biol. 2021, 9, 629182. [Google Scholar] [CrossRef]
- Lopez-Huertas, E. Health Effects of Oleic Acid and Long Chain Omega-3 Fatty Acids (EPA and DHA) Enriched Milks. A Review of Intervention Studies. Pharmacol. Res. 2010, 61, 200–207. [Google Scholar] [CrossRef]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vázquez-Carrera, M. Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2018, 29, 178–190. [Google Scholar] [CrossRef]
- Storniolo, C.E.; Martínez-Hovelman, N.; Martínez-Huélamo, M.; Lamuela-Raventos, R.M.; Moreno, J.J. Extra Virgin Olive Oil Minor Compounds Modulate Mitogenic Action of Oleic Acid on Colon Cancer Cell Line. J. Agric. Food Chem. 2019, 67, 11420–11427. [Google Scholar] [CrossRef]
- Tsarouhas, P.; Achillas, C.; Aidonis, D.; Folinas, D.; Maslis, V. Life Cycle Assessment of Olive Oil Production in Greece. J. Clean. Prod. 2015, 93, 75–83. [Google Scholar] [CrossRef]
- Espadas-Aldana, G.; Vialle, C.; Belaud, J.-P.; Vaca-Garcia, C.; Sablayrolles, C. Analysis and Trends for Life Cycle Assessment of Olive Oil Production. Sustain. Prod. Consum. 2019, 19, 216–230. [Google Scholar] [CrossRef]
- Safitri, L.; Hermantoro, H.; Purboseno, S.; Kautsar, V.; Saptomo, S.K.; Kurniawan, A. Water Footprint and Crop Water Usage of Oil Palm (Eleasis guineensis) in Central Kalimantan: Environmental Sustainability Indicators for Different Crop Age and Soil Conditions. Water 2019, 11, 35. [Google Scholar] [CrossRef]
- Beyer, R.; Rademacher, T. Species Richness and Carbon Footprints of Vegetable Oils: Can High Yields Outweigh Palm Oil’s Environmental Impact? Sustainability 2021, 13, 1813. [Google Scholar] [CrossRef]
- Solarte-Toro, J.C.; Ortiz-Sanchez, M.; Cardona Alzate, C.A. Environmental Life Cycle Assessment (E-LCA) and Social Impact Assessment (SIA) of Small-Scale Biorefineries Implemented in Rural Zones: The Avocado (Persea americana var. americana) Case in Colombia. Environ. Sci. Pollut. Res. 2022, 30, 8790–8808. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The History, State of the Art and Future Prospects for Oleaginous Yeast Research. Microb. Cell Fact. 2021, 20, 221. [Google Scholar] [CrossRef]
- Schulze, I.; Hansen, S.; Großhans, S.; Rudszuck, T.; Ochsenreither, K.; Syldatk, C.; Neumann, A. Characterization of Newly Isolated Oleaginous Yeasts—Cryptococcus podzolicus, Trichosporon porosum and Pichia segobiensis. AMB Express 2014, 4, 24. [Google Scholar] [CrossRef]
- Jiang, W.; Li, C.; Li, Y.; Peng, H. Metabolic Engineering Strategies for Improved Lipid Production and Cellular Physiological Responses in Yeast Saccharomyces cerevisiae. J. Fungi 2022, 8, 427. [Google Scholar] [CrossRef]
- Naranjo, L.; Pernía, B.; Inojosa, Y.; Rojas, D.; D’Anna, L.S.; González, M.; Sisto, Á.D. First Evidence of Fungal Strains Isolated and Identified from Naphtha Storage Tanks and Transporting Pipelines in Venezuelan Oil Facilities. Adv. Microbiol. 2015, 5, 143–154. [Google Scholar] [CrossRef]
- Limtong, S.; Kaewwichian, R.; Jindamorakot, S.; Yongmanitchai, W.; Nakase, T. Candida wangnamkhiaoensis Sp. Nov., an Anamorphic Yeast Species in the Hyphopichia Clade Isolated in Thailand. Antonie Leeuwenhoek 2012, 102, 23–28. [Google Scholar] [CrossRef]
- Hernández-Montañez, Z.; Juárez-Montiel, M.; Velázquez-Ávila, M.; Cristiani-Urbina, E.; Hernández-Rodríguez, C.; Villa-Tanaca, L.; Chávez-Camarillo, G. Production and Characterization of Extracellular α-Amylase Produced by Wickerhamia sp. X-Fep. Appl. Biochem. Biotechnol. 2012, 167, 2117–2129. [Google Scholar] [CrossRef]
- Chávez-Camarillo, G.M.; Santiago-Flores, U.M.; Mena-Vivanco, A.; Morales-Barrera, L.; Cortés-Acosta, E.; Cristiani-Urbina, E. Transient Responses of Wickerhamia sp. Yeast Continuous Cultures to Qualitative Changes in Carbon Source Supply: Induction and Catabolite Repression of α-Amylase Synthesis. Ann Microbiol 2018, 68, 625–635. [Google Scholar] [CrossRef]
- Chávez-Camarillo, G.M.; Lopez-Nuñez, P.V.; Jiménez-Nava, R.A.; Aranda-García, E.; Cristiani-Urbina, E. Production of Extracellular α-Amylase by Single-Stage Steady-State Continuous Cultures of Candida wangnamkhiaoensis in an Airlift Bioreactor. PLoS ONE 2022, 17, e0264734. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol Metabolism and Transport in Yeast and Fungi: Established Knowledge and Ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Rapti, A.; Papafotis, N.; Koutinas, A.A.; Papanikolaou, S. Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain. Molecules 2019, 24, 222. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.B.; Freitas, A.V.; Leitão, R.C.; Pinto, G.A.S.; Santaella, S.T. Anaerobic Digestion of Crude Glycerol: A Review. Environ. Technol. Rev. 2012, 1, 81–92. [Google Scholar] [CrossRef]
- Worthington Worthington Enzyme Handbook: Enzymes, Enzymes Reagents and Related Biochemicals; Biochemical Corporation: Princeton, NJ, USA, 1972.
- Bondioli, P.; Della Bella, L. An Alternative Spectrophotometric Method for the Determination of Free Glycerol in Biodiesel. Eur. J. Lipid Sci. Technol. 2005, 107, 153–157. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A Direct Method for Fatty Acid Methyl Ester Synthesis: Application to Wet Meat Tissues, Oils, and Feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef]
- Kokoreva, A.S.; Isakova, E.P.; Tereshina, V.M.; Klein, O.I.; Gessler, N.N.; Deryabina, Y.I. The Effect of Different Substrates on the Morphological Features and Polyols Production of Endomyces magnusii Yeast during Long-Lasting Cultivation. Microorganisms 2022, 10, 1709. [Google Scholar] [CrossRef]
- Gancedo, J.M. Control of Pseudohyphae Formation in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 107–123. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Sentandreu, R. Different Effectors of Dimorphism in Yarrowia lipolytica. Arch Microbiol 2002, 178, 477–483. [Google Scholar] [CrossRef]
- Merico, A.; Ragni, E.; Galafassi, S.; Popolo, L.; Compagno, C. Generation of an Evolved Saccharomyces cerevisiae Strain with a High Freeze Tolerance and an Improved Ability to Grow on Glycerol. J. Ind. Microbiol. Biotechnol. 2011, 38, 1037–1044. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Makri, A.; Belka, A.; Bellou, S.; Mavrou, M.; Mastoridou, M.; Mystrioti, P.; Onjaro, G.; Aggelis, G.; Papanikolaou, S. Biotechnological Conversions of Biodiesel Derived Waste Glycerol by Yeast and Fungal Species. Energy 2011, 36, 1097–1108. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipid Production by Yarrowia lipolytica Growing on Industrial Glycerol in a Single-Stage Continuous Culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef]
- Angerbauer, C.; Siebenhofer, M.; Mittelbach, M.; Guebitz, G.M. Conversion of Sewage Sludge into Lipids by Lipomyces starkeyi for Biodiesel Production. Bioresour. Technol. 2008, 99, 3051–3056. [Google Scholar] [CrossRef]
- Yang, X.; Jin, G.; Gong, Z.; Shen, H.; Bai, F.; Zhao, Z.K. Recycling Biodiesel-Derived Glycerol by the Oleaginous Yeast Rhodosporidium toruloides Y4 through the Two-Stage Lipid Production Process. Biochem. Eng. J. 2014, 91, 86–91. [Google Scholar] [CrossRef]
- Ramírez-Castrillón, M.; Jaramillo-Garcia, V.P.; Rosa, P.D.; Landell, M.F.; Vu, D.; Fabricio, M.F.; Ayub, M.A.Z.; Robert, V.; Henriques, J.A.P.; Valente, P. The Oleaginous Yeast Meyerozyma guilliermondii BI281A as a New Potential Biodiesel Feedstock: Selection and Lipid Production Optimization. Front. Microbiol. 2017, 8, 1776. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chang, K.-S.; Hsu, C.-L.; Chuang, L.-T.; Chen, C.-Y.; Huang, F.-Y.; Jang, H.-D. A Comparative Study on Batch and Fed-Batch Cultures of Oleaginous Yeast Cryptococcus Sp. in Glucose-Based Media and Corncob Hydrolysate for Microbial Oil Production. Fuel 2013, 105, 711–717. [Google Scholar] [CrossRef]
- Ghanavati, H.; Nahvi, I.; Roghanian, R. Monitoring Growth and Lipid Production of New Isolated Oleaginous Yeast Cryptococcus aerius UIMC65 on Glucose and Xylose Cultures. Biotechnol. Bioprocess. Eng. 2014, 19, 468–477. [Google Scholar] [CrossRef]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic Activities of Biotechnological Interest in Yarrowia lipolytica Grown on Glycerol in Repeated Batch Cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar] [CrossRef]
- Jiru, T.M.; Steyn, L.; Pohl, C.; Abate, D. Production of Single Cell Oil from Cane Molasses by Rhodotorula kratochvilovae (Syn, Rhodosporidium kratochvilovae) SY89 as a Biodiesel Feedstock. Chem. Cent. J. 2018, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Carsanba, E.; Papanikolaou, S.; Fickers, P.; Erten, H. Lipids by Yarrowia lipolytica Strains Cultivated on Glucose in Batch Cultures. Microorganisms 2020, 8, 1054. [Google Scholar] [CrossRef] [PubMed]
- Castanha, R.F.; Mariano, A.P.; de Morais, L.A.S.; Scramin, S.; Monteiro, R.T.R. Optimization of Lipids Production by Cryptococcus laurentii 11 Using Cheese Whey with Molasses. Braz. J. Microbiol. 2014, 45, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Gientka, I.; Kieliszek, M.; Jermacz, K.; Błażejak, S. Identification and Characterization of Oleaginous Yeast Isolated from Kefir and Its Ability to Accumulate Intracellular Fats in Deproteinated Potato Wastewater with Different Carbon Sources. Biomed Res. Int. 2017, 2017, 6061042. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jin, G.; Gong, Z.; Shen, H.; Song, Y.; Bai, F.; Zhao, Z.K. Simultaneous Utilization of Glucose and Mannose from Spent Yeast Cell Mass for Lipid Production by Lipomyces starkeyi. Bioresour. Technol. 2014, 158, 383–387. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chatzifragkou, A.; Fakas, S.; Galiotou-Panayotou, M.; Komaitis, M.; Nicaud, J.-M.; Aggelis, G. Biosynthesis of Lipids and Organic Acids by Yarrowia lipolytica Strains Cultivated on Glucose. Eur. J. Lipid Sci. Technol. 2009, 111, 1221–1232. [Google Scholar] [CrossRef]
- Sankh, S.; Thiru, M.; Saran, S.; Rangaswamy, V. Biodiesel Production from a Newly Isolated Pichia kudriavzevii Strain. Fuel 2013, 106, 690–696. [Google Scholar] [CrossRef]
- Sandoval, N.R.; Papoutsakis, E.T. Engineering Membrane and Cell-Wall Programs for Tolerance to Toxic Chemicals: Beyond Solo Genes. Curr. Opin. Microbiol. 2016, 33, 56–66. [Google Scholar] [CrossRef]
- Doignon, F.; Laquel, P.; Testet, E.; Tuphile, K.; Fouillen, L.; Bessoule, J.-J. Requirement of Phosphoinositides Containing Stearic Acid to Control Cell Polarity. Cell. Mol. Biol. 2016, 36, 765–780. [Google Scholar] [CrossRef]
- Besada-Lombana, P.B.; Fernandez-Moya, R.; Fenster, J.; Da Silva, N.A. Engineering Saccharomyces cerevisiae Fatty Acid Composition for Increased Tolerance to Octanoic Acid. Biotechnol. Bioeng. 2017, 114, 1531–1538. [Google Scholar] [CrossRef]
- Rodríguez-Vargas, S.; Sánchez-García, A.; Martínez-Rivas, J.M.; Prieto, J.A.; Randez-Gil, F. Fluidization of Membrane Lipids Enhances the Tolerance of Saccharomyces cerevisiae to Freezing and Salt Stress. Appl. Environ. Microbiol. 2007, 73, 110–116. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. High-Temperature Ethanol Fermentation from Pineapple Waste Hydrolysate and Gene Expression Analysis of Thermotolerant Yeast Saccharomyces cerevisiae. Sci. Rep. 2022, 12, 13965. [Google Scholar] [CrossRef]
- You, K.M.; Rosenfield, C.-L.; Knipple, D.C. Ethanol Tolerance in the Yeast Saccharomyces cerevisiae Is Dependent on Cellular Oleic Acid Content. App. Environ. Microbiol. 2003, 69, 1499–1503. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kim, N.-R.; Choi, W. Total Fatty Acid Content of the Plasma Membrane of Saccharomyces cerevisiae Is More Responsible for Ethanol Tolerance than the Degree of Unsaturation. Biotechnol. Lett. 2011, 33, 509–515. [Google Scholar] [CrossRef]
- Patti, A.; Lecocq, H.; Serghei, A.; Acierno, D.; Cassagnau, P. The Universal Usefulness of Stearic Acid as Surface Modifier: Applications to the Polymer Formulations and Composite Processing. J. Ind. Eng. Chem. 2021, 96, 1–33. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Aggelis, G.; Marc, I. Kinetic Profile of the Cellular Lipid Composition in an Oleaginous Yarrowia lipolytica Capable of Producing a Cocoa-Butter Substitute from Industrial Fats. Antonie Van Leeuwenhoek 2001, 80, 215–224. [Google Scholar] [CrossRef]
- Loften, J.R.; Linn, J.G.; Drackley, J.K.; Jenkins, T.C.; Soderholm, C.G.; Kertz, A.F. Invited Review: Palmitic and Stearic Acid Metabolism in Lactating Dairy Cows. J. Dairy Sci. 2014, 97, 4661–4674. [Google Scholar] [CrossRef]
- Sun, P.; Xia, B.; Ni, Z.-J.; Wang, Y.; Elam, E.; Thakur, K.; Ma, Y.-L.; Wei, Z.-J. Characterization of Functional Chocolate Formulated Using Oleogels Derived from β-Sitosterol with γ-Oryzanol/Lecithin/Stearic Acid. Food Chem. 2021, 360, 130017. [Google Scholar] [CrossRef]
- Zhang, L.; Lim, E.Y.; Loh, K.-C.; Dai, Y.; Tong, Y.W. Two-Stage Fermentation of Lipomyces starkeyi for Production of Microbial Lipids and Biodiesel. Microorganisms 2021, 9, 1724. [Google Scholar] [CrossRef]
- Vasconcelos, B.; Teixeira, J.C.; Dragone, G.; Teixeira, J.A. Oleaginous Yeasts for Sustainable Lipid Production—From Biodiesel to Surf Boards, a Wide Range of “Green” Applications. Appl. Microbiol. Biotechnol. 2019, 103, 3651–3667. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous Yeasts for Biodiesel: Current and Future Trends in Biology and Production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef] [PubMed]
- Ranade, S.; Thiagarajan, P. A Review on Persea americana Mill. (Avocado)- Its Fruit and Oil. Int. J. Pharmtech Res. 2015, 8, 72–77. [Google Scholar]
- Thevenieau, F.; Nicaud, J.-M. Microorganisms as Sources of Oils. OCL 2013, 20, D603. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M. Avocado Oil: Production and Market Demand, Bioactive Components, Implications in Health, and Tendencies and Potential Uses. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4120–4158. [Google Scholar] [CrossRef]
- Mba, O.I.; Dumont, M.-J.; Ngadi, M. Palm Oil: Processing, Characterization and Utilization in the Food Industry—A Review. Food Biosci. 2015, 10, 26–41. [Google Scholar] [CrossRef]
- Datos Abiertos|Servicio de Información Agroalimentaria y Pesquera|Gob.Mx. Available online: http://infosiap.siap.gob.mx/gobmx/datosAbiertos_a.php (accessed on 9 March 2023).
- Vossen, P. Growing Olives for Oil. In Handbook of Olive Oil: Analysis and Properties; Aparicio, R., Harwood, J., Eds.; Springer US: Boston, MA, USA, 2013; pp. 19–56. ISBN 978-1-4614-7777-8. [Google Scholar]
- Global Olive Production 2012–2020. Available online: https://www.statista.com/statistics/937274/olive-production-global/ (accessed on 9 March 2023).
- Murphy, D.J.; Goggin, K.; Paterson, R.R.M. Oil Palm in the 2020s and beyond: Challenges and Solutions. CABI Agric. Biosci. 2021, 2, 39. [Google Scholar] [CrossRef]
- Pande, G.; Akoh, C.C.; Lai, O.-M. 19—Food Uses of Palm Oil and Its Components. In Palm Oil; Lai, O.-M., Tan, C.-P., Akoh, C.C., Eds.; AOCS Press: Urbana, IL, USA, 2012; pp. 561–586. ISBN 978-0-9818936-9-3. [Google Scholar]
- Vijay, V.; Pimm, S.L.; Jenkins, C.N.; Smith, S.J. The Impacts of Oil Palm on Recent Deforestation and Biodiversity Loss. PLoS ONE 2016, 11, e0159668. [Google Scholar] [CrossRef]
- Kareem, M.A.; Raheem, A.A.; Oriola, K.O.; Abdulwahab, R. A Review on Application of Oil Palm Shell as Aggregate in Concrete—Towards Realising a Pollution-Free Environment and Sustainable Concrete. Environ. Chall. 2022, 8, 100531. [Google Scholar] [CrossRef]
- UNdata|Record View|Avocados. Available online: http://data.un.org/Data.aspx?d=FAO&f=itemCode%3A572 (accessed on 9 March 2023).
- Peraza-Magallanes, A.Y.; Pereyra-Camacho, M.A.; Sandoval-Castro, E.; Medina-Godoy, S.; Valdez-Morales, M.; Clegg, M.T.; Calderón-Vázquez, C.L. Exploring Genetic Variation, Oil and α-Tocopherol Content in Avocado (Persea americana) from Northwestern Mexico. Genet. Resour. Crop Evol. 2017, 64, 443–449. [Google Scholar] [CrossRef]
- Urquiza, L.S.P.; Rebollar, S.R.; Juárez, N.C.; Martínez, J.H.; Tenorio, G.G. Análisis de viabilidad económica para la producción comercial de aguacate Hass. Rev. Mex. Agronegocios 2015, 36, 1325–1338. [Google Scholar]
- Silber, A.; Naor, A.; Cohen, H.; Bar-Noy, Y.; Yechieli, N.; Levi, M.; Noy, M.; Peres, M.; Duari, D.; Narkis, K.; et al. Avocado Fertilization: Matching the Periodic Demand for Nutrients. Sci. Hortic. 2018, 241, 231–240. [Google Scholar] [CrossRef]
- UNdata|Record View|Olives (Including Preserved). Available online: http://data.un.org/Data.aspx?q=olive&d=FAO&f=itemCode%3a2563 (accessed on 9 March 2023).
- Kiritsakis, A.; Turkan, K.M.; Kiritsakis, K. Olive Oil. In Bailey’s Industrial Oil and Fat Products; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 1–38. ISBN 978-0-471-67849-6. [Google Scholar]
- El Riachy, M.; Hamade, A.; Ayoub, R.; Dandachi, F.; Chalak, L. Oil Content, Fatty Acid and Phenolic Profiles of Some Olive Varieties Growing in Lebanon. Front. Nutr. 2019, 6, 94. [Google Scholar] [CrossRef]
- The World of Olive Oil. Available online: https://www.internationaloliveoil.org/the-world-of-olive-oil/ (accessed on 9 March 2023).
- Lentza-Rizos, C.; Avramides, E.J. Pesticide Residues in Olive Oil. In Reviews of Environmental Contamination and Toxicology: Continuation of Residue Reviews; Ware, G.W., Gunther, F.A., Eds.; Springer: New York, NY, USA, 1995; pp. 111–134. ISBN 978-1-4612-2530-0. [Google Scholar]
- UNdata|Record View|Oil Palm Fruit. Available online: http://data.un.org/Data.aspx?q=oil+palm&d=FAO&f=itemCode%3a254 (accessed on 9 March 2023).
- Tranbarger, T.J.; Dussert, S.; Joët, T.; Argout, X.; Summo, M.; Champion, A.; Cros, D.; Omore, A.; Nouy, B.; Morcillo, F. Regulatory Mechanisms Underlying Oil Palm Fruit Mesocarp Maturation, Ripening, and Functional Specialization in Lipid and Carotenoid Metabolism. Plant Physiol. 2011, 156, 564–584. [Google Scholar] [CrossRef]
- Global Production Volume Palm Oil 20222/23. Available online: https://www.statista.com/statistics/613471/palm-oil-production-volume-worldwide/ (accessed on 8 March 2023).
- Watson-Hernández, F.; Gómez-Calderón, N.; da Silva, R.P. Oil Palm Yield Estimation Based on Vegetation and Humidity Indices Generated from Satellite Images and Machine Learning Techniques. AgriEngineering 2022, 4, 279–291. [Google Scholar] [CrossRef]
- Papong, S.; Chom-In, T.; Noksa-nga, S.; Malakul, P. Life Cycle Energy Efficiency and Potentials of Biodiesel Production from Palm Oil in Thailand. Energy Policy 2010, 38, 226–233. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Formula | Nomenclature |
|---|---|---|
| Substrate consumption efficiency, Ef (%) | St: Residual concentration of the substrate at time t (h) (grams of substrate L−1) S0: Substrate concentration at the initial time t = 0 h (grams of substrate L−1) Pt: Lipid concentration at time t (h) (grams of lipids L−1) P0: Lipid concentration at the initial time t = 0 h (grams of lipids L−1) Xt: Biomass concentration at time t (h) (grams of biomass L−1) X0: Biomass concentration at the initial time t = 0 h (grams of biomass L−1) FAt: Fatty acid (FA) content at time t (h) (milligrams of FAs L−1) FA0: Fatty acid (FA) content at the initial time t = 0 h (milligrams of FAs L−1) | |
| Lipid yield based on biomass, YPX (%) | ||
| Lipid yield based on substrate, YPS (g g−1) | ||
| Fatty acid yield based on biomass, YFAX (mg g−1) | ||
| Volumetric biomass productivity, Rx (mg L−1 h−1) | ||
| Volumetric lipid productivity, RP (mg L−1 h−1) | ||
| Specific lipid productivity, Rpx (mg g−1 h−1) |
| Time (h) | Ef (%) | X (g L−1) | YPX (%) | Lipids (g L−1) | YPS (g g−1) | RX (mg L−1 h−1) | RP (mg L−1 h−1) | RPX (mg g−1 h−1) |
|---|---|---|---|---|---|---|---|---|
| 24 | 100 | 2.25 ± 0.05 a | 16.56 a | 0.37 ± 0.04 a | 0.03 ± 0.004 a | 93.75 ± 2.09 a | 15.52 ± 1.67 a | 6.90 ± 0.74 a |
| 48 | 100 | 2.68 ± 0.05 b | 18.26 a | 0.48 ± 0.02 a | 0.05 ± 0.002 a | 55.73 ± 0.99 b | 10.01 ± 0.42 b | 3.81 ± 0.16 b |
| 72 | 100 | 3.35 ± 0.07 c | 25.61 b | 0.87 ± 0.02 b | 0.09 ± 0.001 b | 46.53 ± 0.89 c | 12.09 ± 0.03 ab | 3.56 ± 0.01 b |
| Time (h) | Ef (%) | X (g L−1) | YPX (%) | Lipids (g L−1) | YPS (g g−1) | RX (mg L−1 h−1) | RP (mg L−1 h−1) | RPX (mg g−1 h−1) |
|---|---|---|---|---|---|---|---|---|
| 24 | 11.79 | 0.58 ± 0.08 a | 15.35 a | 0.10 ± 0.01 a | 0.08 ± 0.02 a | 23.96 ± 3.31 a | 4.12 ± 0.21 a | 6.39 ± 0.45 a |
| 48 | 34.41 | 1.53 ± 0.11 b | 21.06 b | 0.35 ± 0.02 b | 0.09 ± 0.007 a | 31.77 ± 2.38 b | 7.22 ± 0.49 b | 4.38 ± 0.23 b |
| 72 | 56.97 | 2.51 ± 0.06 c | 23.21 b | 0.58 ± 0.02 c | 0.09 ± 0.004 a | 34.89 ± 0.81 b | 8.09 ± 0.31 b | 3.22 ± 0.13 c |
| 96 | 74.00 | 3.33 ± 0.06 d | 24.64 b | 0.82 ± 0.04 d | 0.10 ± 0.005 a | 34.64 ± 0.67 b | 8.86 ± 0.49 b | 2.67 ± 0.18 cd |
| 120 | 96.35 | 3.63 ± 0.08 d | 24.06 b | 0.88 ± 0.05 d | 0.10 ± 0.007 a | 30.52 ± 0.69 ab | 7.36 ± 0.44 b | 2.00 ± 0.14 d |
| Yeast | Carbon Source | C/N Ratio (mol mol−1) | Initial pH | Temp (°C) | YPX (%) | Incubation Time (h) | Fatty Acid Composition (% w/w) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Myristic (14:0) | Palmitic (16:0) | Palmitoleic (16:1) | Stearic (18:0) | Oleic (18:1) | Linoleic (18:2) | Linolenic (18:3) | Ref. | |||||||
| Candida inconspicua IGII | YPGlc | NR | 5.6 | 28 | 32 | 96 | ND | 16.61 | 2.18 | 12.28 | 48.5 | 18.28 | 2.15 | [45] |
| Candida wangnamkhiaoensis | Glucose | 23.7 | 6.0 | 30 | 25 | 72 | ND | 30 | 6 | 14 | 43 | 7 | ND | This work |
| Cryptococcus aerius UIMC65 | Glucose | 40 | 5.5 | 28 | 77 | 72 | 0.2 | 1.7 | 21.21 | 7.67 | 61.39 | 6.17 | NR | [40] |
| Debaryomyces hansenii 1 | YPGlc | NR | 5.6 | 28 | 46 | 96 | ND | 15.94 | 3.8 | 11.5 | 46.2 | 15.21 | 2.53 | [5] |
| Lipomyces starkeyi AS 2.1560 | Glucose | 1540 * | 6.0 | 30 | 59.3 | 120 | 0.2 | 35.6 | 3.8 | 6.0 | 53.1 | 0.7 | 0.4 | [46] |
| Yarrowia lipolytica JMY 794 | Glucose | 200 | 6.2 | 28 | 9 | 100 | ND | 19.3 | 11.3 | 10.2 | 41.1 | 18.1 | ND | [47] |
| Candida oleophila ATCC 20177 | Pure glycerol | 66 | 6 | 28 | 15.3 | 150 | NR | 12.9 | 2.5 | 6.6 | 65.6 | 11.0 | NR | [34] |
| Candida wangnamkhiaoensis | Pure glycerol | 23.7 | 6.0 | 30 | 24 | 48 | ND | 29 | 7 | 13 | 39 | 13 | ND | This work |
| Meyerozyma guilliermondii BI281A | Pure glycerol | 407 | NR | 28 | 34.97 | 120 | NR | 25.75 | ND | 34.17 | 48.76 | NR | 4.2 | [38] |
| Pichia kudriavzevii MTCC 5493 | Crude glycerol | NR | 5.5 | 28 | 18.6 | 110 | NR | 29.4 | NR | 8.9 | 41.9 | 9.2 | NR | [48] |
| Rhodosporidium toruloides Y4 | Pure glycerol | NR | 5.5 | 30 | 34.8 | 120 | 1.4 | 27.8 | 0.6 | 21.8 | 43.8 | 2.9 | 1.2 | [37] |
| Rhodotorula sp. LFMB22 | Pure glycerol | 66 | 6 | 28 | 22 | 187 | NR | 21.7 | 1.1 | 7.4 | 55.9 | 12.4 | NR | [34] |
| Crop | World Crop Production (Million Metric tons year−1) | Crop Yield (tons ha−1 year−1) | Pulp Content (%) | Oil Contained in the Pulp (%) | World Oil Production (Million Metric tons year−1) | Oil Yield (tons ha−1 year−1) | Productive Time of Trees (Years) | |
|---|---|---|---|---|---|---|---|---|
| To Begin Yield | Maximum Productive | |||||||
| Age | ||||||||
| Avocado (Persea americana Mill.) | 8.06 [74] | 9.6 [67] | 60 [5] | 7–37 [75] | 0.371 [65] | NDA | 3–4 [76] | 30–50 [77] |
| Olive (Olea europaea L.) | 21.33 [78] | 0.5–12 [68] | 65–85 [79] | 30–40 [80] | 3.1 [81] | 9.0 [82] | 5–6 [68] | >100 [68] |
| Oil palm (Elaeis guineensis Jacq.) | 418.4 [83] | 14.56 [83] | 35–75 [84] | 45–70 [66,71,84] | 73.8 [85] | 1.92–15.49 [70,85,86] | 3–6 [87] | 25 [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Rodríguez, A.; Flores-Ortiz, C.M.; Chávez-Camarillo, G.M.; Cristiani-Urbina, E.; Morales-Barrera, L. Potential Capacity of Candida wangnamkhiaoensis to Produce Oleic Acid. Fermentation 2023, 9, 443. https://doi.org/10.3390/fermentation9050443
Pérez-Rodríguez A, Flores-Ortiz CM, Chávez-Camarillo GM, Cristiani-Urbina E, Morales-Barrera L. Potential Capacity of Candida wangnamkhiaoensis to Produce Oleic Acid. Fermentation. 2023; 9(5):443. https://doi.org/10.3390/fermentation9050443
Chicago/Turabian StylePérez-Rodríguez, Alejandro, César Mateo Flores-Ortiz, Griselda Ma. Chávez-Camarillo, Eliseo Cristiani-Urbina, and Liliana Morales-Barrera. 2023. "Potential Capacity of Candida wangnamkhiaoensis to Produce Oleic Acid" Fermentation 9, no. 5: 443. https://doi.org/10.3390/fermentation9050443
APA StylePérez-Rodríguez, A., Flores-Ortiz, C. M., Chávez-Camarillo, G. M., Cristiani-Urbina, E., & Morales-Barrera, L. (2023). Potential Capacity of Candida wangnamkhiaoensis to Produce Oleic Acid. Fermentation, 9(5), 443. https://doi.org/10.3390/fermentation9050443