
Calm Hu Sheep Have a Different Microbiome Profile and Higher Energy Utilization Efficiency Than Nervous Hu Sheep
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection for Temperament
2.2. Animal Feeding Management
2.3. Sample Collection and Processing
2.4. Measurement Indexes and Methods
2.4.1. Determination of Rumen Fermentation Parameters
2.4.2. DNA Extraction and High-Throughput Sequencing
2.5. Statistical and Abundance Analyses
3. Results
3.1. The Effect of Temperament on Growth and Feed Utilization
3.2. Rumen Fermentation Parameters
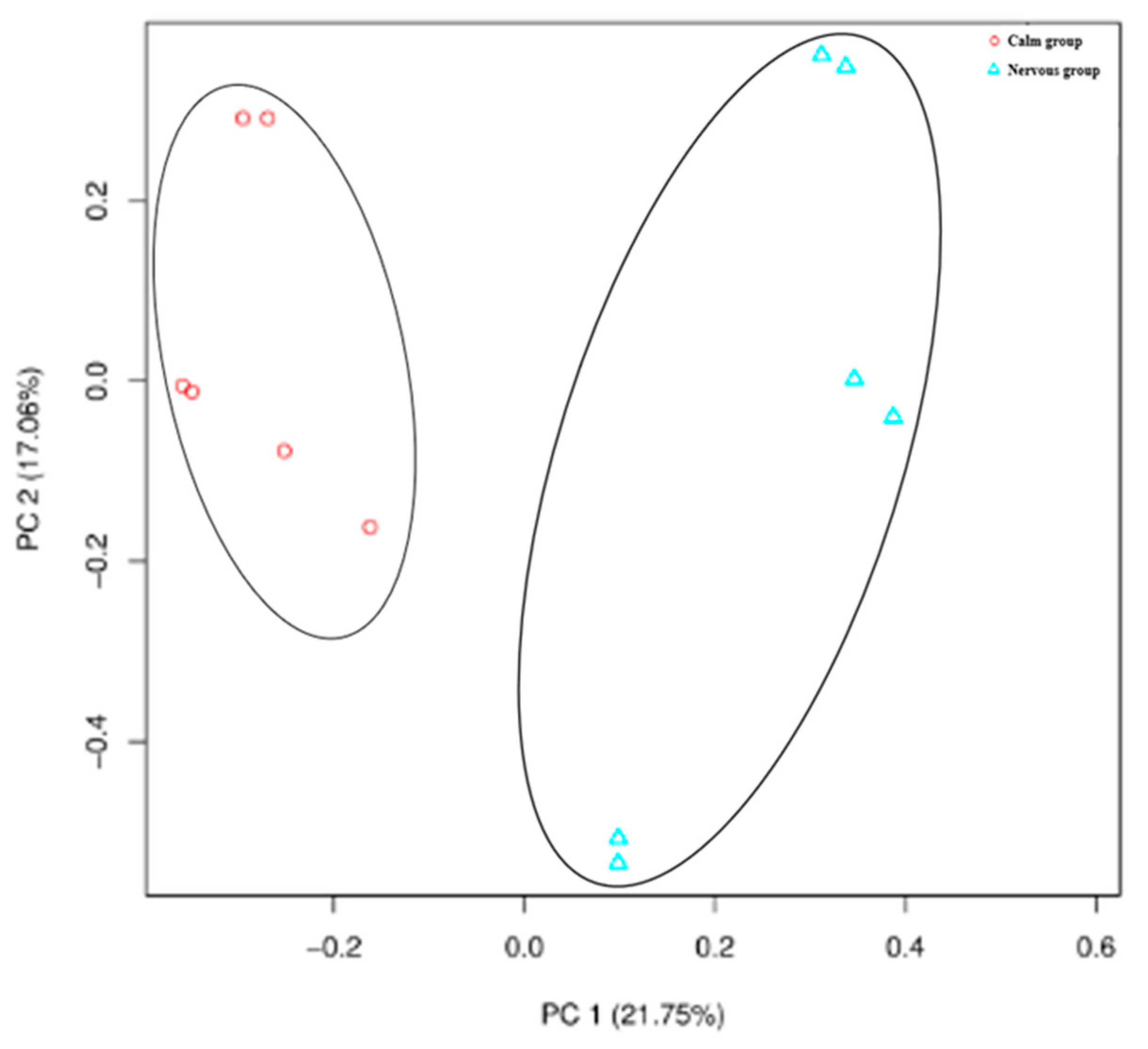
3.3. The Effect of Temperament on Rumen Microbial Structure
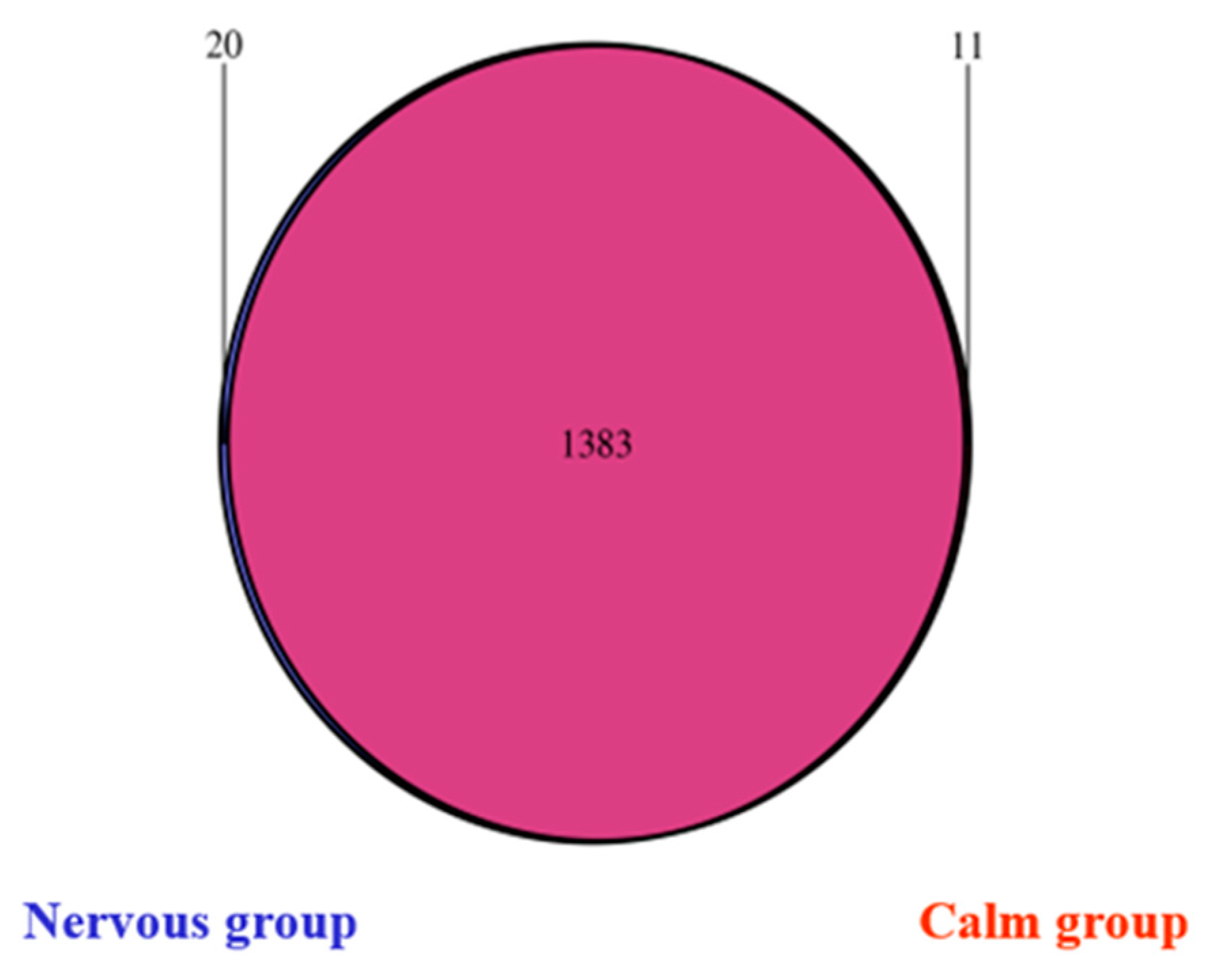
3.3.1. Rumen Bacterial Sequencing OTUs and Alpha Diversities
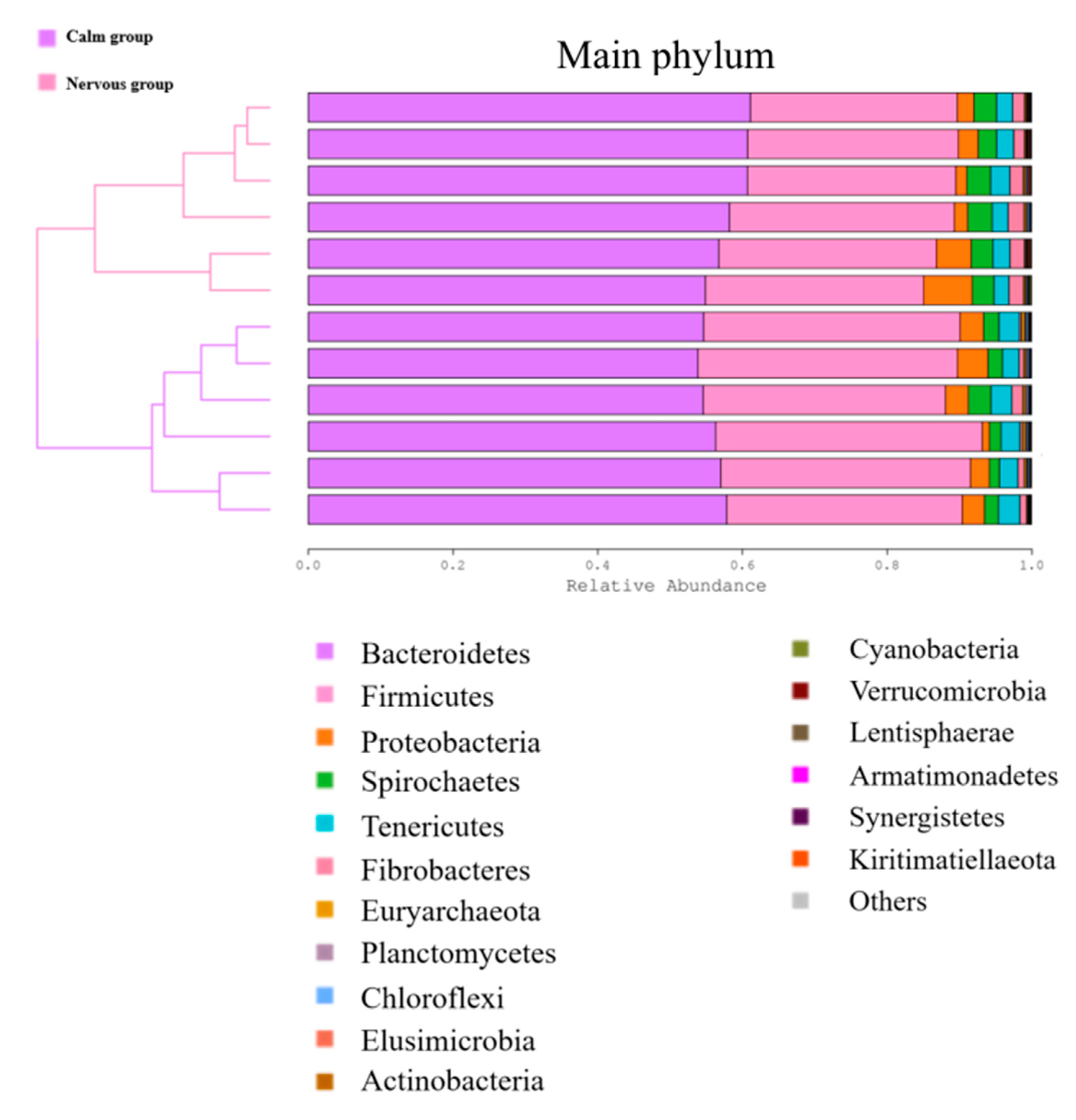
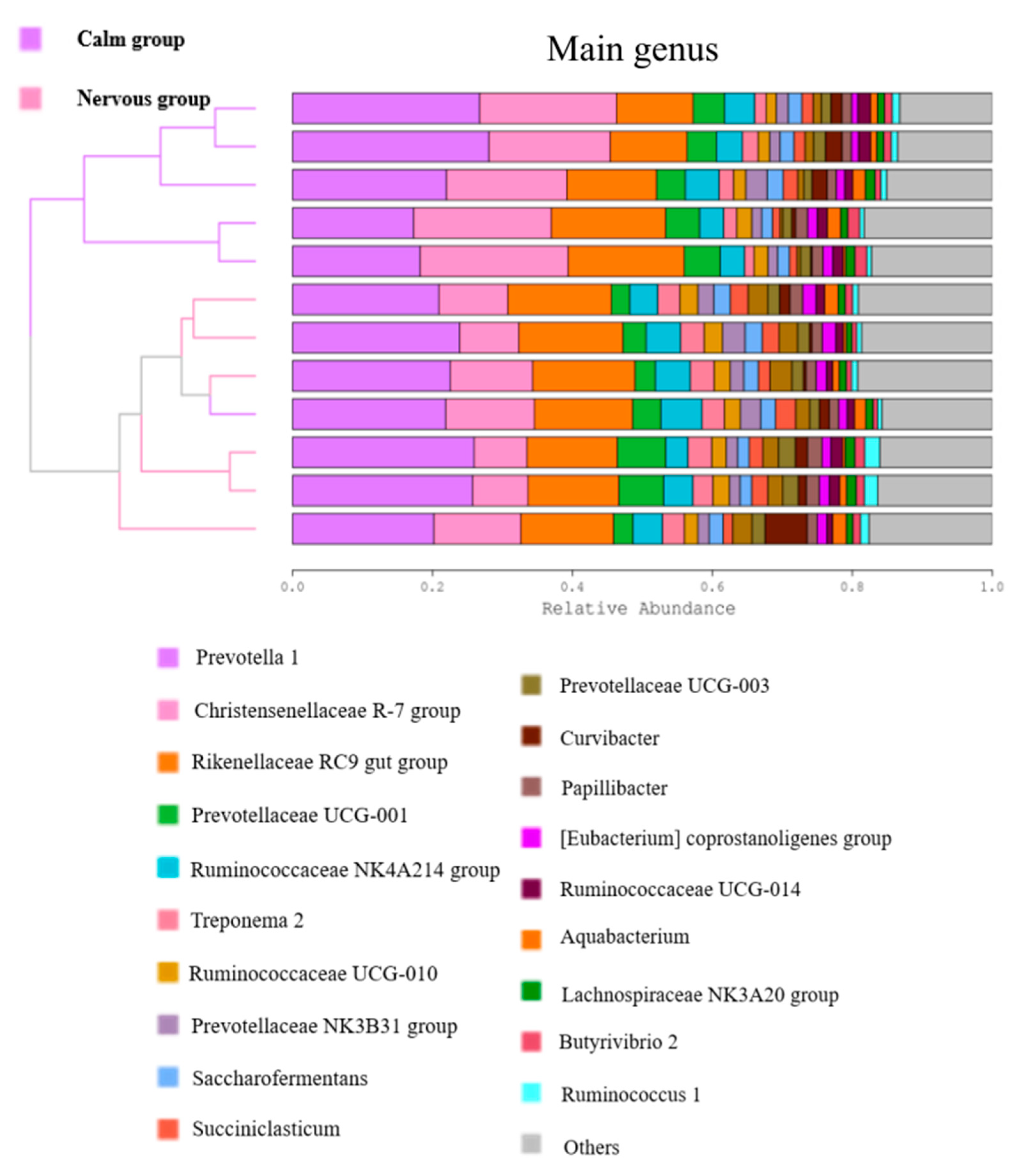
3.3.2. The Effect of Temperament on the Relative Abundance of the Rumen Bacterial Community at the Phylum and Genus Level
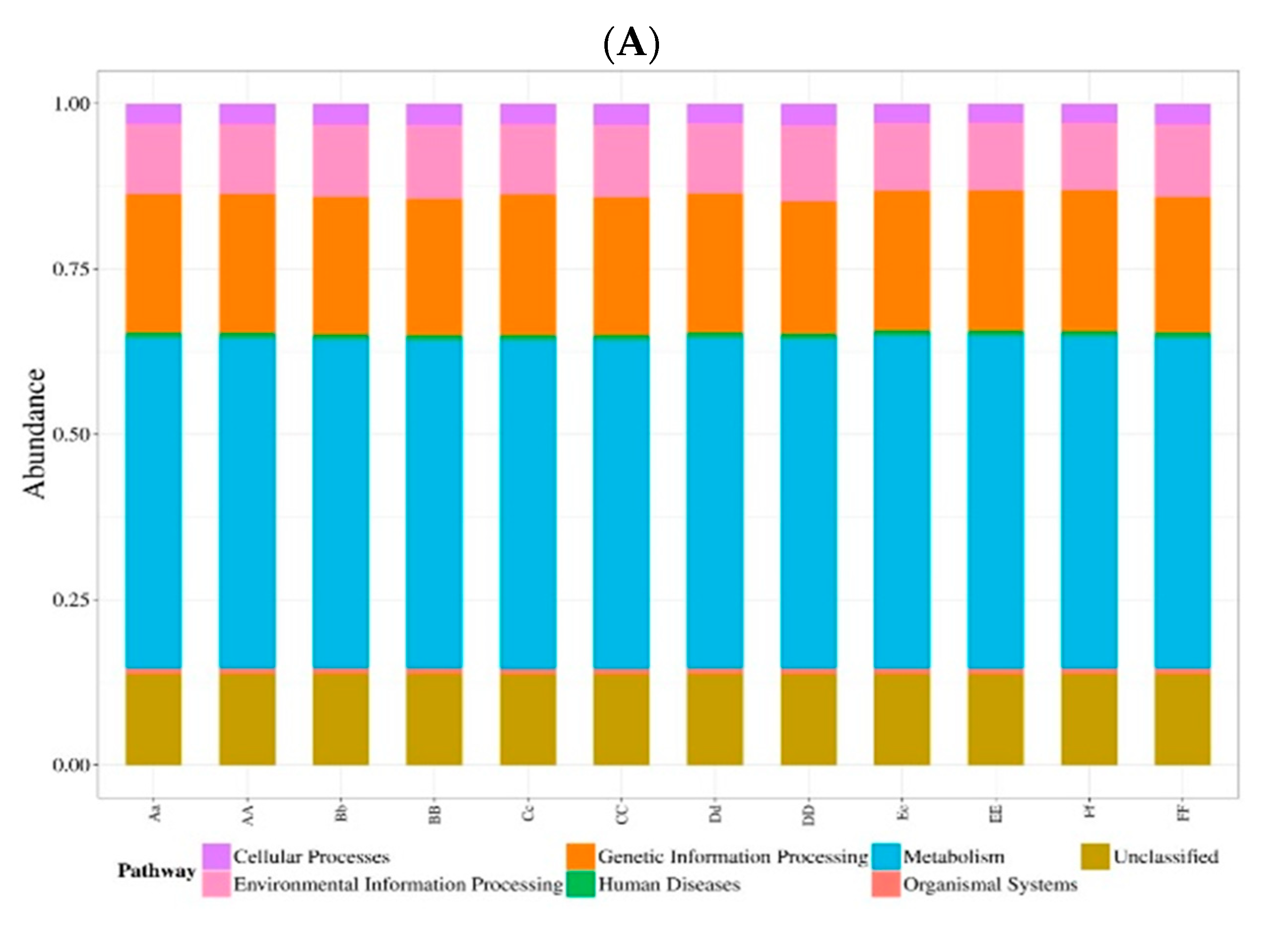
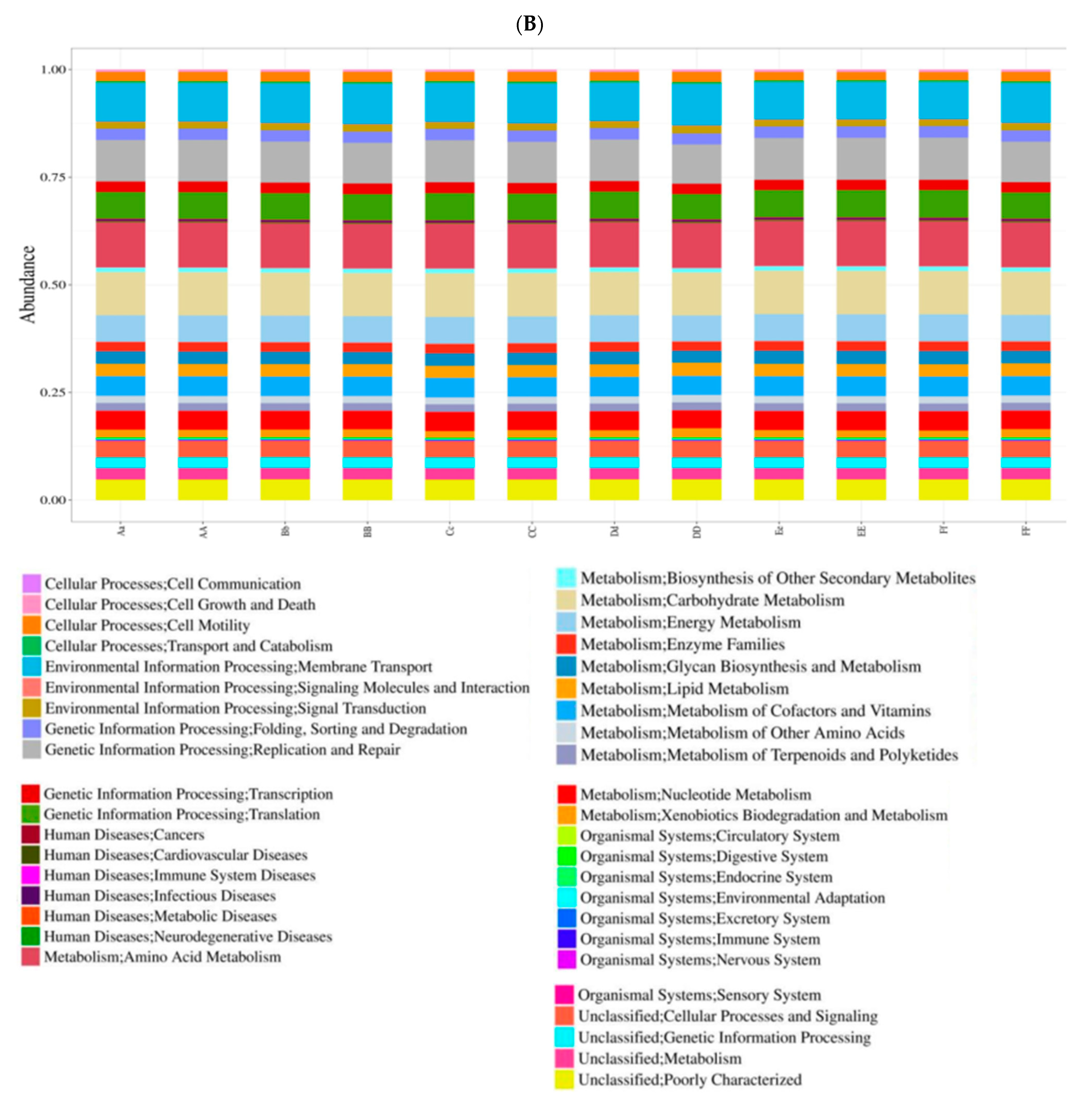
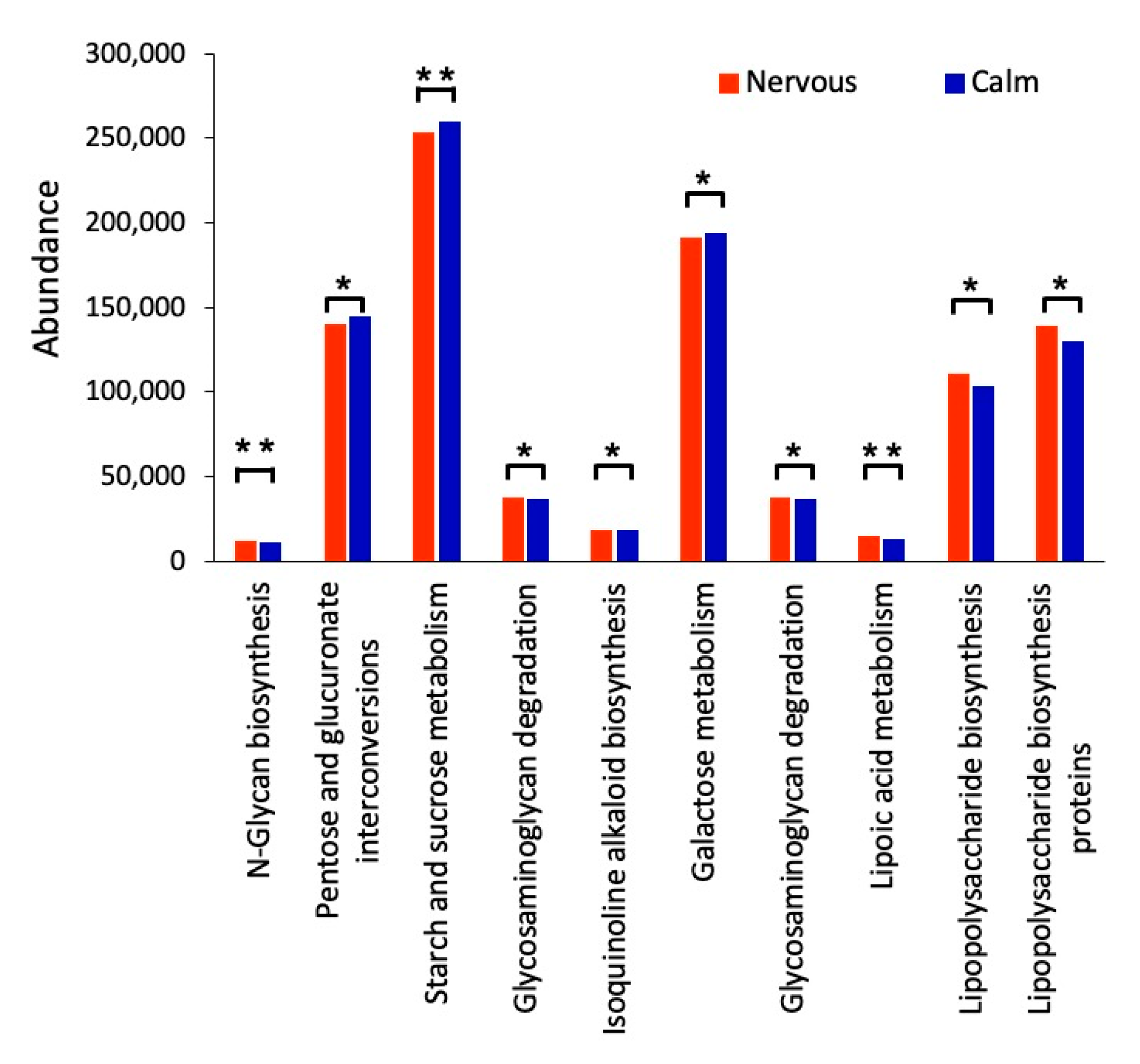
3.4. Prediction of Bacterial Functions via KEGG
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sutherland, M.A.; Rogers, A.R.; Verkerk, G.A. The effect of temperament and responsiveness towards humans on the behavior, physiology and milk production of multi-parous dairy cows in a familiar and novel milking environment. Physiol. Behav. 2012, 107, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.M. Individual differences in temperament of dairy goats and the inhibition of milk ejection. Appl. Anim. Behav. Sci. 1989, 22, 269–282. [Google Scholar] [CrossRef]
- Beausoleil, N.J.; Blache, D.; Stafford, K.J.; Mellor, D.J.; Noble, A.D.L. Exploring the basis of divergent selection for ‘temperament’ in domestic sheep. Appl. Anim. Behav. Sci. 2008, 109, 261–274. [Google Scholar] [CrossRef]
- Burrow, H.M. Measurements of temperament and their relationships with performance traits of beef cattle. Anim. Breed. Abstr. 1997, 65, 477–495. [Google Scholar]
- Paredes-Sánchez, F.A.; Sifuentes-Rincón, A.M.; Casas, E.; Arellano-Vera, W.; Parra-Bracamonte, G.M.; Riley, D.G.; Welsh, T.H., Jr.; Randel, R.D. Novel genes involved in the genetic architecture of temperament in Brahman cattle. PLoS ONE 2020, 15, e0237825. [Google Scholar] [CrossRef] [PubMed]
- Tulloh, N.M. Behaviour of cattle in yards. II. A study of temperament. Anim. Behav. 1961, 9, 25–30. [Google Scholar] [CrossRef]
- dos Santos, F.C.; Peixoto, M.G.C.D.; Fonseca, P.A.D.S.; Pires, M.D.F.; Ventura, R.V.; Rosse, I.D.C.; Bruneli, F.A.T.; Machado, M.A.; Carvalho, M.R.S. Identification of candidate genes for reactivity in guzerat (bos indicus) cattle: A genome-wide association study. PLoS ONE 2017, 12, e0169163. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The amount of shade influences the behavior and physiology of dairy cattle. J. Dairy Sci. 2010, 93, 125–133. [Google Scholar] [CrossRef]
- Reefmann, N.; Wechsler, B.; Gygax, L. Behavioural and physiological assessment of positive and negative emotion in sheep. Anim. Behav. 2009, 78, 651–659. [Google Scholar] [CrossRef]
- Graunke, K.L.; Nürnberg, G.; Repsilber, D.; Puppe, B.; Langbein, J. Describing Temperament in an Ungulate: A Multidimensional Approach. PLoS ONE 2013, 8, e74579. [Google Scholar] [CrossRef]
- Vallée, A.; Daures, J.; van Arendonk, J.A.M.; Bovenhuis, H. Genome-wide association study for behavior, type traits, and muscular development in Charolais beef cattle1. J. Anim. Sci. 2016, 94, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
- Blache, D.; Martin, G.; Maloney, S.K.; Martin, G. Towards Ethically Improved Animal Experimentation in the Study of Animal Reproduction. Reprod. Domest. Anim. 2008, 43, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A. Fear and Fearfulness in Animals. Q. Rev. Biol. 1995, 70, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Boulos, Z.; Terman, M. Food availability and daily biological rhythms. Neurosci. Biobehav. Rev. 1980, 4, 119–131. [Google Scholar] [CrossRef]
- Costilla, R.; Kemper, K.E.; Byrne, E.M.; Porto-Neto, L.R.; Carvalheiro, R.; Purfield, D.C.; Doyle, J.L.; Berry, D.P.; Moore, S.S.; Wray, N.R.; et al. Genetic control of temperament traits across species: Association of autism spectrum disorder risk genes with cattle temperament. Genet. Sel. Evol. 2020, 52, 51. [Google Scholar] [CrossRef]
- Nkrumah, J.D.; Crews, D.H., Jr.; Basarab, J.A.; Price, M.A.; Okine, E.K.; Wang, Z.; Li, C.; Moore, S.S. Genetic and phenotypic relationships of feeding behavior and temperament with performance, feed efficiency, ultrasound, and carcass merit of beef cattle1. J. Anim. Sci. 2007, 85, 2382–2390. [Google Scholar] [CrossRef]
- Haskell, M.J.; Esimm, G.; Turner, S.P. Genetic selection for temperament traits in dairy and beef cattle. Front. Genet. 2014, 5, 368. [Google Scholar] [CrossRef]
- Fordyce, G.; Goddard, M.; Seifert, G. The measurement of temperament in cattle and the effect of experience and genotype. In Proceedings of the Australian Society for Animal Production; Pergamon Press: Sydney, NSW, Australia, 1982; pp. 329–332. [Google Scholar]
- Voisinet, B.D.; Grandin, T.; Tatum, J.D.; O’Connor, S.F.; Struthers, J.J. Feedlot cattle with calm temperaments have higher average daily gains than cattle with excitable temperaments. J. Anim. Sci. 1997, 75, 892–896. [Google Scholar] [CrossRef]
- Cooke, R.F.; Kunkle, B.E. Interdisciplinary Beef Symposium: Temperament and acclimation to human handling influence growth, health, and reproductive responses in Bos taurus and Bos indicus cattle1. J. Anim. Sci. 2014, 92, 5325–5333. [Google Scholar] [CrossRef]
- Cooke, R.F.; Moriel, P.; Cappellozza, B.I.; Miranda, V.F.B.; Batista, L.F.D.; Colombo, E.A.; Ferreira, V.S.M.; Miranda, M.F.; Marques, R.S.; Vasconcelos, J.L.M. Effects of temperament on growth, plasma cortisol concentrations and puberty attainment in Nelore beef heifers. Animal 2019, 13, 1208–1213. [Google Scholar] [CrossRef]
- Francisco, C.L.; Resende, F.D.; Benatti, J.M.B.; Castilhos, A.M.; Cooke, R.F.; Jorge, A.M. Impacts of temperament on Nellore cattle: Physiological responses, feedlot performance, and carcass characteristics. J. Anim. Sci. 2015, 93, 5419–5429. [Google Scholar] [CrossRef] [PubMed]
- Blache, D.; Bickell, S.L. Temperament and reproductive biology: Emotional reactivity and reproduction in sheep Biologia temperamental e reprodutiva: Reatividade emocional e reprodução em ovinos. Rev. Bras. De Zootec. 2010, 39, 401–408. [Google Scholar] [CrossRef]
- Bickell, S.; Poindron, P.; Nowak, R.; Chadwick, A.; Ferguson, D.; Blache, D. Genotype rather than non-genetic behavioural transmission determines the temperament of Merino lambs. Anim. Welf. 2009, 18, 459–466. [Google Scholar] [CrossRef]
- Sebastian, T.; Watts, J.M.; Stookey, J.M.; Buchanan, F.; Waldner, C. Temperament in beef cattle: Methods of measurement and their relationship to production. Can. J. Anim. Sci. 2011, 91, 557–565. [Google Scholar] [CrossRef]
- Rietema, S.E.; Blackberry, M.A.; Maloney, S.K.; Martin, G.B.; Hawken, P.A.R.; Blache, D. Twenty-four-hour profiles of metabolic and stress hormones in sheep selected for a calm or nervous temperament. Domest. Anim. Endocrinol. 2015, 53, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef]
- Johnson, K.V.-A. Gut microbiome composition and diversity are related to human personality traits. Hum. Microbiome J. 2020, 15, 100069. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Tian, T.; Mao, Q.; Zou, T.; Zhou, C.-J.; Xie, J.; Chen, J.-J. Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl. Psychiatry 2020, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.-J.; Fan, S.-H.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Deusch, S.; Camarinha-Silva, A.; Conrad, J.; Beifuss, U.; Rodehutscord, M.; Seifert, J. A structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microbiol. 2017, 8, 1605. [Google Scholar] [CrossRef]
- Seshadri, R.; Leahy, S.C.; Attwood, G.T.; Teh, K.H.; Lambie, S.C.; Cookson, A.L.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Hadjithomas, M.; Varghese, N.J.; et al. Cultivation and sequencing of rumen microbiome members from the Hungate1000 Collection. Nat. Biotechnol. 2018, 36, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Kunkle, W.E.; Baker, F.S.; Palmer, A.Z. Factors Affecting Performance of Florida Steers and Heifers in the Feedlot; University of Florida: Gainesville, FL, USA, 1986; p. 87. [Google Scholar]
- Hammond, A.; Olson, T.; Chase Jr, C.; Bowers, E.; Randel, R.; Murphy, C.; Vogt, D.; Tewolde, A. Heat tolerance in two tropically adapted Bos taurus breeds, Senepol and Romosinuano, compared with Brahman, Angus, and Hereford cattle in Florida. J. Anim. Sci. 1996, 74, 295–303. [Google Scholar] [CrossRef]
- Baur, F.J.; Ensminger, L.G. Association of official analytical chemists. J. Am. Oil Chem. Soc. 1977, 54, 171–172. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Fortina, R.; Patrucco, S.G.; Barbera, S.; Tassone, S. Rumen Fluid from Slaughtered Animals: A Standardized Procedure for Sampling, Storage and Use in Digestibility Trials. Methods Protoc. 2022, 5, 59. [Google Scholar] [CrossRef]
- Chaudhry, A.S. Slaughtered cattle as source of rumen fluid to evaluate supplements for in vitro degradation of grass nuts and barley Straw. Open Vet. Sci. J. 2008, 2, 16–22. [Google Scholar] [CrossRef]
- Kristensen, N.B. Quantification of whole blood Short-chain fatty acids by gas chromatographic determination of plasma 2-chloroethyl derivatives and correction for dilution space in erythrocytes. Acta Agric. Scand. Sect. A Anim. Sci. 2000, 50, 231–236. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Enemark, J.M.D. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. Vet. J. 2008, 176, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Morgante, M.; Gianesella, M.; Casella, S.; Ravarotto, L.; Stelletta, C.; Giudice, E. Blood gas analyses, ruminal and blood pH, urine and faecal pH in dairy cows during subacute ruminal acidosis. Comp. Clin. Pathol. 2008, 18, 229–232. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.; Zheng, N.; Beckers, Y.; Wang, J. Urea metabolism and regulation by rumen bacterial urease in ruminants—A review. Ann. Anim. Sci. 2018, 18, 303–318. [Google Scholar] [CrossRef]
- Santoso, B.; Kilmaskossu, A.; Sambodo, P. Effects of saponin from Biophytum petersianum Klotzsch on ruminal fermentation, microbial protein synthesis and nitrogen utilization in goats. Anim. Feed. Sci. Technol. 2007, 137, 58–68. [Google Scholar] [CrossRef]
- McDonald, P.; Edwards, R.A.; Greenhalge, J.F.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, R.G. Animal Nutrition, 6th ed.; Pearson: London, UK, 2002. [Google Scholar]
- de Oliveira, M.N.V.; Jewell, K.A.; Freitas, F.S.; Benjamin, L.A.; Tótola, M.R.; Borges, A.C.; Moraes, C.A.; Suen, G. Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore steer. Vet. Microbiol. 2013, 164, 307–314. [Google Scholar] [CrossRef]
- Hook, S.E.; Steele, M.A.; Northwood, K.S.; Dijkstra, J.; France, J.; Wright, A.-D.G.; McBride, B.W. Impact of subacute ruminal acidosis (SARA) adaptation and recovery on the density and diversity of bacteria in the rumen of dairy cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef]
- Pitta, D.W.; Pinchak, W.E.; Dowd, S.; Dorton, K.; Yoon, I.; Min, B.R.; Fulford, J.D.; Wickersham, T.A.; Malinowski, D.P. Longitudinal shifts in bacterial diversity and fermentation pattern in the rumen of steers grazing wheat pasture. Anaerobe 2014, 30, 11–17. [Google Scholar] [CrossRef]
- Spence, C.; Wells, W.G.; Smith, C.J. Characterization of the Primary Starch Utilization Operon in the Obligate Anaerobe Bacteroides fragilis: Regulation by Carbon Source and Oxygen. J. Bacteriol. 2006, 188, 4663–4672. [Google Scholar] [CrossRef] [PubMed]
- Lamendella, R.; Santo Domingo, J.W.; Ghosh, S.; Martinson, J.; Oerther, D.B. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 2011, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Paster, B.J.; Canale-Parola, E. Physiological diversity of rumen spirochetes. Appl. Environ. Microbiol. 1982, 43, 686–693. [Google Scholar] [CrossRef]
- Stanton, T.B.; Canale-Parola, E. Enumeration and selective isolation of rumen spirochetes. Appl. Environ. Microbiol. 1979, 38, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Khol-Parisini, A.; Gruber, L.; Zebeli, Q. Microbial populations and fermentation profiles in rumen liquid and solids of Holstein cows respond differently to dietary barley processing. J. Appl. Microbiol. 2015, 119, 1502–1514. [Google Scholar] [CrossRef] [PubMed]
- Golder, H.; Denman, S.; McSweeney, C.; Wales, W.; Auldist, M.; Wright, M.; Marett, L.; Greenwood, J.; Hannah, M.; Celi, P.; et al. Effects of partial mixed rations and supplement amounts on milk production and composition, ruminal fermentation, bacterial communities, and ruminal acidosis. J. Dairy Sci. 2014, 97, 5763–5785. [Google Scholar] [CrossRef]
- Pitta, D.W.; Pinchak, W.E.; Indugu, N.; Vecchiarelli, B.; Sinha, R.; Fulford, J.D. Metagenomic analysis of the rumen microbiome of steers with wheat-induced frothy bloat. Front. Microbiol. 2016, 7, 689. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef]
- Neumann, A.P.; McCormick, C.A.; Suen, G. Fibrobacter communities in the gastrointestinal tracts of diverse hindgut-fermenting herbivores are distinct from those of the rumen: Ecology of Fibrobacter in hindgut-fermenting herbivores. Environ. Microbiol. 2017, 19, 3768–3783. [Google Scholar] [CrossRef]
- Stanton, T.B.; Canale-Parola, E. Treponema bryantii sp. nov., a rumen spirochete that interacts with cellulolytic bacteria. Arch. Microbiol. 1980, 127, 145–156. [Google Scholar] [CrossRef]
- Huws, S.A.; Kim, E.J.; Lee, M.R.F.; Scott, M.B.; Tweed, J.K.S.; Pinloche, E.; Wallace, R.J.; Scollan, N.D. As yet uncultured bacteria phylogenetically classified as Prevotella, Lachnospiraceae incertae sedis and unclassified Bacteroidales, Clostridiales and Ruminococcaceae may play a predominant role in ruminal biohydrogenation. Environ. Microbiol. 2011, 13, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Gagen, E.J.; Padmanabha, J.; Denman, S.E.; McSweeney, C.S. Hydrogenotrophic culture enrichment reveals rumen Lachnospiraceae and Ruminococcaceae acetogens and hydrogen-responsive Bacteroidetes from pasture-fed cattle. FEMS Microbiol. Lett. 2015, 362, fnv104. [Google Scholar] [CrossRef] [PubMed]
- Opdahl, L.J.; Gonda, M.G.; St-Pierre, B. Identification of Uncultured Bacterial Species from Firmicutes, Bacteroidetes and CANDIDATUS Saccharibacteria as Candidate Cellulose Utilizers from the Rumen of Beef Cows. Microorganisms 2018, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Brulc, J.M.; Antonopoulos, D.A.; Miller, M.E.B.; Wilson, M.K.; Yannarell, A.C.; Dinsdale, E.A.; Edwards, R.E.; Frank, E.D.; Emerson, J.B.; Wacklin, P.; et al. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. USA 2009, 106, 1948–1953. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content (%) |
|---|---|
| Silage corn | 24.80 |
| Leymus chinensis | 15.20 |
| Corn | 18.00 |
| Wheat bran | 9.78 |
| Soybean meal | 16.20 |
| Brown rice | 8.52 |
| Beer yeast | 4.02 |
| Corn gluten meal | 1.20 |
| Calcium dihydrogen phosphate | 0.72 |
| Stone powder | 0.96 |
| Salt | 0.30 |
| Premix (1) | 0.30 |
| Total | 100.00 |
| Nutrient (2) | |
| Digestible Energy (MJ/kg) | 13.12 |
| Crude protein (%) | 20.72 |
| Neutral detergent fiber (%) | 31.80 |
| Acid detergent fiber (%) | 19.62 |
| Ether extract (%) | 2.10 |
| Calcium (%) | 0.74 |
| Total phosphorus (%) | 0.51 |
| Items | Calm Group | Nervous Group | p-Value |
|---|---|---|---|
| Bodyweight (kg) | |||
| Start on the experiment | 29.7 ± 2.2 | 30.0 ± 1.8 | 0.30 |
| End of the experiment | 47.7 ± 0.23 | 49.3± 1.63 | 0.20 |
| Average daily weight gain (kg/d) | 0.26 ± 0.22 | 0.36 ± 0.05 | 0.49 |
| Daily dry matter intake (kg/d) | 1.48 ± 0.17 | 1.51 ± 0.23 | 0.30 |
| Items | Calm Group | Nervous Group | p-Value |
|---|---|---|---|
| pH | 6.70 ± 0.10 | 6.57 ± 0.06 | 0.116 |
| Acetic acid (mmol/L) | 15.86 ± 1.71 | 16.52 ± 1.06 | 0.754 |
| Propionic acid (mmol/L) | 2.71 ± 0.23 | 3.55 ± 0.28 | 0.045 |
| Isobutyric acid (mmol/L) | 0.28 ± 0.03 | 0.36 ± 0.05 | 0.194 |
| Butyrate acid (mmol/L) | 1.13 ± 0.18 | 1.41 ± 0.23 | 0.372 |
| Isovaleric acid (mmol/L) | 0.30 ± 0.03 | 0.45 ± 0.04 | 0.018 |
| Valeric acid (mmol/L) | 0.13 ± 0.01 | 0.21 ± 0.03 | 0.022 |
| Total VFA concentration (mmol/L) | 20.43 ± 1.83 | 22.50 ± 1.68 | 0.424 |
| Acetic acid/acid ratio (%) | 5.00 ± 0.80 | 4.67 ± 0.08 | 0.134 |
| Ammonia nitrogen concentration (mg/100 mL) | 3.68 ± 0.43 | 1.94 ± 0.09 | 0.007 |
| Alpha Diversity Index | Calm Group | Nervous Group | p-Value |
|---|---|---|---|
| Chao value | 1288.51 ± 12.08 | 1279.92 ± 13.77 | 0.649 |
| Shannon index | 7.78 ± 0.06 | 8.15 ± 0.03 | < 0.01 |
| Simpson index | 0.9850 ± 0.001 | 0.9907 ± 0.0002 | < 0.01 |
| Phylum | Calm Group | Nervous Group | p-Value |
|---|---|---|---|
| Bacteroidetes | 55.72 ± 0.64 | 58.74 ± 1.04 | 0.033 |
| Firmicutes | 34.78 ± 0.65 | 29.63 ± 0.39 | < 0.01 |
| Proteobacteria | 2.87 ± 0.44 | 3.30 ± 0.83 | 0.650 |
| Spirochaetes | 2.04 ± 0.24 | 3.06 ± 0.12 | 0.004 |
| Tenericutes | 2.69 ± 0.10 | 2.32 ± 0.08 | 0.020 |
| Genus | Calm Group | Nervous Group | p-Value |
|---|---|---|---|
| Prevotella 1 | 16.77 ± 1.42 | 16.60 ± 0.70 | 0.917 |
| Christensenellaceae R-7 group | 13.37 ± 0.82 | 6.91 ± 0.61 | < 0.01 |
| Prevotellaceae UCG-001 | 3.35 ± 0.10 | 2.98 ± 0.58 | 0.546 |
| Ruminococcaceae NK4A214 group | 3.20 ± 0.32 | 3.03 ± 0.19 | 0.649 |
| Treponema 2 | 1.56 ± 0.21 | 2.31 ± 0.60 | 0.006 |
| Ruminococcaceae UCG-010 | 1.40 ± 0.96 | 1.65 ± 0.75 | 0.067 |
| Ruminococcaceae UCG-003 | 1.05 ± 0.05 | 1.38 ± 0.10 | 0.014 |
| Fibrobacter | 0.76 ± 0.19 | 1.87 ± 0.11 | < 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Ding, L.; Wang, J.; Chen, Q.; Thapa, A.; Mao, J.; Wang, M. Calm Hu Sheep Have a Different Microbiome Profile and Higher Energy Utilization Efficiency Than Nervous Hu Sheep. Fermentation 2023, 9, 470. https://doi.org/10.3390/fermentation9050470
Wu F, Ding L, Wang J, Chen Q, Thapa A, Mao J, Wang M. Calm Hu Sheep Have a Different Microbiome Profile and Higher Energy Utilization Efficiency Than Nervous Hu Sheep. Fermentation. 2023; 9(5):470. https://doi.org/10.3390/fermentation9050470
Chicago/Turabian StyleWu, Feifan, Luoyang Ding, Jiasheng Wang, Qiaoqing Chen, Asmita Thapa, Jianyu Mao, and Mengzhi Wang. 2023. "Calm Hu Sheep Have a Different Microbiome Profile and Higher Energy Utilization Efficiency Than Nervous Hu Sheep" Fermentation 9, no. 5: 470. https://doi.org/10.3390/fermentation9050470
APA StyleWu, F., Ding, L., Wang, J., Chen, Q., Thapa, A., Mao, J., & Wang, M. (2023). Calm Hu Sheep Have a Different Microbiome Profile and Higher Energy Utilization Efficiency Than Nervous Hu Sheep. Fermentation, 9(5), 470. https://doi.org/10.3390/fermentation9050470