
Utilization of Meat and Bone Meal for Yeast Fermentation to Produce Astaxanthin


Abstract
:1. Introduction
2. Materials and Methods
2.1. Meat and Bone Meal and Chemicals
2.2. Microbial Cultivation
2.3. Optimization of Carbon-to-Nitrogen (C/N) Ratio on Astaxanthin Production
2.3.1. Effect of Glucose Loading on Astaxanthin Production
2.3.2. Effect of Nitrogen Loading on Astaxanthin Production
2.4. Meat and Bone Meal Fermentation
2.4.1. MBM Defatting
2.4.2. Enzymatic and Alkaline Hydrolysis of Meat and Bone Meal
2.4.3. Astaxanthin Fermentation with Meat and Bone Meal Hydrolysates
2.4.4. Astaxanthin Fermentation with Lyophilized Meat and Bone Meal Hydrolysates
2.5. Analytical Methods
2.6. Data Analysis
3. Results and Discussion
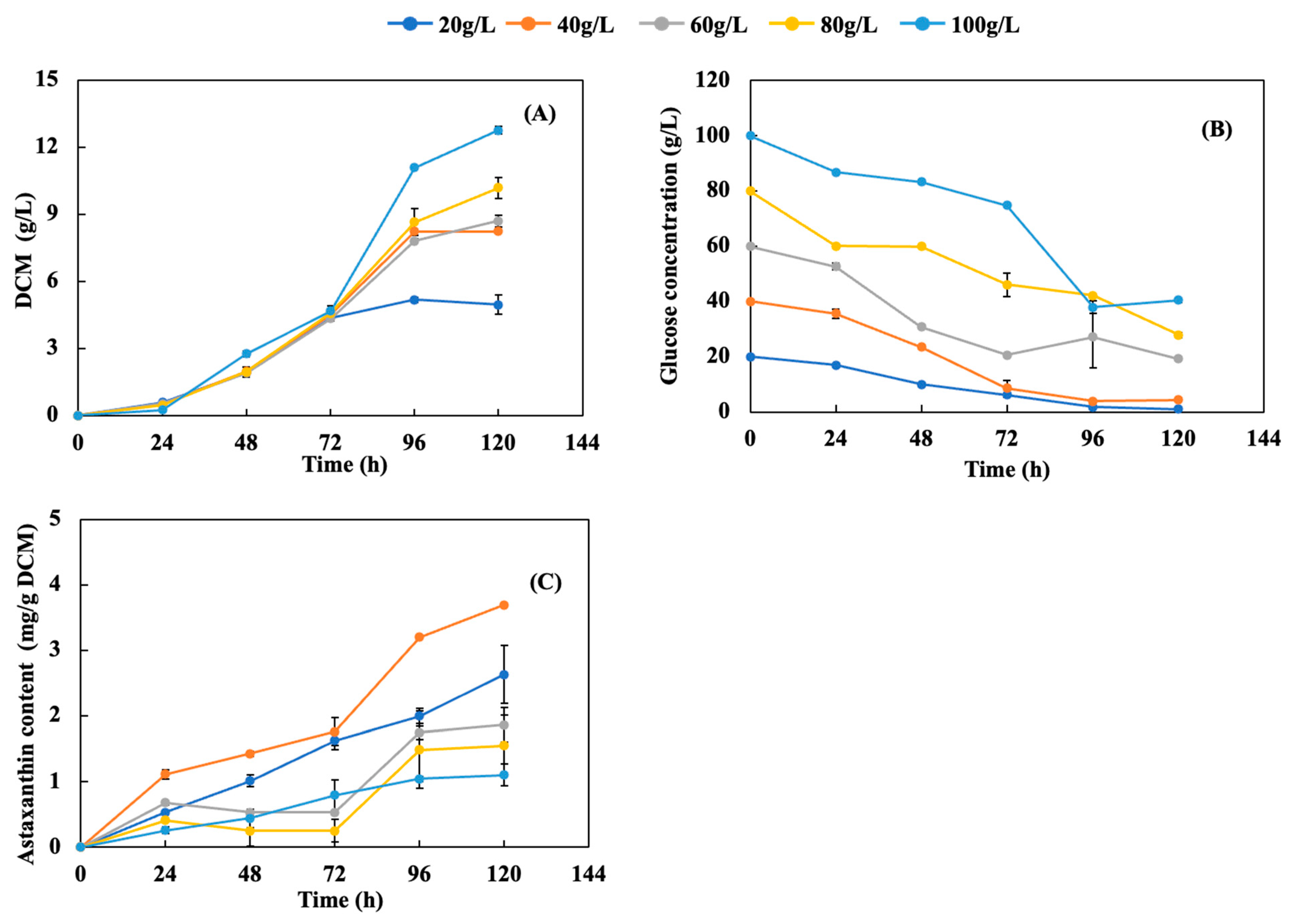
3.1. Effect of Glucose Loading on Astaxanthin Production
3.2. Effects of Nitrogen Loading on Astaxanthin Production
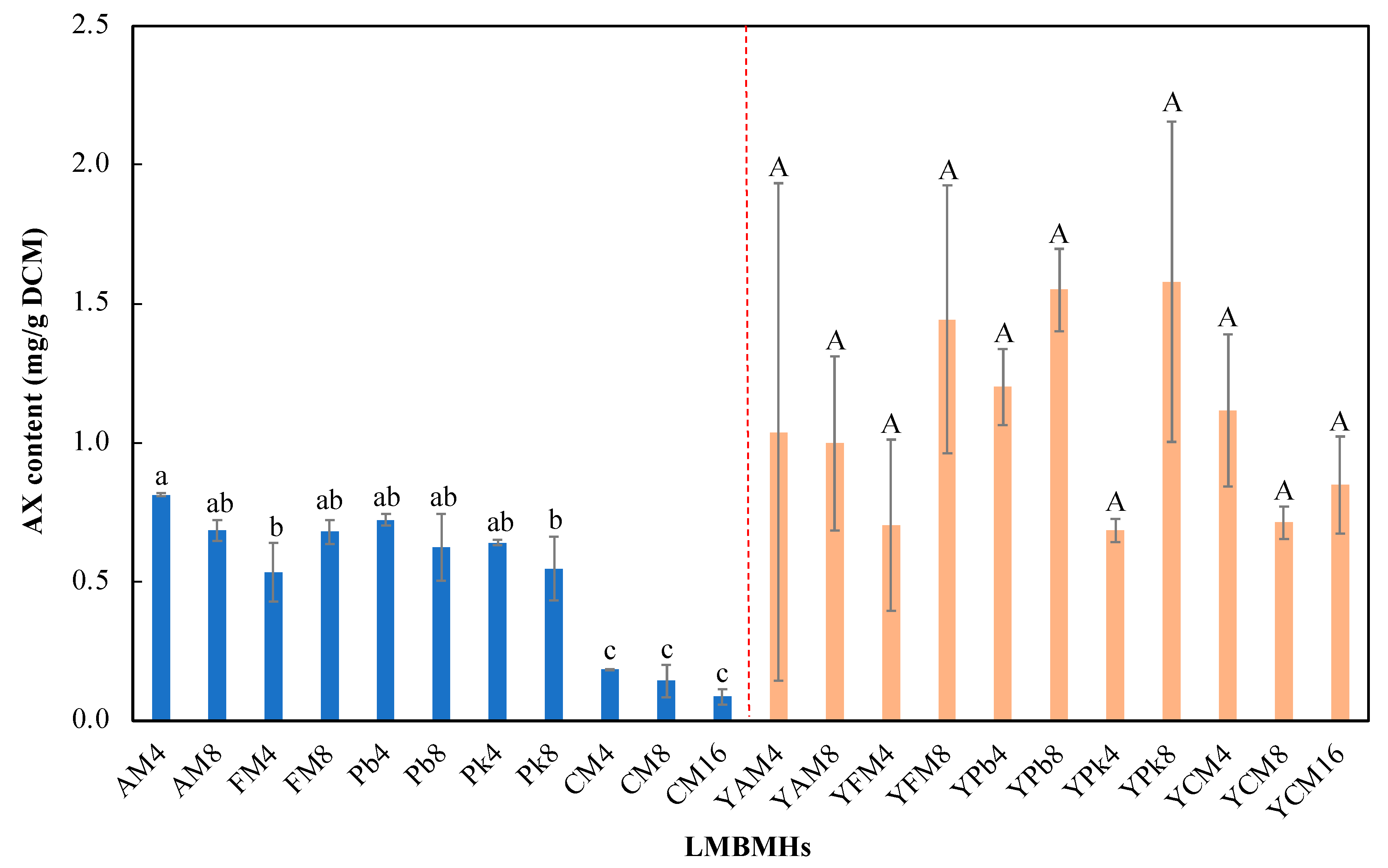
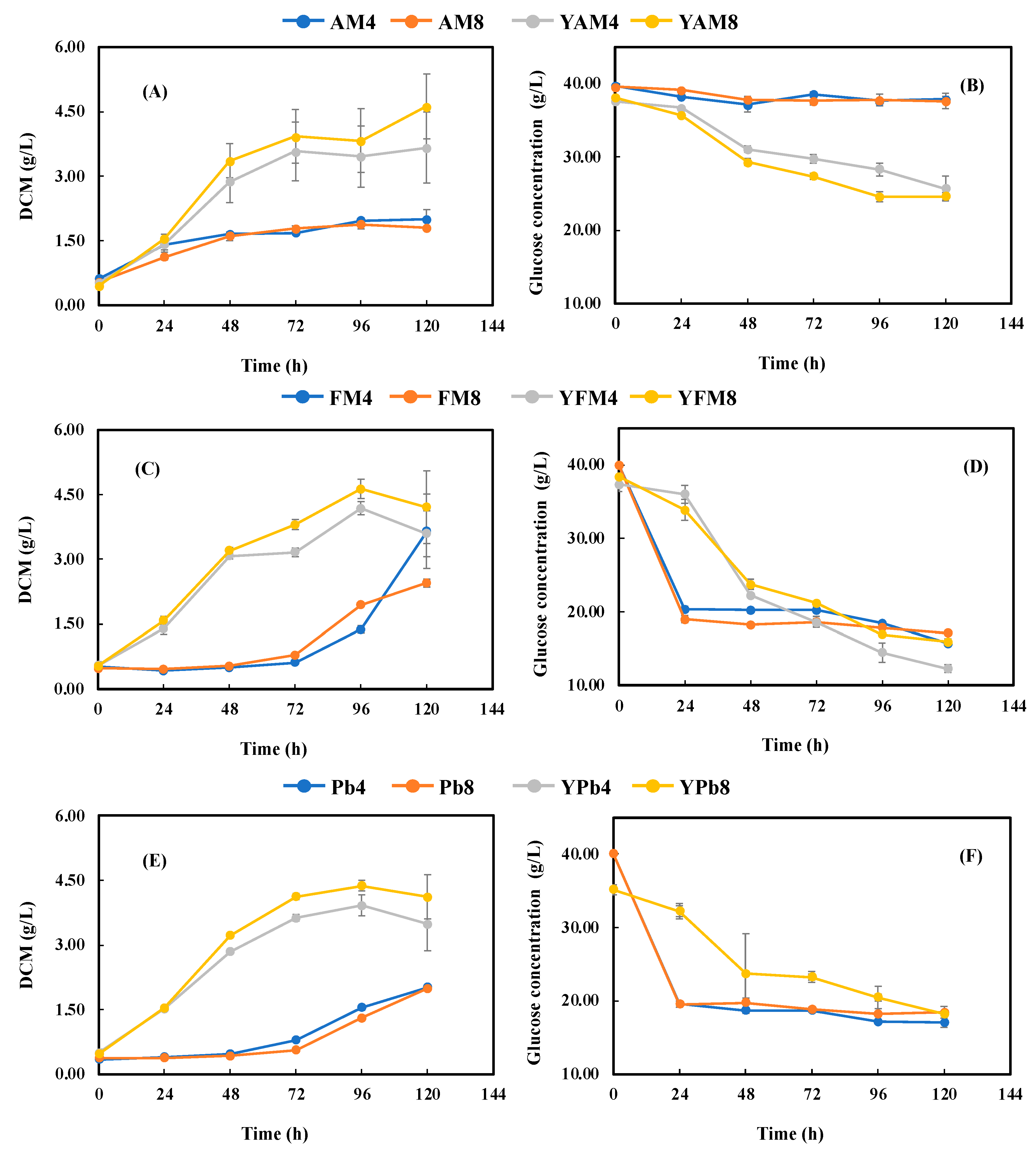
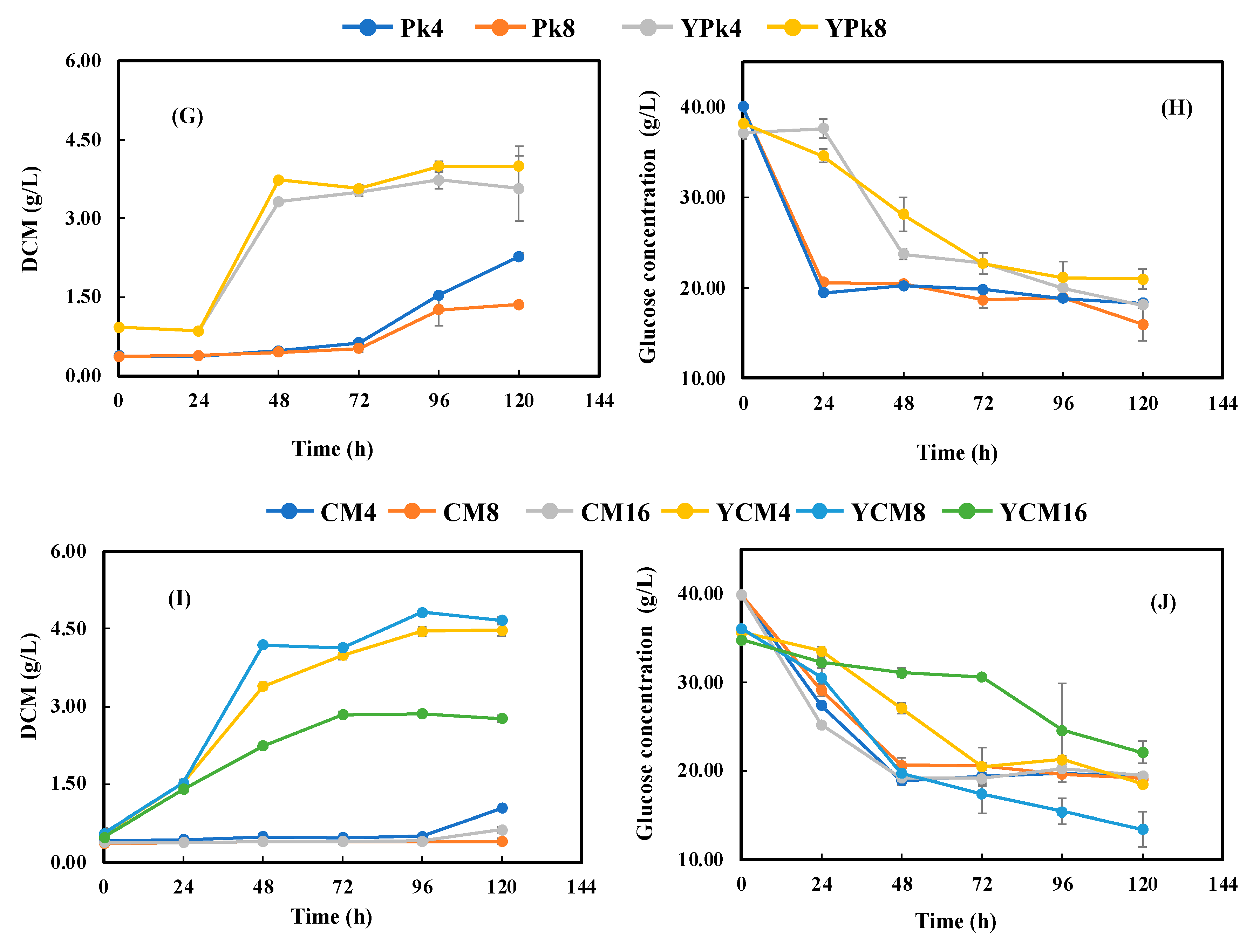
3.3. Astaxanthin Production with Meat and Bone Meal Hydrolysates (MBMHs) as Nitrogen Sources
3.4. Fermentation with Lyophilized Meat and Bone Meal Hydrolysates (LMBMHs) as Nitrogen Sources
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swisher, K. US Market Report—A mixed bag during a disruptive year. Render 2021, 10–15. [Google Scholar]
- Garcia, R.A.; Rosentrater, K.A. Concentration of Key Elements in North American Meat & Bone Meal. Biomass Bioenergy 2008, 32, 887–891. [Google Scholar] [CrossRef]
- Beck, J.; Brandenstein, J.; Unterberger, S.; Hein, K.R.G. Effects of Sewage Sludge and Meat and Bone Meal Co-Combustion on SCR Catalysts. Appl. Catal. B-Environ. 2004, 49, 15–25. [Google Scholar] [CrossRef]
- Aranda Uson, A.; Lopez-Sabiron, A.M.; Ferreira, G.; Llera Sastresa, E. Uses of Alternative Fuels and Raw Materials in the Cement Industry as Sustainable Waste Management Options. Renew. Sustain. Energy Rev. 2013, 23, 242–260. [Google Scholar] [CrossRef]
- Piazza, G.J.; Garcia, R.A. Meat & Bone Meal Extract and Gelatin as Renewable Flocculants. Bioresour. Technol. 2010, 101, 781–787. [Google Scholar] [CrossRef]
- Jeng, A.; Haraldsen, T.K.; Vagstad, N.; Gronlund, A. Meat and Bone Meal as Nitrogen Fertilizer to Cereals in Norway. Agric. Food Sci. 2004, 13, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Garcia, R.A.; Piazza, G.J.; Wen, Z.; Pyle, D.J.; Solaiman, D.K.Y. The Non-Nutritional Performance Characteristics of Peptones Made from Rendered Protein. J. Ind. Microbiol. Biotechnol. 2010, 37, 95–102. [Google Scholar] [CrossRef]
- Garcia, R.A.; Pyle, D.J.; Piazza, G.J.; Wen, Z. Hydrolysis of Animal Protein Meals for Improved Utility in Non-Feed Applications. Appl. Eng. Agric. 2011, 27, 269–275. [Google Scholar] [CrossRef]
- Liang, Y.; Garcia, R.A.; Piazza, G.J.; Wen, Z. Nonfeed Application of Rendered Animal Proteins for Microbial Production of Eicosapentaenoic Acid by the Fungus Pythium irregulare. J. Agric. Food Chem. 2011, 59, 11990–11996. [Google Scholar] [CrossRef]
- Batghare, A.H.; Moholkar, V.S. Production of Nutraceutical Astaxanthin from Waste Resources. In Waste Biorefinery: Value Addition through Resource Utilization; Bhaskar, T., Varjani, S., Pandey, A., Rene, E.R., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2021; pp. 181–205. [Google Scholar] [CrossRef]
- Liu, X.; Xie, J.; Zhou, L.; Zhang, J.; Chen, Z.; Xiao, J.; Cao, Y.; Xiao, H. Recent Advances in Health Benefits and Bioavailability of Dietary Astaxanthin and Its Isomers. Food Chem. 2023, 404, 134605. [Google Scholar] [CrossRef]
- Aneesh, P.A.; Ajeeshkumar, K.K.; Lekshmi, R.G.K.; Anandan, R.; Ravishankar, C.N.; Mathew, S. Bioactivities of Astaxanthin from Natural Sources, Augmenting Its Biomedical Potential: A Review. Trends Food Sci. Technol. 2022, 125, 81–90. [Google Scholar] [CrossRef]
- Ni, H.; Chen, Q.; Ruan, H.; Yang, Y.; Li, L.; Wu, G.; Hu, Y.; He, G. Studies on Optimization of Nitrogen Sources for Astaxanthin Production by Phaffia rhodozyma. J. Zhejiang Univ. Sci. B 2007, 8, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamane, Y.; Higashida, K.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Influence of Oxygen and Glucose on Primary Metabolism and Astaxanthin Production by Phaffia rhodozyma in Batch and Fed-Batch Cultures: Kinetic and Stoichiometric Analysis. Appl. Environ. Microbiol. 1997, 63, 4471–4478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Ling, X.; Zheng, Z.; Xiong, W.; Lu, Y. Effect of Vitamins on Astaxanthin Production of Phaffia rhodozyma. J. Xiamen Univ. (Nat. Sci.) 2011, 50, 111–115. [Google Scholar]
- Nghiem, N.P.; Montanti, J.; Johnston, D. Production of Astaxanthin from Corn Fiber as a Value-Added Co-Product of Fuel Ethanol Fermentation. Appl. Biochem. Biotechnol. 2009, 154, 227–237. [Google Scholar] [CrossRef]
- Sriperm, N.; Pesti, G.M.; Tillman, P.B. Evaluation of the Fixed Nitrogen-to-Protein (N:P) Conversion Factor (6.25) versus Ingredient Specific N:P Conversion Factors in Feedstuffs. J. Sci. Food Agric. 2011, 91, 1182–1186. [Google Scholar] [CrossRef]
- Xiao, R.; Yang, X.; Li, M.; Li, X.; Wei, Y.; Cao, M.; Ragauskas, A.; Thies, M.; Ding, J.; Zheng, Y. Investigation of Composition, Structure and Bioactivity of Extracellular Polymeric Substances from Original and Stress-Induced Strains of Thraustochytrium striatum. Carbohydr. Polym. 2018, 195, 515–524. [Google Scholar] [CrossRef]
- Stoklosa, R.J.; Johnston, D.B.; Nghiem, N.P. Phaffia rhodozyma Cultivation on Structural and Non-Structural Sugars from Sweet Sorghum for Astaxanthin Generation. Process Biochem. 2019, 83, 9–17. [Google Scholar] [CrossRef]
- Reynders, M.B.; Rawlings, D.E.; Harrison, S.T.L. Demonstration of the Crabtree Effect in Phaffia rhodozyma during Continuous and Fed-Batch Cultivation. Biotechnol. Lett. 1997, 19, 549–552. [Google Scholar] [CrossRef]
- Flores-Cotera, L.B.; Chávez-Cabrera, C.; Martínez-Cárdenas, A.; Sánchez, S.; García-Flores, O.U. Deciphering the Mechanism by Which the Yeast Phaffia rhodozyma Responds Adaptively to Environmental, Nutritional, and Genetic Cues. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab048. [Google Scholar] [CrossRef]
- Miao, L.; Chi, S.; Wu, M.; Liu, Z.; Li, Y. Deregulation of Phytoene-β-Carotene Synthase Results in Derepression of Astaxanthin Synthesis at High Glucose Concentration in Phaffia rhodozyma Astaxanthin-Overproducing Strain MK19. BMC Microbiol. 2019, 19, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amado, I.R.; Vázquez, J.A. Mussel Processing Wastewater: A Low-Cost Substrate for the Production of Astaxanthin by Xanthophyllomyces dendrorhous. Microb. Cell Factories 2015, 14, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Wang, B.; Gerken, H.G.; Lu, Y.; Ling, X. Proteomic Analysis of Astaxanthin Biosynthesis in Xanthophyllomyces dendrorhous in Response to Low Carbon Levels. Bioprocess Biosyst. Eng. 2017, 40, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Cabrera, C.; Flores-Bustamante, Z.R.; Marsch, R.; Montes, M.d.C.; Sánchez, S.; Cancino-Díaz, J.C.; Flores-Cotera, L.B. ATP-Citrate Lyase Activity and Carotenoid Production in Batch Cultures of Phaffia rhodozyma under Nitrogen-Limited and Nonlimited Conditions. Appl. Microbiol. Biotechnol. 2010, 85, 1953–1960. [Google Scholar] [CrossRef]
- Chávez-Cabrera, C.; Marsch, R.; Bartolo-Aguilar, Y.; Flores-Bustamante, Z.R.; Hidalgo-Lara, M.E.; Martínez-Cárdenas, A.; Cancino-Díaz, J.C.; Sánchez, S.; Flores-Cotera, L.B. Molecular Cloning and Characterization of the ATP Citrate Lyase from Carotenogenic Yeast Phaffia rhodozyma. FEMS Yeast Res. 2015, 15, fov054. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Pan, X.; Jia, J.; Xiong, W.; Manirafasha, E.; Ling, X.; Yinghua, L. Strategy and Regulatory Mechanisms of Glutamate Feeding to Enhance Astaxanthin Yield in Xanthophyllomyces dendrorhous. Enzym. Microb. Technol. 2019, 125, 45–52. [Google Scholar] [CrossRef]
- Taheri, A.; Abedian Kenari, A.; Motamedzadegan, A.; Habibi-Rezaei, M. Poultry By-Products and Enzymatic Hydrolysis: Optimization by Response Surface Methodology Using Alcalase® 2.4 L. Int. J. Food Eng. 2011, 7. [Google Scholar] [CrossRef]
- Fang, T.J.; Wang, J.-M. Extractability of Astaxanthin in a Mixed Culture of a Carotenoid Over-Producing Mutant of Xanthophyllomyces dendrorhous and Bacillus Circulans in Two-Stage Batch Fermentation. Process Biochem. 2002, 37, 1235–1245. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, S.-W.; Kim, S.-W.; Chang, H.-I. High-Level Production of Astaxanthin by Xanthophyllomyces dendrorhous Mutant JH1 Using Statistical Experimental Designs. Biosci. Biotechnol. Biochem. 2005, 69, 1743–1748. [Google Scholar] [CrossRef]
- Stoklosa, R.J.; Johnston, D.B.; Nghiem, N.P. Utilization of Sweet Sorghum Juice for the Production of Astaxanthin as a Biorefinery Co-Product by Phaffia rhodozyma. ACS Sustain. Chem. Eng. 2018, 6, 3124–3134. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition | Values | ||||
|---|---|---|---|---|---|
| Glucose (g/L) | 40 | ||||
| Yeast extract + peptone (g/L) | 3.0 + 5.0 | 1.5 + 2.5 | 1.0 + 1.6 | 0.7 + 1.2 | 0.6 + 1.0 |
| C/N (element ratio) | 15 | 30 | 47 | 64 | 76 |
| “C/N” (g/g) | 5 (N5) | 10 (N10) | 15 (N15) | 20 (N20) | 25 (N25) |
| Hydrolysis Type | Reagent | Hydrolysis Condition | Label a |
|---|---|---|---|
| Enzymatic hydrolysis | Alcalase® 2.4L | Alcalase® 2.4L, 0.4 U/g substrate, 50 °C, pH 8.5, 4 h | AM4 |
| Alcalase® 2.4L,0.4 U/g substrate, 50 °C, pH 8.5, 8 h | AM8 | ||
| Flavourzyme® | Flavourzyme®, 50 LAPU/g substrate, pH 7.0, 50 °C, 4 h | FM4 | |
| Flavourzyme®, 50 LAPU/g substrate, pH 7.0, 50 °C, 8 h | FM8 | ||
| Proteinase K | 1 mg/g MBM, pH 7.5, 37 °C, 4 h | Pk4 | |
| 1 mg/g MBM, pH 7.5, 37 °C, 8 h | Pk8 | ||
| Bacterial proteinase | 1 mg/g MBM, pH 7.5, 37 °C, 4 h | Pb4 | |
| 1 mg/g MBM, pH 7.5, 37 °C, 8 h | Pb8 | ||
| Alkaline hydrolysis | Calcium hydroxide | 0.1 g/g MBM, 85 °C, 4 h | CM4 |
| 0.1 g/g MBM, 85 °C, 8 h | CM8 | ||
| 0.1 g/g MBM, 85 °C, 16 h | CM16 |
| MBMHs | Nitrogen Content (wt%, g/g Medium) | DCM (g/L) | Astaxanthin Content (mg/g DCM) | Residual Glucose Concentration (g/L) |
|---|---|---|---|---|
| AM4 | 2.29 ± 0.03 | - | - d | - |
| AM8 | 2.17 ± 0.08 | - | - d | - |
| FM4 | 2.17 ± 0.04 | - | - d | - |
| FM8 | 2.78 ± 0.02 | 2.28 ± 0.04 | 0.09 ± 0.05 d | 16 ± 1.96 |
| Pb4 | 1.84 ± 0 | - | - | - |
| Pb8 | 2.2 ± 0.01 | 3.06 ± 0.48 | 0.52 ± 0.14 c | 13.04 ± 0.5 |
| Pk4 | 1.9 ± 0.02 | - | - | - |
| Pk8 | 1.85 ± 0.05 | 4.96 ± 1.44 | 0.75 ± 0.11 bc | 11.55 ± 0.39 |
| CM4 | 3.1 ± 0 | 4.66 ± 0.56 | 1.02 ± 0.12 a | 14.81 ± 1.13 |
| CM8 | 2.96 ± 0.01 | 3.85 ± 1.19 | 0.79 ± 0.03 ab | 12.68 ± 0.8 |
| CM16 | 2.91 ± 0.01 | 5.71 ± 0.49 | 0.81 ± 0.05 ab | 13.13 ± 1 |
| YAM4 | 2.29 ± 0.03 | - | - A | - |
| YAM8 | 2.17 ± 0.08 | - | - A | - |
| YFM4 | 2.17 ± 0.04 | - | - A | - |
| YFM8 | 2.78 ± 0.02 | 10.18 ± 0.44 | 0.57 ± 0.61 A | 17.54 ± 1.29 |
| YPb4 | 1.84 ± 0 | - | - | - |
| YPb8 | 2.2 ± 0.01 | 4.36 ± 0.04 | 0.66 ± 0.05 A | 9.39 ± 0.61 |
| YPk4 | 1.9 ± 0.02 | - | - | - |
| YPk8 | 1.85 ± 0.05 | 5.99 ± 0.37 | 0.71 ± 0.06 A | 5.72 ± 0.18 |
| YCM4 | 3.1 ± 0 | 10.89 ± 2.15 | 0.21 ± 0.04 A | 9.34 ± 2.81 |
| YCM8 | 2.96 ± 0.01 | 4.08 ± 0.48 | 0.35 ± 0.12 A | 10.85 ± 0.42 |
| YCM16 | 2.91 ± 0.01 | 11.47 ± 0.99 | 0.26 ± 0.12 A | 5.9 ± 1.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Zheng, Y. Utilization of Meat and Bone Meal for Yeast Fermentation to Produce Astaxanthin. Fermentation 2023, 9, 630. https://doi.org/10.3390/fermentation9070630
Li S, Zheng Y. Utilization of Meat and Bone Meal for Yeast Fermentation to Produce Astaxanthin. Fermentation. 2023; 9(7):630. https://doi.org/10.3390/fermentation9070630
Chicago/Turabian StyleLi, Sang, and Yi Zheng. 2023. "Utilization of Meat and Bone Meal for Yeast Fermentation to Produce Astaxanthin" Fermentation 9, no. 7: 630. https://doi.org/10.3390/fermentation9070630
APA StyleLi, S., & Zheng, Y. (2023). Utilization of Meat and Bone Meal for Yeast Fermentation to Produce Astaxanthin. Fermentation, 9(7), 630. https://doi.org/10.3390/fermentation9070630