
Characteristics of a Recombinant Lentinula edodes Ferulic Acid Esterase and Its Adverse Effects on In Vitro Fermentation of Wheat Straw

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
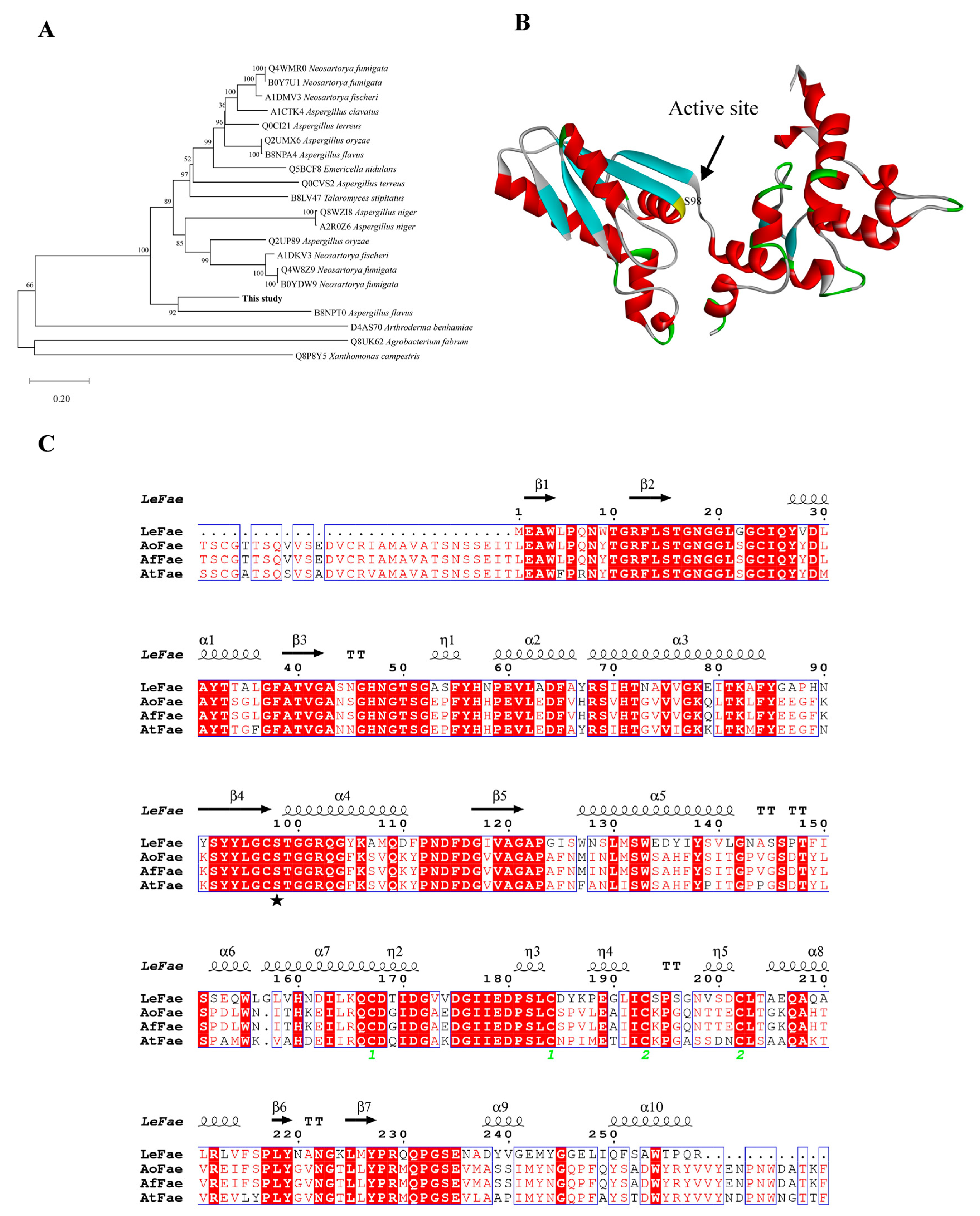
2.1. Analysis of Sequence Features
2.2. Construction of the Recombinant Strain
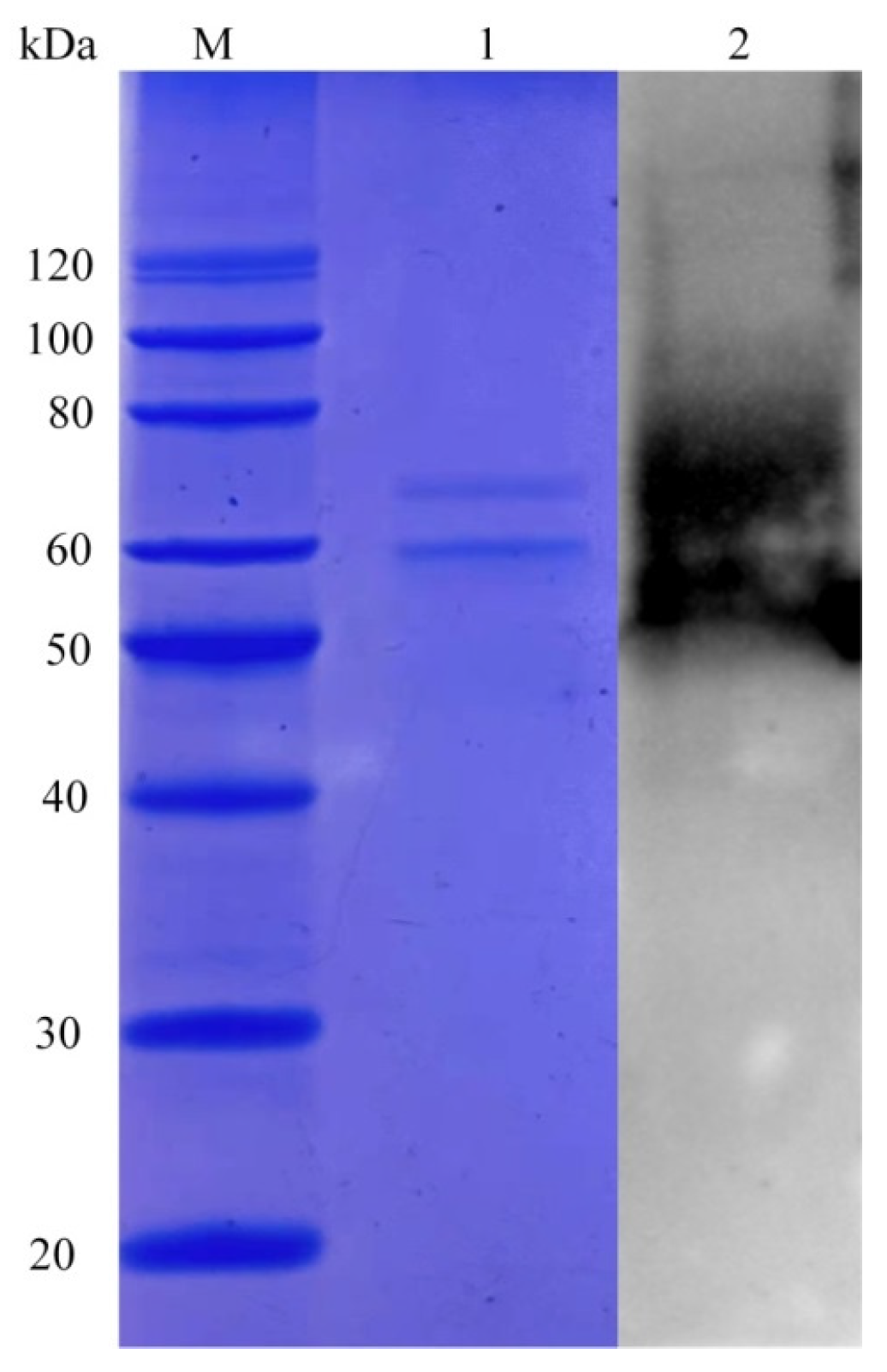
2.3. Laboratory-Scale Production of LeFae
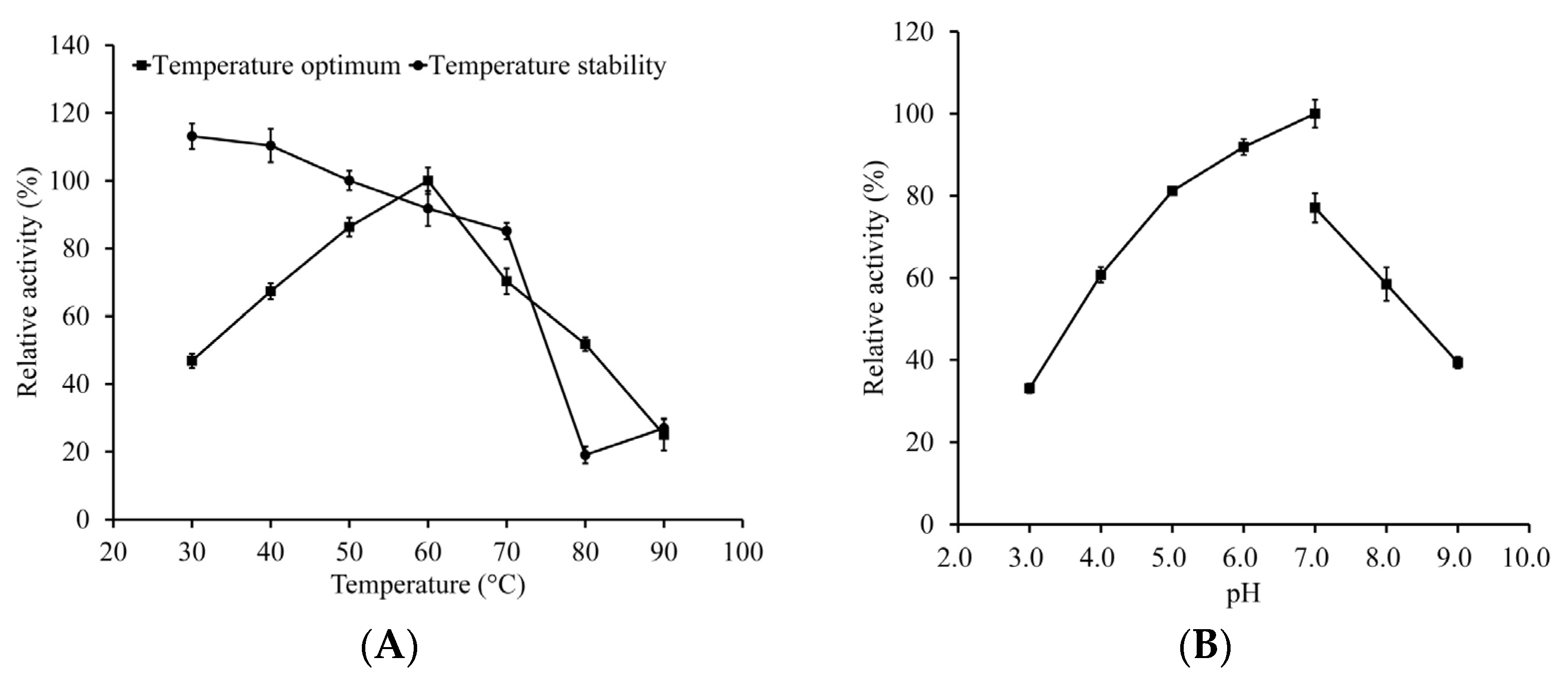
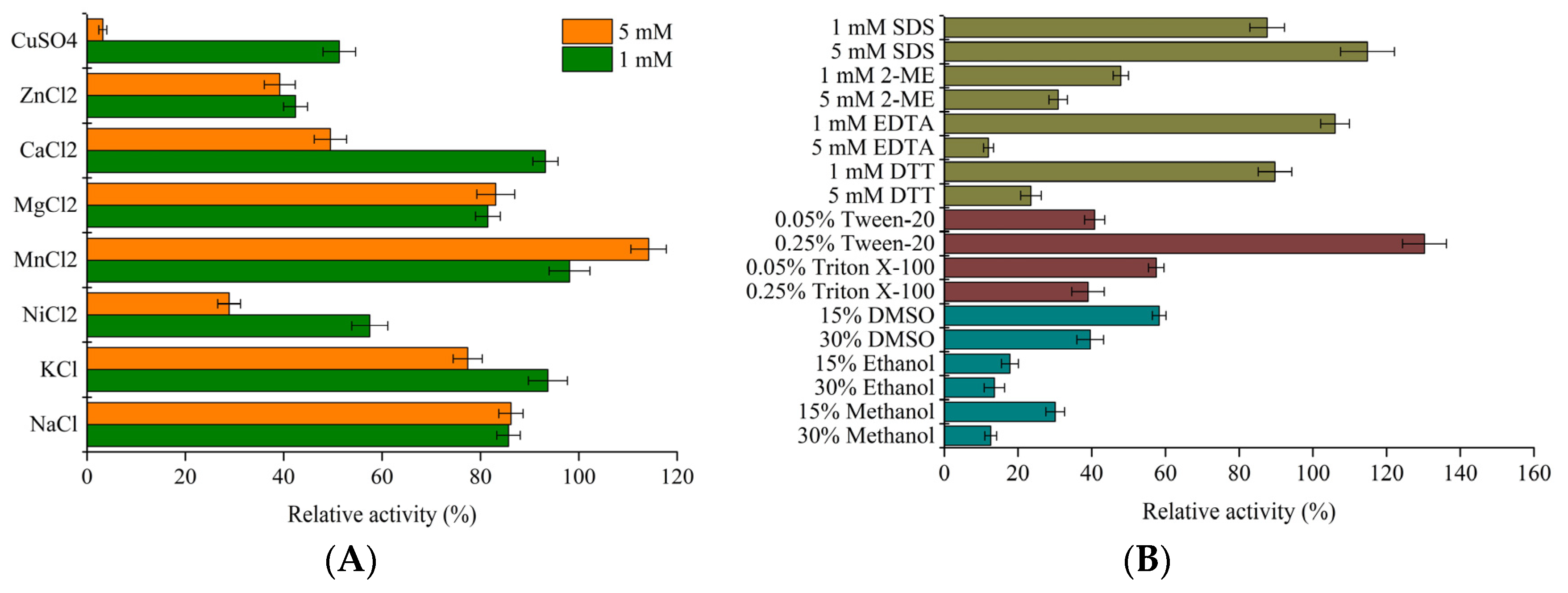
2.4. Characterization of LeFae
2.5. Hydrolysis of Wheat Straw Using LeFae
2.6. Synergism of LeFae on the Enzymatic Hydrolysis of Wheat Straw
2.7. In Vitro Fermentation of Wheat Straw
2.8. In Vitro Fermentation of Wheat Straw Pretreated with LeFae
2.9. Analytical Procedures
2.10. Statistical Analyses
3. Results and Discussion
3.1. Analysis and Production of LeFae
3.2. Characterization of LeFae
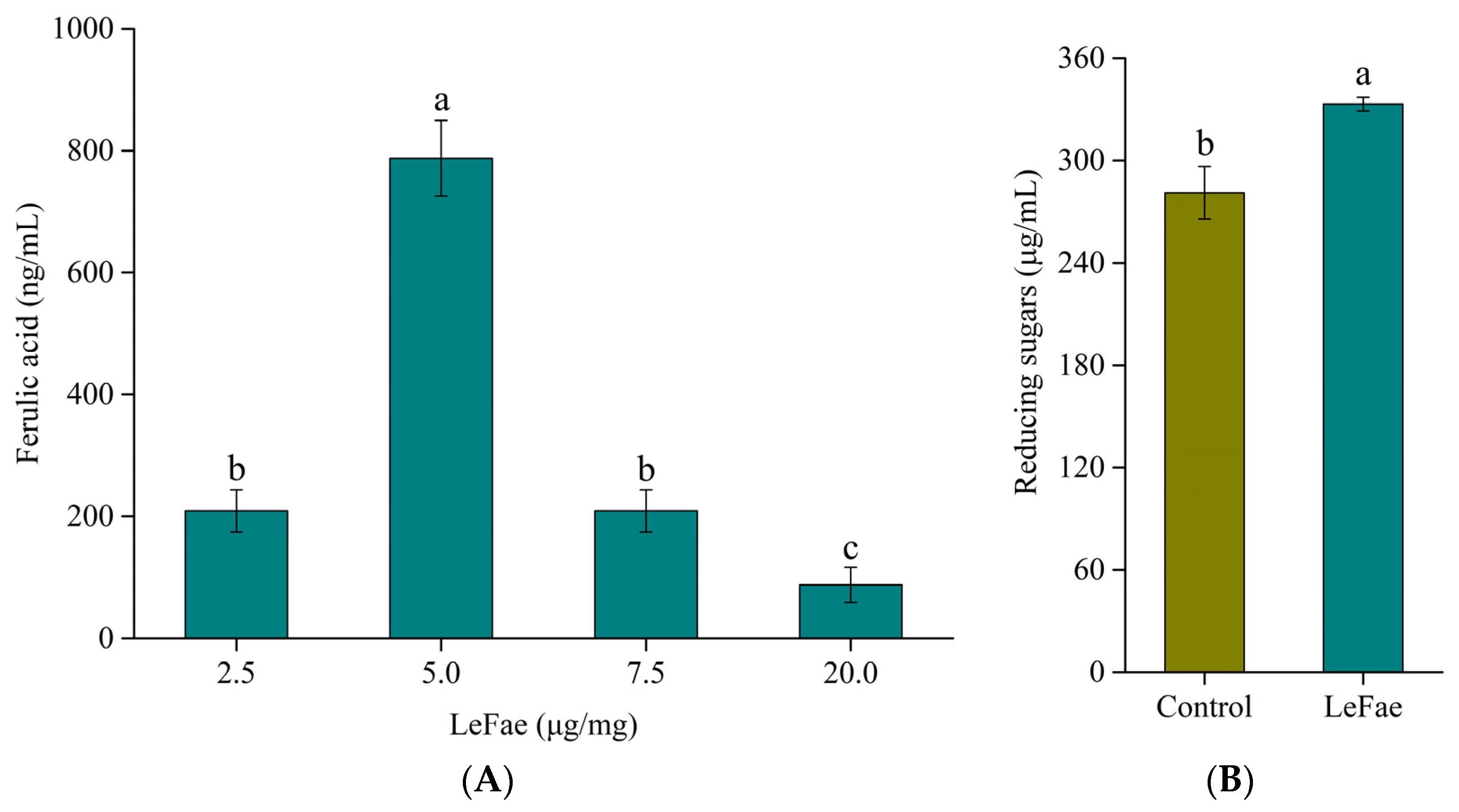
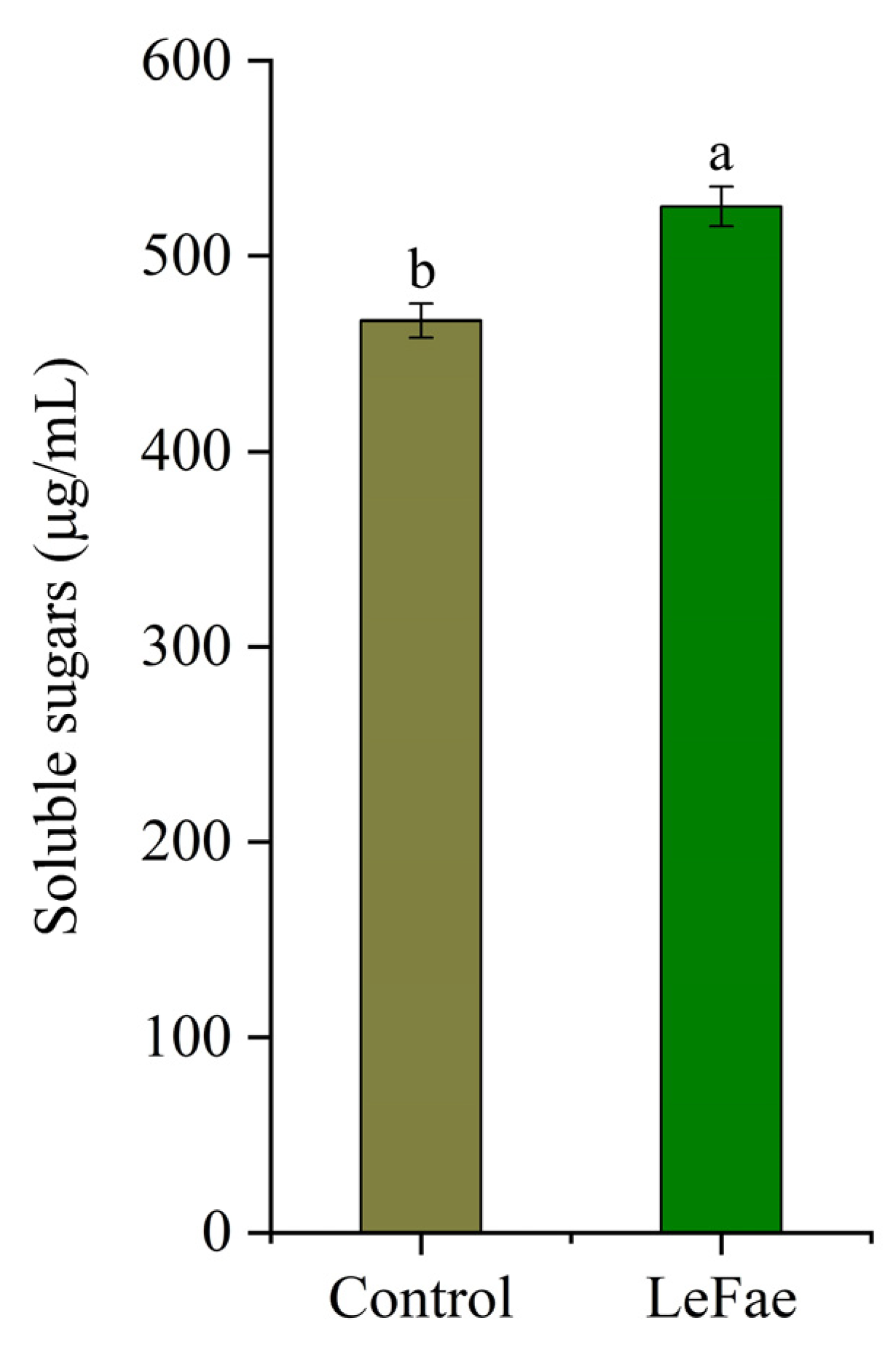
3.3. Hydrolysis of Wheat Straw Using LeFae
3.4. Synergism between LeFae and Cellulase
3.5. In Vitro Fermentation of Wheat Straw
3.6. In Vitro Fermentation of Wheat Straw Pretreated with LeFae
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tufail, T.; Saeed, F.; Afzaal, M.; Ain, H.B.U.; Gilani, S.A.; Hussain, M.; Anjum, F.M. Wheat straw: A natural remedy against different maladies. Food Sci. Nutr. 2021, 9, 2335–2344. [Google Scholar] [CrossRef]
- Togtokhbayar, N.; Cerrillo, M.A.; Rodríguez, G.B.; Elghandour, M.M.; Salem, A.Z.; Urankhaich, C.; Jigjidpurev, S.; Odongo, N.E.; Kholif, A.E.J.A.S.J. Effect of exogenous xylanase on rumen in vitro gas production and degradability of wheat straw. Anim. Sci. J. 2015, 86, 765–771. [Google Scholar] [CrossRef]
- Haddad, S.; Grant, R.; Kachman, S. Effect of wheat straw treated with alkali on ruminal function and lactational performance of dairy cows. J. Dairy Sci. 1998, 81, 1956–1965. [Google Scholar] [CrossRef]
- Jung, H.; Allen, M. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J. Anim. Sci. 1995, 73, 2774–2790. [Google Scholar] [CrossRef]
- Pan, G.X.; Bolton, J.L.; Leary, G.J. Determination of ferulic and p-coumaric acids in wheat straw and the amounts released by mild acid and alkaline peroxide treatment. J. Agric. Food Chem. 1998, 46, 5283–5288. [Google Scholar] [CrossRef]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jørgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic cross-links: Building and de-constructing the plant cell wall. Nat. Prod. Rep. 2020, 37, 919–961. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Mäkelä, M.R.; Aguilar-Pontes, M.V.; Benoit-Gelber, I.; Hildén, K.S.; de Vries, R.P. Diversity of fungal feruloyl esterases: Updated phylogenetic classification, properties, and industrial applications. Biotechnol. Biofuels 2016, 9, 231. [Google Scholar] [CrossRef] [Green Version]
- Li, J.S.; Lau, Y.Q.; Sun, T.Y.; Chen, C.S. Purification and biochemical characterization of an alkaline feruloyl esterase from Penicillium sumatrense NCH-S2 using rice bran as substrate. CyTA J. Food 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Mafa, M.S.; Malgas, S.; Pletschke, B.I. Feruloyl esterase (FAE-1) sourced from a termite hindgut and GH10 xylanases synergy improves degradation of arabinoxylan. AMB Express 2021, 11, 21. [Google Scholar] [CrossRef]
- Record, E.; Asther, M.; Sigoillot, C.; Pagès, S.; Punt, P.J.; Delattre, M.; Haon, M.; Hondel, C.A.M.J.J.V.D.; Sigoillot, J.-C.; Lesage-Meessen, L. Overproduction of the Aspergillus niger feruloyl esterase for pulp bleaching application. Appl. Microbiol. Biotechnol. 2003, 62, 349–355. [Google Scholar] [CrossRef]
- Li, L.; Qu, M.; Liu, C.; Xu, L.; Pan, K.; Song, X.; OuYang, K.; Li, Y.; Zhao, X. Expression of a Recombinant Lentinula edodes Xylanase by Pichia pastoris and Its Effects on Ruminal Fermentation and Microbial Community in in vitro Incubation of Agricultural Straws. Front. Microbiol. 2018, 9, 2944. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Nakade, K.; Sato, S.; Yoshida, K.; Miyazaki, K.; Natsume, S.; Konno, N. Lentinula edodes Genome Survey and Postharvest Transcriptome Analysis. Appl. Environ. Microbiol. 2017, 83, e02990-16. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.; Harrison, B.; Schneider, H.; MacKenzie, C.; Fontana, J. Xylan-hydrolysing enzymes from Streptomyces spp. Enzym. Microb. Technol. 1988, 10, 403–409. [Google Scholar] [CrossRef]
- Siew, C.K.; Williams, P.A. Role of Protein and Ferulic Acid in the Emulsification Properties of Sugar Beet Pectin. J. Agric. Food Chem. 2008, 56, 4164–4171. [Google Scholar] [CrossRef]
- Li, L.; Qu, M.; Liu, C.; Pan, K.; Xu, L.; OuYang, K.; Song, X.; Li, Y.; Zhao, X. Expression of a recombinant Lentinula edodes cellobiohydrolase by Pichia pastoris and its effects on in vitro ruminal fermentation of agricultural straws. Int. J. Biol. Macromol. 2019, 134, 146–155. [Google Scholar] [CrossRef]
- Hughes, M.; Mlambo, V.; Lallo, C.H.O.; Jennings, P.G.A. Potency of microbial inocula from bovine faeces and rumen fluid for in vitro digestion of different tropical forage substrates. Grass Forage Sci. 2012, 67, 263–273. [Google Scholar] [CrossRef]
- Zhao, X.H.; Liu, C.J.; Liu, Y.; Li, C.Y.; Yao, J.H. Effects of replacing dietary starch with neutral detergent-soluble fibre on ruminal fermentation, microbial synthesis and populations of ruminal cellulolytic bacteria using the rumen simulation technique (RUSITEC). J. Anim. Physiol. Anim. Nutr. 2012, 97, 1161–1169. [Google Scholar] [CrossRef]
- Makkar, H.; Sharma, O.; Dawra, R.; Negi, S. Simple Determination of Microbial Protein in Rumen Liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef]
- Chen, L.-Y.; Cheng, C.-W.; Liang, J.-Y. Effect of esterification condensation on the Folin–Ciocalteu method for the quantitative measurement of total phenols. Food Chem. 2015, 170, 10–15. [Google Scholar] [CrossRef]
- Nielsen, S.S. Total Carbohydrate by Phenol-Sulfuric Acid Method. In Food Analysis Laboratory Manual. Food Science Text Series; Springer: Cham, Switzerland, 2017; pp. 137–141. [Google Scholar] [CrossRef]
- Dimarogona, M.; Topakas, E.; Christakopoulos, P.; Chrysina, E.D. The crystal structure of a Fusarium oxysporum feruloyl esterase that belongs to the tannase family. FEBS Lett. 2020, 594, 1738–1749. [Google Scholar] [CrossRef]
- Suzuki, K.; Hori, A.; Kawamoto, K.; Thangudu, R.R.; Ishida, T.; Igarashi, K.; Samejima, M.; Yamada, C.; Arakawa, T.; Wakagi, T.; et al. Crystal structure of a feruloyl esterase belonging to the tannase family: A disulfide bond near a catalytic triad. Proteins Struct. Funct. Bioinform. 2014, 82, 2857–2867. [Google Scholar] [CrossRef]
- Li, L.; Shaw, P.E. A STAT3 dimer formed by inter-chain disulphide bridging during oxidative stress. Biochem. Biophys. Res. Commun. 2004, 322, 1005–1011. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Liu, Y.; Leng, J. Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing. Appl. Sci. 2021, 11, 4453. [Google Scholar] [CrossRef]
- Kanauchi, M.; Watanabe, S.; Tsukada, T.; Atta, K.; Kakuta, T.; Koizumi, T. Purification and Characteristics of Feruloyl Esterase from Aspergillus awamori G-2 Strain. J. Food Sci. 2008, 73, C458–C463. [Google Scholar] [CrossRef]
- Donaghy, J.; McKay, A.M. Purification and characterization of a feruloyl esterase from the fungus Penicilliumexpansum. J. Appl. Microbiol. 1997, 83, 718–726. [Google Scholar] [CrossRef]
- Hemida, S.K.; Omar, S.A.; Abdel-Mallek, A.Y. Microbial populations and enzyme activity in soil treated with heavy metals. Water Air Soil Pollut. 1997, 95, 13–22. [Google Scholar] [CrossRef]
- Esteban-Torres, M.; Reverón, I.; Mancheño, J.M.; Rivas, B.d.L.; Muñoz, R. Characterization of a Feruloyl Esterase from Lactobacillus plantarum. Appl. Environ. Microbiol. 2013, 79, 5130–5136. [Google Scholar] [CrossRef] [Green Version]
- Akira, Y.; Hiroshi, M.; Yutaka, T.; Naomi, W.; Akira, M.; Hajime, N.; Takashi, I.; Kazue, O.; Junji, Y. Lymphocyte transformation and thiol compounds: The role of ADF/thioredoxin as an endogenous reducing agent. Mol. Immunol. 1992, 29, 263–270. [Google Scholar] [CrossRef]
- Fu, Z.; Zhu, Y.; Teng, C.; Fan, G.; Li, X. Biochemical characterization of a novel feruloyl esterase from Burkholderia pyrrocinia B1213 and its application for hydrolyzing wheat bran. 3 Biotech 2021, 12, 24. [Google Scholar] [CrossRef]
- Fazary, A.E.; Ismadji, S.; Ju, Y.-H. Biochemical studies on native and cross-linked aggregates of Aspergillus awamori feruloyl esterase. Int. J. Biol. Macromol. 2009, 44, 240–248. [Google Scholar] [CrossRef]
- Cheng, F.; Sheng, J.; Cai, T.; Jin, J.; Liu, W.; Lin, Y.; Du, Y.; Zhang, M.; Shen, L. A Protease-Insensitive Feruloyl Esterase from China Holstein Cow Rumen Metagenomic Library: Expression, Characterization, and Utilization in Ferulic Acid Release from Wheat Straw. J. Agric. Food Chem. 2012, 60, 2546–2553. [Google Scholar] [CrossRef]
- Ghazali, N.F.; Makhtar, N.A. Enzymatic hydrolysis of oil palm empty fruit bunch and its kinetics. Malays. J. Anal. Sci. 2018, 22, 715–722. [Google Scholar] [CrossRef]
- Martin, S. Effects of p-coumaric acid and ferulic acid on methane production and fibre digestion by mixed rumen micro-organisms in vitro. Lett. Appl. Microbiol. 1988, 7, 113–114. [Google Scholar] [CrossRef]
- Chesson, A.; Stewart, C.S.; Wallace, R.J. Influence of Plant Phenolic Acids on Growth and Cellulolytic Activity of Rumen Bacteria. Appl. Environ. Microbiol. 1982, 44, 597–603. [Google Scholar] [CrossRef]
- Akin, D.; Rigsby, L.; Theodorou, M.; Hartley, R. Population changes of fibrolytic rumen bacteria in the presence of phenolic acids and plant extracts. Anim. Feed. Sci. Technol. 1988, 19, 261–275. [Google Scholar] [CrossRef]
- Soberon, M.; Cherney, D.; Cherney, J. Free ferulic acid uptake in ram lambs1. J. Anim. Sci. 2012, 90, 1885–1891. [Google Scholar] [CrossRef]
- Marvin, H.J.P.; Krechting, C.F.; Van Loo, E.N.; Snijders, C.H.A.; Lommen, A.; Dolstra, O. Relationship between Phenolic Acids Formed During Rumen Degradation of Maize Samples and in vitro Digestibility. J. Sci. Food Agric. 1996, 71, 111–118. [Google Scholar] [CrossRef]
- Kanelias, K.; Mould, F. The effect of ferulic acid esterase on the in vitro degradability of wheat straw. Proc. Br. Soc. Anim. Sci. 2006, 189. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Wang, W.-K.; Wu, Q.-C.; Yang, H.-J. The release and catabolism of ferulic acid in plant cell wall by rumen microbes: A review. Anim. Nutr. 2022, 9, 335–344. [Google Scholar] [CrossRef]
- Gu, S.; Liu, C.; Zhang, W.; Qu, M.; Li, Y.; Zang, Y.; Xiong, X.; Pan, K.; Zhao, X. Characteristics of a recombinant Fusarium verticillioides cutinase and its effects on enzymatic hydrolysis of rice straw. Int. J. Biol. Macromol. 2021, 171, 382–388. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Control | LeFae | p |
|---|---|---|---|
| IVDMD (%) | 31.7 ± 1.46 | 22.1 ± 3.46 | 0.01 |
| pH | 5.8 ± 0.05 | 6.1 ± 0.04 | <0.01 |
| Total VFA (mM) | 71.4 ± 1.93 | 61.5 ± 1.45 | <0.01 |
| Acetate (mM) | 42.7 ± 0.78 | 35.2 ± 0.73 | <0.01 |
| Propionate (mM) | 14.0 ± 0.12 | 11.1 ± 0.30 | <0.01 |
| Butyrate (mM) | 5.9 ± 0.67 | 5.5 ± 0.12 | 0.38 |
| Isobutyrate (mM) | 1.4 ± 0.25 | 1.1 ± 0.06 | 0.08 |
| Valerate (mM) | 1.3 ± 0.18 | 1.5 ± 0.32 | 0.59 |
| Isovalerate (mM) | 6.2 ± 0.24 | 7.2 ± 0.46 | 0.03 |
| Ammonia-N (mM) | 6.4 ± 0.16 | 7.1 ± 0.11 | 0.03 |
| FLMCP (µg/mL) | 262.4 ± 1.96 | 253.6 ± 24.72 | 0.57 |
| Ferulic acid (ng/mL) | 35.0 ± 3.89 | 63.2 ± 11.2 | 0.02 |
| Soluble phenols (μg-GAE/mL) | 24.3 ± 0.46 | 26.7 ± 1.85 | 0.10 |
| 16S rDNA gene copy numbers of microorganisms (log10N/mL) | |||
| Total bacteria | 9.79 ± 0.02 | 9.82 ± 0.04 | 0.35 |
| R. flavefaciens | 7.58 ± 0.32 | 4.54 ± 0.27 | <0.01 |
| F. succinogenes | 4.58 ± 0.17 | 4.65 ± 0.15 | 0.59 |
| R. albus | 5.65 ± 0.35 | 5.22 ± 0.37 | 0.21 |
| Item | Control | LeFae | p |
|---|---|---|---|
| IVDMD (%) | 30.3 ± 1.94 | 27.0 ± 2.40 | 0.14 |
| pH | 7.3 ± 0.074 | 7.6 ± 0.22 | 0.11 |
| Total VFA (mM) | 42.0 ± 1.87 | 41.3 ± 0.37 | 0.56 |
| Ammonia-N (mM) | 4.4 ± 0.17 | 4.3 ± 0.29 | 0.77 |
| FLMCP (µg/mL) | 150.3 ± 1.67 | 142.0 ± 13.60 | 0.40 |
| Soluble phenols (μg-GAE/mL) | 29.6 ± 0.08 | 29.8 ± 0.29 | 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Lei, X.; Ouyang, K.; Zhang, W.; Liu, C.; Li, Y.; Qiu, Q.; Zang, Y.; Qu, M.; Pan, K.; et al. Characteristics of a Recombinant Lentinula edodes Ferulic Acid Esterase and Its Adverse Effects on In Vitro Fermentation of Wheat Straw. Fermentation 2023, 9, 683. https://doi.org/10.3390/fermentation9070683
Zhang X, Lei X, Ouyang K, Zhang W, Liu C, Li Y, Qiu Q, Zang Y, Qu M, Pan K, et al. Characteristics of a Recombinant Lentinula edodes Ferulic Acid Esterase and Its Adverse Effects on In Vitro Fermentation of Wheat Straw. Fermentation. 2023; 9(7):683. https://doi.org/10.3390/fermentation9070683
Chicago/Turabian StyleZhang, Xiangyu, Xiaowen Lei, Kehui Ouyang, Wenjing Zhang, Chanjuan Liu, Yanjiao Li, Qinghua Qiu, Yitian Zang, Mingren Qu, Ke Pan, and et al. 2023. "Characteristics of a Recombinant Lentinula edodes Ferulic Acid Esterase and Its Adverse Effects on In Vitro Fermentation of Wheat Straw" Fermentation 9, no. 7: 683. https://doi.org/10.3390/fermentation9070683
APA StyleZhang, X., Lei, X., Ouyang, K., Zhang, W., Liu, C., Li, Y., Qiu, Q., Zang, Y., Qu, M., Pan, K., & Zhao, X. (2023). Characteristics of a Recombinant Lentinula edodes Ferulic Acid Esterase and Its Adverse Effects on In Vitro Fermentation of Wheat Straw. Fermentation, 9(7), 683. https://doi.org/10.3390/fermentation9070683