Molecular Characterization of Terpenoid Biosynthetic Genes and Terpenoid Accumulation in Phlomis umbrosa Turczaninow

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Total RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.3. Next Generation Sequencing of Transcriptome
2.4. Sequence Analysis
2.5. Isolation and HPLC Analysis of P. umbrosa Terpenoids
2.6. Statistical Analysis
3. Results
3.1. Sequencing and Transcriptome Assembly
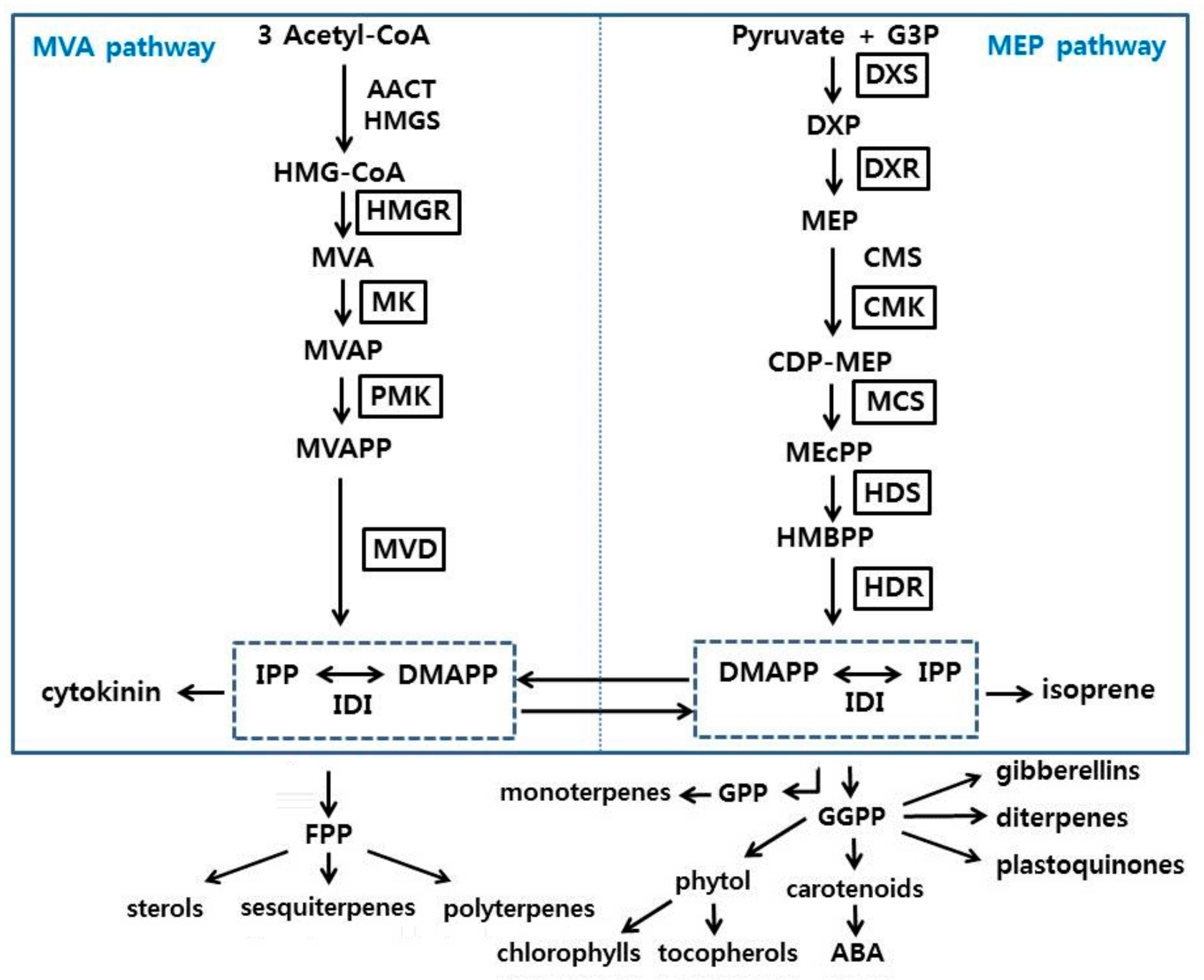
3.2. Isolation and Sequence Analysis of Terpenoid Biosynthetic Genes from P. umbrosa
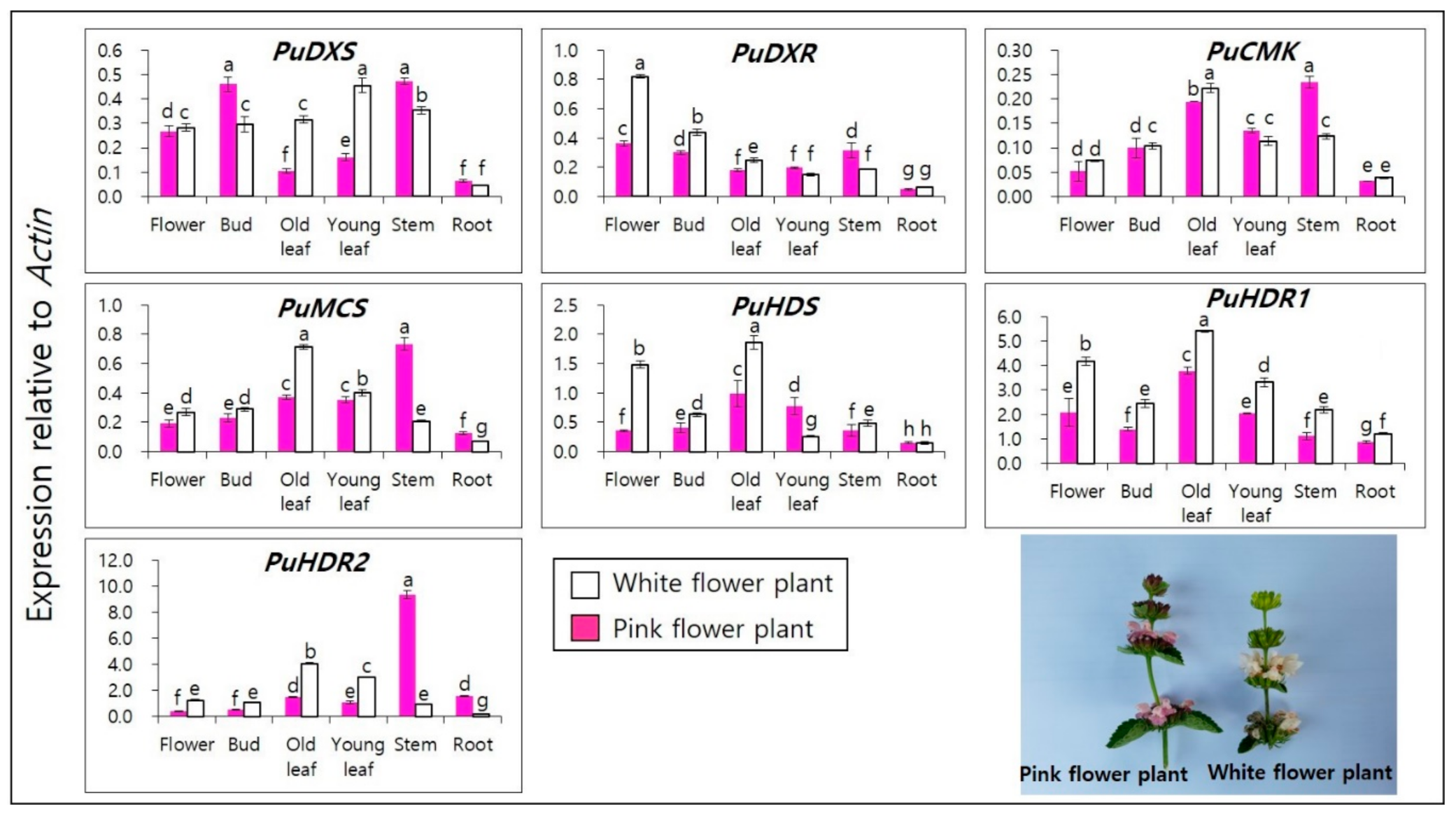
3.3. Terpenoid Biosynthetic Genes Spatial Expression Profiles in of P. umbrosa
3.4. Terpenoid (Iridoid Glycosides) Analysis in Different Organs of P. umbrosa by HPLC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Wang, Z.Z. Comparative analysis of essential oil componets of three Phlomis species in Qinling Mountains of China. J. Pharm. Miomed. Anal. 2008, 47, 213–217. [Google Scholar] [CrossRef]
- Liu, P.; Deng, R.X.; Duan, H.Q.; Yin, W.P.; Zhao, T.Z. Phenylethanoid glycosides from the roots of Phlomis umbrosa. J. Asian Nat. Prod. Res. 2009, 11, 69–74. [Google Scholar] [CrossRef]
- Lee, D.; Kim, Y.S.; Song, J.; Kim, H.S.; Lee, H.J.; Guo, H.; Kim, H. Effects of Phlomis umbrosa root on longitudinal bone growth rate in adolescent female rats. Molecules 2016, 21, 461. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.M.; Yan, F.L.; Tan, J.; Bai, Y.X.; Wang, X.; Yang, Y.X. Two new dammarane-type glycosides from Phlomis umbrosa. Nat. Prod. Res. 2014, 28, 18–23. [Google Scholar] [CrossRef]
- Shin, T.Y.; Kim, S.H.; Kim, D.K.; Leem, K.H.; Park, J.S. Phlomis umbrosa root inhibits mast cell-dependent allergic reactions and inflammatory cytokine secretion. Phytother. Res. 2008, 22, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Wang, J.; Li, M.; Miao, X.; Pan, H.; Yang, Y.; Wang, Y. Anti-nociceptive and anti-inflammatory activities of phlomis umbrosa turcz extract. Fitoterapia 2011, 82, 716–721. [Google Scholar] [CrossRef]
- Breitmaier, E. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones. In Terpene: Importance, General Structure, and Biosynthesis; Wilely–VCH: Weinheim, Germany, 2006; pp. 2–3. [Google Scholar]
- Inouye, H.; Takeda, Y.; Nishmura, H.; Kanomi, A.; Okuda, T.; Puff, C. Chemotaxonomic studies of Rubiaceous plants containing iridoid glycosieds. Phytochemistry 1988, 27, 2591–2598. [Google Scholar] [CrossRef]
- Alpieva, K.I.; Taskova, R.M.; Evstatieva, L.N.; Handjieva, N.V.; Popov, S.S. Benzoxazinoids and iridoid glucosides from four Laminum species. Phytochemistry 2003, 64, 1413–1417. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Guo, L.; Cao, L.J.; Zhang, J.; Yin, Z.Q. A new iridoid glycoside from the fruits of Vitex rotundifolia. Nat. Prod. Res. 2017, 31, 2491–2496. [Google Scholar] [CrossRef]
- Fan, P.C.; Ma, H.P.; Hao, Y.; He, X.R.; Sun, A.J.; Jiang, W.; Li, M.X.; Jing, L.L.; He, L.; Ma, J.; et al. A new anti-fibrinolytic hemostatic compound 8-O-acetyl shanzhiside methylester extracted from Lamiopholomis rotata. J. Ethnopharmacol. 2016, 187, 232–238. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, Y.; Zhao, D.; Feng, X.; Zhang, L.; Liu, C. Loganin prevents BV-2 microglia cells from Aβ1-42-induced inflammatiion via regulating TLR4/TRAF-κB axis. Cell Biol. Int. 2018, 42, 1632–1642. [Google Scholar] [CrossRef]
- Czerwinska, M.E.; Swierczewska, A.; Granica, S. Bioactive constituents of Lamium album L. as inhibitors of cytokine secretion in human neutrophils. Molecules 2018, 23, 2770. [Google Scholar] [CrossRef] [Green Version]
- Webb, H.; Foley, W.J.; Külheim, C. The genetic basis of foliar terpene yield: Implications for breeding and profitability of Australian essential oil crops. Plant. Biotechnol. 2014, 31, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.J.; Zhan, J.Y.; Zheng, T.R.; Sun, R.; Wang, T.; Tang, Z.Z.; Bu, T.L.; Li, C.L.; Wu, Q.; Chen, H. The jasmonate-responsive transcription factor CbWRK24 regulates terpenoid biosynthetic genes to promote sapnin biosynthesis in Conyza blinii H. Léb. J. Genet. 2018, 97, 1379–1388. [Google Scholar] [CrossRef]
- Yu, N.; Yang, J.C.; Yin, G.T.; Li, R.S.; Zou, W.T. Transcriptome analysis of oleoresin-producing tree Sindora Glabra and characterizationof sesquiterpene synthases. Front. Plant Sci. 2018, 9, 1619. [Google Scholar] [CrossRef]
- Zhang, H.; Jin, B.; Bu, J.; Guo, J.; Chen, T.; Ma, Y.; Tang, J.; Cui, G.; Huang, L. Transcriptomic insight into terpenoid biosynthesis and functional characterizaion of three diterpene synthases in Scutellaria barbata. Molecules 2018, 23, 2952. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Hussain, R.M.; Rehman, N.U.; She, G.; Li, P.; Wan, X.; Guo, L.; Zhao, J. De novo transcriptome sequencing and metabolite profiling analysis reveal the comples metabolic genes involved in the terpenoid biosynthesis in blue anise sage (Salvia guaranitica L.). DNA Res. 2018, 25, 597–617. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, S.S.; Kim, Y.B.; Lee, S.W.; Park, S.U. Expression of terpenoid biosynthetic genes and accumulation of chemical constituents in Valeriana fauriei. Molecules 2016, 21, 691. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Wei, L.; Fan, P.; Zhang, Q.; Jia, Z. Determination of five iridoid glycosides in Phlomis younghusbandii by HPLC. China J. Chin. Mater. Med. 2011, 36, 594–597. [Google Scholar]
- Sprenger, G.A.; Schörken, U.; Wiegert, T.; Grolle, S.; de Graaf, A.A.; Taylor, S.V.; Begley, T.P.; Bringer-Meyer, S.; Sahm, H. Identification of a thiamin-dependent synthase in Escherichia coli required for the formation of the 1-deoxy-D-xylulose 5-phosphate precursor to isoprenoids, thiamin, and pyridoxol. Proc. Natl. Acad. Sci. USA 1997, 94, 12857–12862. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Gong, Y.; Zuo, K.; Ling, H.; Qiu, C.; Zhang, F.; Wang, Y.; Pi, Y.; Liu, X.; Sun, X.; et al. Molecular cloning, expression profiling and functional analysis of DXR gene encoding 1-deoxy-D-Xylulose 5-phosphate reductoisomerase from Camptotheca acuminata. J. Plant Physiol. 2008, 165, 203–213. [Google Scholar] [CrossRef]
- Rohdich, F.; Wungsintaweekul, J.; Lüttgen, H.; Fischer, M.; Eisenreich, W.; Schuhr, C.A.; Fellermeier, M.; Schramek, N.; Zenk, M.H.; Bacher, A. Biosynthesis of terpenoids: 4-Diphosphocytidyl-2-C-methyl-D-erythritol kinase from tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 8251–8256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Kuzuyama, T.; Chang, Y.J.; Kim, S.U. Cloning and characterizatiion of 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MECS) gene from Ginkgo biloba. Plant Cell Rep. 2006, 25, 829–833. [Google Scholar] [CrossRef]
- Wolff, M.; Seemann, M.; Grosdemange-Billiard, C.; Tritsch, D.; Campos, N.; Rodriguez-Concepción, M.; Boronat, A.; Rohmer, M. Isoprenoid biosynthesis via the methylerythritol phosphate pathway. (E)-4-Hydroxy-3-methylbut-2-enyl diphosphate: Chemical synthesis and formation from methylerythritol cyclodiphosphate by a cell-free sstem from Escherichia coli. Tetrahedron Lett. 2002, 43, 2555–2559. [Google Scholar] [CrossRef]
- Hsieh, W.Y.; Hsieh, M.H. The amino-terminal conserved domain of 4-hydroxy-3-methylbut-2enyl diphosphate reductase is critical for its function in oxygen-evolving photosynthetic organisms. Plant Signal. Behav. 2015, 10, e988072. [Google Scholar] [CrossRef] [Green Version]
- Istvan, E.S. Bacterial and mamalian HMG-CoA reductases: Related enzymes with distinct architectures. Curr. Opin. Struc. Biol. 2001, 11, 746–751. [Google Scholar] [CrossRef]
- Andeassi, J.L., 2nd.; Bilder, P.W.; Vetting, M.W.; Roderick, S.L.; Leyh, T.S. Crystal structure of the streptococcus pneumoniae mevalonate kinase in complex with diphosphomevalonate. Protein Sci. 2007, 16, 983–989. [Google Scholar] [CrossRef] [Green Version]
- Doun, S.S.; Burgner, J.W., 2nd.; Briggs, S.D.; Rodwell, V.W. Enterococcus faecalis phosphomevalonate kinase. Protein Sci. 2005, 14, 1134–1139. [Google Scholar] [CrossRef] [Green Version]
- Cordier, H.; Lacombe, C.; Karst, F.; Bergès, T. The Saccharomyces cerevisiae mevalonate diphosphate decarboxylase (Erg19p) forms homodimers in vivo, and a single substitution in a structurally conserved region impairs dimerization. Curr. Microbiol. 1999, 38, 290–294. [Google Scholar] [CrossRef]
- Xu, C.; Li, H.; Yang, X.; Gu, G.; Mu, H.; Yue, Y.; Wang, L. Cloning and expression analysis of MEP pathway enzyme-encoding genes in Osmanthus fragrans. Genes 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Liu, G.S.; Hussian, Z.; Sun, Y.H. Cloning and expression analysis of terpenoid metabolism-related gene NsyCMS in Nicotiana sylvestris. Genet. Mol. Res. 2015, 14, 3300–3308. [Google Scholar] [CrossRef]
- Souret, F.F.; Kim, Y.; Wyslouzil, B.E.; Wobbe, K.K.; Weathers, P.J. Scale-up of Artemisia annua L. hairy root cultures produces complex patterns of terpenoid gene expression. Biotechnol. Bioeng. 2003, 83, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Cui, G.; Zhou, S.F.; Zhang, X.; Huang, L. Cloning and characterization of a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase gene from Salvia miltiorrhiza involved in diterpenoid tanshinone accumulation. J. Plant Physiol. 2011, 168, 148–157. [Google Scholar] [CrossRef]
- Xu, J.W.; Xu, Y.N.; Zhong, J.J. Enhancement of ganoderic acid accumulation by overexpression of an N-terminally truncated 3-hydroxy-3-methylglutaryl coenzyme A reductase gene in basidomycete Ganoderma lucidum. J. Appl. Environ. Microbiol. 2012, 78, 7968–7976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.M.; Chiang, A.; Lund, S.T.; Bohlmann, J. Biosynthesis of wine aroma; transcript profiles of hydroxymethybutenyl diphosphate reductase, geranyl diphosphate synthase, and linalool/nerolidol synthase parallel monoterpenol glycoside accumulation in gewürztraminer grapes. Planta 2012, 236, 919–929. [Google Scholar] [CrossRef]
- Fuji, Y.; Uchida, A.; Fukahori, K.; Chino, M.; Ohtsuki, T.; Matsufuji, H. Chemical characterization and biological activity in young sesame leaves (Sesamum indicum L.) and changes in iridoid and polyphenol content at different growth stages. PLoS ONE 2018, 13, e0194449. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ → 3′) | Size (bp) | GenBank Access. No. |
|---|---|---|---|
| PuDXS (F) PuDXS (R) | ACGGGCATGAACCTCTTCCA GATGCCTTCACAGGCCAACC | 108 | KU317508 |
| PuDXR (F) PuDXR (R) | TCCAAGGTGGATTGCCTTTGAA GTTTCGGGCCATCCCAAGTT | 147 | KU317509 |
| PuCMK (F) PuCMK (R) | CTGCTACGCTCCCAACAACG AGCGGTGGCTTTGATCGAATA | 148 | MK482372 |
| PuMCS (F) PuMCS (R) | GCTGCTGCCTCAAGCACTGT GAGCCGGTGAAGGTCGAATC | 105 | MK482371 |
| PuHDS (F) PuHDS (R) | GGTCTCAAAACCAGGGATCA TAGCAATGGGCTTCCTTCAG | 180 | MK482373 |
| PuHDR1 (F) PuHDR1 (R) | GGCCATCTCTCTGCAATTCT TTGCTCCTCGTCAAGTTGTG | 180 | MK482374 |
| PuHDR2 (F) PuHDR2 (R) | ATGGCGATTTCTCTGCAGTT CCCCTTGCGATTGTAGTTGT | 207 | MK482375 |
| PuHMGR1 (F) PuHMGR1 (R) | CGCCTACTTTCTGCTTCACC ATGTTCCGATCTTCGTGGAG | 216 | KU317502 |
| PuHMGR2 (F) PuHMGR2 (R) | CACCCCTCTCCACATCCTTA GCGTGTATCTTCGTTCAGCA | 190 | KU317501 |
| PuMK (F) PuMK (R) | CCTACTCCCGAGGAAAATGAT TGGCCTCGAATGAACATGATA | 151 | KU317505 |
| PuPMK (F) PuPMK (R) | CCAAATGCCGGGATTGTACTG TGGCCATCTGAGGAGAAGTGAG | 129 | KU317506 |
| PuMVD (F) PuMVD (R) | GCGATAGCATTGCTGTTCAA ATCGCGCTTTTGTATTGCTT | 209 | KU317504 |
| PuActin (F) PuActin (R) | TCCAGCCTTCATTGATCGGAA GGTCAGCGATACCAGGGAACA | 150 | KU317507 |
| Total Number | Total Nucleotides (nt) | Mean Length (nt) | N50 | |
|---|---|---|---|---|
| Total raw reads | 21,281,712 | - | - | - |
| Total Contigs | 149,488 | 184,148,836 | 1231 | 1935 |
| Total unigenes | 75,392 | 64,062,377 | 849 | 1422 |
| Type | Organ | Sesamoside | Shanzhiside Methylester | Umbroside | Acteoside |
|---|---|---|---|---|---|
| White flower plant | Flower | 40.68 ± 0.10h | 1.10 ± 0.01h | 0.70 ± 0.00i | 18.58 ± 0.04c |
| Bud | 49.94 ± 0.35e | 1.58 ± 0.04f | 1.30 ± 0.02h | 62.70 ± 0.64a | |
| Young leaf | 45.50 ± 0.01f | 0.49 ± 0.01j | 11.37 ± 0.10d | 6.48 ± 0.83d | |
| Old leaf | 36.75 ± 0.27i | 1.72 ± 0.01e | 10.79 ± 0.04f | 1.62 ± 0.35ef | |
| Stem | 11.60 ± 0.12k | 0.65 ± 0.01i | 0.63 ± 0.00i | 0.20 ± 0.00g | |
| Root | 133.78 ± 1.38a | 16.12 ± 0.17a | 11.07 ± 0.13e | 0.81 ± 0.01fg | |
| Pink flower plant | Flower | 59.64 ± 0.72c | 2.37 ± 0.08c | 1.15 ± 0.02h | 17.92 ± 0.25c |
| Bud | 54.58 ± 0.79d | 1.43 ± 0.08g | 2.00 ± 0.03g | 44.14 ± 0.92b | |
| Young leaf | 43.08 ± 0.23g | 0.53 ± 0.01j | 25.73 ± 0.10a | 5.70 ± 0.78d | |
| Old leaf | 36.43 ± 0.41i | 1.91 ± 0.05d | 18.47 ± 0.25b | 2.24 ± 0.24e | |
| Stem | 16.03 ± 0.23j | 0.71 ± 0.01i | 0.31 ± 0.00j | 0.40 ± 0.01g | |
| Root | 116.10 ± 0.48b | 3.00 ± 0.01b | 12.91 ± 0.04c | 1.46 ± 0.01ef |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-Y.; Lee, S.-H.; Lee, Y.-S.; Han, S.-H.; Song, B.-H.; Reddy, C.S.; Kim, Y.B.; Park, S.U. Molecular Characterization of Terpenoid Biosynthetic Genes and Terpenoid Accumulation in Phlomis umbrosa Turczaninow. Horticulturae 2020, 6, 76. https://doi.org/10.3390/horticulturae6040076
Hwang S-Y, Lee S-H, Lee Y-S, Han S-H, Song B-H, Reddy CS, Kim YB, Park SU. Molecular Characterization of Terpenoid Biosynthetic Genes and Terpenoid Accumulation in Phlomis umbrosa Turczaninow. Horticulturae. 2020; 6(4):76. https://doi.org/10.3390/horticulturae6040076
Chicago/Turabian StyleHwang, Si-Yoon, Sang-Hoon Lee, Young-Seob Lee, Sin-Hee Han, Beom-Heon Song, Chinreddy Subramanyam Reddy, Yeon Bok Kim, and Sang Un Park. 2020. "Molecular Characterization of Terpenoid Biosynthetic Genes and Terpenoid Accumulation in Phlomis umbrosa Turczaninow" Horticulturae 6, no. 4: 76. https://doi.org/10.3390/horticulturae6040076
APA StyleHwang, S. -Y., Lee, S. -H., Lee, Y. -S., Han, S. -H., Song, B. -H., Reddy, C. S., Kim, Y. B., & Park, S. U. (2020). Molecular Characterization of Terpenoid Biosynthetic Genes and Terpenoid Accumulation in Phlomis umbrosa Turczaninow. Horticulturae, 6(4), 76. https://doi.org/10.3390/horticulturae6040076