Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota


Abstract
:1. Introduction
2. Materials and Methods
2.1. Differential Thermal Analysis
2.2. Winter Injury
2.3. Vineyard Re-Establishment
2.4. Statistical Analysis
3. Results
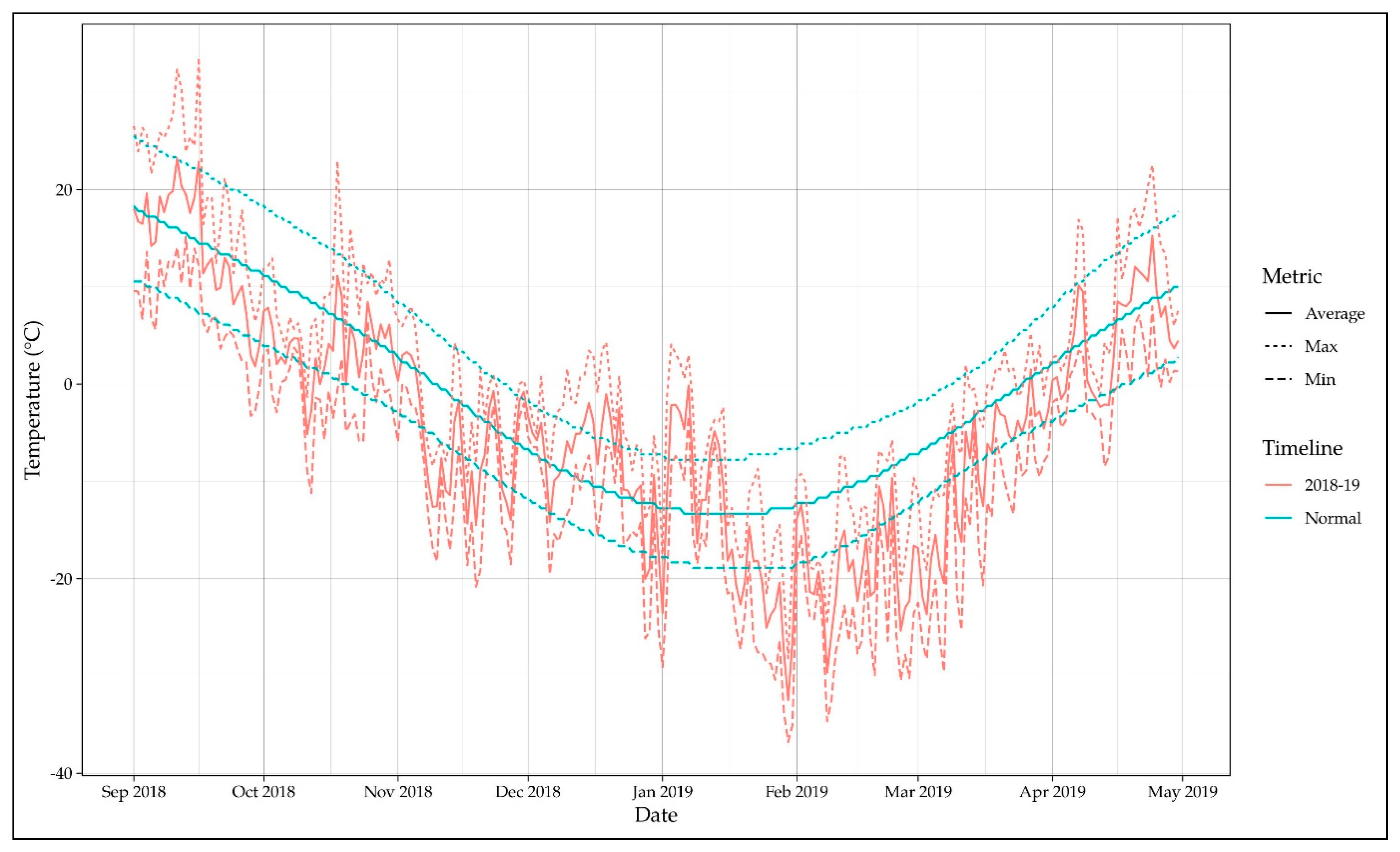
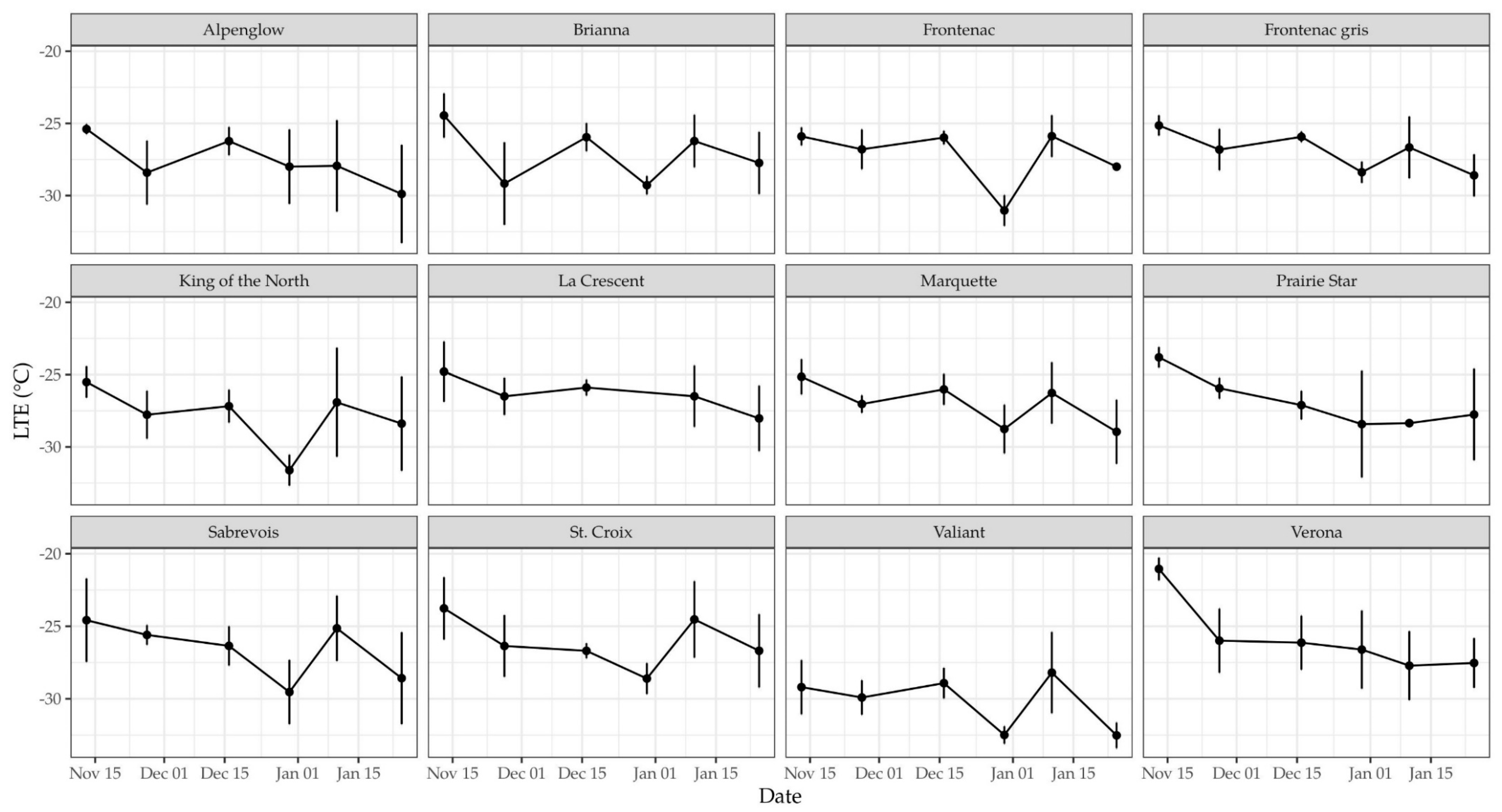
3.1. Differential Thermal Analysis of Dormant Buds
3.2. Winter Injury
3.3. Vineyard Re-Establishment
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Percent Trunks Failing to Break Bud | Percent Trunks Collapsed Midseason | Percent Surviving Trunks |
|---|---|---|---|
| DM 8521 | 0 | 100 | 0 |
| E.S. 3-20-33 | 100 | 0 | 0 |
| E.S. 4-22-60 | 100 | 0 | 0 |
| E.S. 9-4-72 | 50 | 17 | 33 |
| E.S. 10-18-14 | 100 | 0 | 0 |
| E.S. 10-18-58 | 0 | 67 | 33 |
| E.S. 10-18-75 | 100 | 0 | 0 |
| JB 13 | 33 | 33 | 33 |
| LxT 10 | 67 | 33 | 0 |
| Genotype | Retained Suckers (No.) | Live Nodes (No./Sucker) | Live Sucker Length (cm/Sucker) | Internode Length (cm) | Sucker Circumference (mm) | Sucker Caliper Ratio |
|---|---|---|---|---|---|---|
| DM 8521 | 1.0 | 30.0 | 322.6 | 10.7 | 37.9 | 1.21 |
| E.S. 3-20-33 | 2.0 | 12.8 | 137.8 | 10.8 | 19.0 | 1.21 |
| E.S. 4-22-60 | 3.5 | 21.1 | 238.0 | 10.7 | 32.6 | 1.09 |
| E.S. 9-4-72 | 1.5 | 27.7 | 408.9 | 14.6 | 32.4 | 1.06 |
| E.S. 10-18-14 | 3.0 | 19.0 | 210.0 | 10.4 | 23.1 | 1.12 |
| E.S. 10-18-58 | 2.0 | 21.3 | 269.9 | 11.5 | 29.3 | 1.10 |
| E.S. 10-18-75 | 2.0 | 29.5 | 368.9 | 12.3 | 30.1 | 1.19 |
| JB 13 | 3.0 | 24.0 | 256.5 | 10.7 | 25.0 | 1.12 |
| LxT 10 | 3.0 | 13.5 | 139.3 | 8.7 | 18.0 | 1.18 |
References
- Pierquet, P.; Stushnoff, C. Relationship of low temperature exotherms to cold injury in Vitis riparia Michx. Am. J. Enol. Vitic. 1980, 31, 1–6. [Google Scholar]
- Goffinet, C.M. Anatomy of Grapevine Winter Injury and Recovery. Available online: http://www.hort.cornell.edu/goffinet/Anatomy_of_Winter_Injury_hi_res.pdf (accessed on 11 December 2019).
- Londo, J.P.; Kovaleski, A.P. Characterization of wild North American grapevine cold hardiness using differential thermal analysis. Am. J. Enol. Vitic. 2017, 68, 203–212. [Google Scholar] [CrossRef]
- Howell, G. Grape cold hardiness: Mechanisms of cold acclimation, mid-winter hardiness maintenance, and spring deacclimation. In Proceedings of the ASEV Annual Meeting, Seattle, WA, USA, 19–23 June 2000; pp. 35–45. [Google Scholar]
- Zabadal, T.J.; Dami, I.E.; Goffinet, M.C.; Martinson, T.E.; Chien, M.L. Winter İnjury to Grapevines and Methods of Protection; Michigan State University Extension: East Lansing, MI, USA, 2007. [Google Scholar]
- Dami, I.E.; Ennahli, S.; Zhang, Y. Assessment of winter injury in grape cultivars and pruning strategies following a freezing stress event. Am. J. Enol. Vitic. 2012, 63, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, W. Vine and vineyard management following low temperature injury. In Proceedings of the ASEV Annual Meeting, Seattle, WAs, USA, 19–23 June 2000; pp. 101–110. [Google Scholar]
- Fennell, A. Freezing tolerance and injury in grapevines. J. Crop Improv. 2004, 10, 201–235. [Google Scholar] [CrossRef]
- Hatterman-Valenti, H.M.; Auwarter, C.P.; Stenger, J.E. Evaluation of cold-hardy grape cultivars for North Dakota and the North Dakota State University germplasm enhancement project. Acta Hort. 2016, 1115, 13–22. [Google Scholar] [CrossRef]
- Willwerth, J.; Ker, K.; Inglis, D. Best Management Practices for Reducing Winter İnjury in Grapevines; Technical Report; Cool Climate Oenology & Viticulture Institute, Brock University: Saint Catharines, ON, Canada, 2014; Available online: https://brocku.ca/ccovi/wp-content/uploads/sites/125/Research-Best-Practices-Manual-Winter-Injury.pdf (accessed on 26 December 2019).
- Todaro, T.M.; Dami, I.E. Trunk renewal methods for recovery of ‘Cabernet franc’ grapevines after winter injury in Ohio. HortTechnology 2018, 28, 382–390. [Google Scholar] [CrossRef]
- Goffinet, M.C. Grapevine cold injury and recovery after tissue damage and using cane burial to avoid winter injury. In Proceedings of the Understanding and Preventing Freeze Damage in Vineyards, Workshop Proceedings, University of Missouri-Columbia, Columbia, MO, USA, 5–6 December 2007. [Google Scholar]
- Wang, S.; Li, H.; Ye, Q.; Wang, H. Winter chill protection of grapevines by burial: Evaluation of the crawled cordon training system. VITIS-J. Grapevine Res. 2016, 55, 45–51. [Google Scholar] [CrossRef]
- Howell, G.S. Cultural manipulation of vine cold hardiness. In Proceedings of the Second International Cool Climate Viticulture and Oenology Symposium, Auckland, New Zealand, 11–15 January 1988; pp. 98–102. [Google Scholar]
- NOAA. National Oceanic and Atmospheric Administration. Available online: http://www.legislative.noaa.gov/NIYS/ (accessed on 11 November 2019).
- NDAWN. North Dakota Agriculture Weather Network. Available online: https://ndawn.ndsu.nodak.edu/ (accessed on 30 November 2019).
- Mills, L.J.; Ferguson, J.C.; Keller, M. Cold-hardiness evaluation of grapevine buds and cane tissues. Am. J. Enol. Vitic. 2006, 57, 194–200. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-0-387-98141-3. [Google Scholar]
- Meiering, A.G.; Paroschy, J.H.; Peterson, R.L.; Hostetter, G.; Neff, A. Mechanical freezing injury in grapevine trunks. Am. J. Enol. Vitic. 1980, 31, 81–89. [Google Scholar]
- Pratt, C.; Pool, R.M. Anatomy of recovery of canes of Vitis vinifera L. from simulated freezing injury. Am. J. Enol. Vitic. 1981, 32, 223–227. [Google Scholar]
- Keller, M.; Mills, L.J. Effect of pruning on recovery and productivity of cold-injured Merlot grapevines. Am. J. Enol. Vitic. 2007, 58, 351–357. [Google Scholar]
- Antivilo, F.G.; Paz, R.C.; Tognetti, J.; Keller, M.; Cavagnaro, M.; Barrio, E.E.; Juñent, F.R. Winter injury to grapevine secondary phloem and cambium impairs budbreak, cambium activity, and yield formation. J. Plant Growth Regul. 2019, 1, 1–12. [Google Scholar] [CrossRef]
- Antivilo, F.G.; Paz, R.C.; Echeverria, M.; Keller, M.; Tognetti, J.; Borgo, R.; Juñent, F.R. Thermal history parameters drive changes in physiology and cold hardiness of young grapevine plants during winter. Agric. For. Met. 2019, 262, 227–236. [Google Scholar] [CrossRef]
- USDA Plant Hardiness Zone Map. Agricultural Research Service. U.S. Department of Agriculture. 2012. Available online: https://planthardiness.ars.usda.gov/PHZMWeb/ (accessed on 20 May 2020).
- Clark, M.D. Development of Cold Climate Grapes in the Upper Midwestern US: The Pioneering Work of Elmer Swenson. In Plant Breeding Reviews; Goldman, I., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; Volume 43, pp. 31–60. [Google Scholar] [CrossRef]
- Todaro, T.M.; Dami, I.E. Cane morphology and anatomy influence freezing tolerance in Vitis vinifera Cabernet franc. Intl. J. Fruit Sci. 2017, 17, 391–406. [Google Scholar] [CrossRef]
- Lavee, S. Usefullness of growth regulators for controlling vine growth and improving grape quality in intensive vineyards. Acta Hort. 1986, 206, 89–108. [Google Scholar] [CrossRef]
- Wample, R.L.; Schnabel, B.; Ahmedullah, M. Leaf area and conductance, internode length, and root structure of five cultivars of Vitis vinifera treated with paclobutrazol. Am. J. Enol. Vitic. 1987, 38, 255–259. [Google Scholar]
- Chou, M.Y.; Heuvel, J.E. Annual under-vine cover crops mitigate vine vigor in a mature and vigorous Cabernet franc vineyard. Am. J. Enol. Vitic. 2019, 70, 98–108. [Google Scholar] [CrossRef]
- Gohil, H.; Mills, L.J.; Keller, M. Cordon Renewal Strategies for Declining Merlot Grapevines. Catal. Discov. Pract. 2019, 3, 1–7. [Google Scholar] [CrossRef]
- Howell, G.S.; Shaulis, N. Factors influencing within-vine variation in the cold resistance of cane and primary bud tissues. Am. J. Enol. Vitic. 1980, 31, 158–161. [Google Scholar]
- Khanizadeh, S.; Rekika, D.; Levasseur, A.; Groleau, Y.; Richer, C.; Fisher, H. The effects of different cultural and environmental factors on grapevine growth, winter hardiness and performance, in three locations, in Canada. Small Fruits Rev. 2008, 4, 3–28. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Rekika, D.; Levasseur, A.; Groleau, Y.; Richer, C.; Fisher, H. Growing grapes in a cold climate with winter temperature below −25 °C. Acta Hort. 2004, 663, 931–936. [Google Scholar] [CrossRef]
- Luo, G. Dragon system of training and pruning in China’s Viticulture. Am. J. Enol. Vitic. 1986, 37, 152–157. [Google Scholar]
- Haggerty, L.L. Ripening Profile of Grape Berry Acids and Sugars in University of Minnesota Wine Grape Cultivars, Select Vitis vinifera and Other Hybrid Cultivars (Order No. 1547856). Master’s Thesis, University of Minnesota, Minneapolis, MN, USA, 2013; p. 91. [Google Scholar]
- Shaulis, N.; Einset, J.; Pack, A.B. Growing Cold-Tender Grape Varieties in New York; Bulletin Number 821; New York State Agricultural Experiment Station: Geneva, NY, USA, 1968; pp. 1–16. [Google Scholar]





| Genotype | Establishment Year | Percent Trunks Failing to Break Bud | Percent Trunks Collapsed Midseason | Percent Surviving Trunks |
|---|---|---|---|---|
| ---Established Genotypes--- | ||||
| Alpenglow | 2004 | 100 | 0 | 0 |
| Baltica | 2004 | 14 | 18 | 68 |
| Bluebell | 2004 | 27 | 19 | 54 |
| Brianna | 2008 | 87 | 13 | 0 |
| Edelweiss | 2009 | 87 | 13 | 0 |
| E.S. 5-4-71 | 2004 | 100 | 0 | 0 |
| Frontenac | 2004 | 17 | 76 | 7 |
| Frontenac gris | 2004 | 29 | 67 | 4 |
| GR 7 | 2009 | 100 | 0 | 0 |
| John Viola | 2009 | 7 | 4 | 89 |
| Kay Gray | 2009 | 20 | 72 | 8 |
| King of the North | 2004 | 0 | 7 | 93 |
| La Crescent | 2004 | 94 | 6 | 0 |
| Laura’s Laughter | 2009 | 92 | 8 | 0 |
| Léon Millot | 2009 | 100 | 0 | 0 |
| Louise Swenson | 2009 | 90 | 10 | 0 |
| Maréchal Foch | 2009 | 100 | 0 | 0 |
| Marquette | 2005 | 100 | 0 | 0 |
| MN1131 | 2004 | 42 | 29 | 29 |
| MN1200 | 2004 | 90 | 3 | 7 |
| MN1220 | 2008 | 100 | 0 | 0 |
| MN1235 | 2008 | 81 | 19 | 0 |
| MN1258 | 2008 | 100 | 0 | 0 |
| Petite Amie | 2008 | 100 | 0 | 0 |
| Prairie Star | 2008 | 96 | 4 | 0 |
| Sabrevois | 2004 | 78 | 18 | 4 |
| Somerset Seedless | 2004 | 100 | 0 | 0 |
| St. Croix | 2004 | 89 | 11 | 0 |
| Valiant | 2004 | 0 | 0 | 100 |
| Verona | 2007 | 100 | 0 | 0 |
| ---Establishing Genotypes--- | ||||
| Adalmiina | 2017 | 88 | 12 | 0 |
| Crimson Pearl | 2014 | 92 | 0 | 8 |
| Frontenac blanc | 2012 | 89 | 11 | 0 |
| Itasca | 2017 | 100 | 0 | 0 |
| Petite Pearl | 2012 | 100 | 0 | 0 |
| Genotype | Retained Suckers (No.) | Live Nodes (No./Sucker) | Live Sucker Length (cm/Sucker) | Internode Length (cm) | Sucker Circumference (mm) | Sucker Caliper Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alpenglow | 3.7 | a 1 | 20.8 | bcde | 246.8 | bc | 11.6 | bcde | 22.3 | efg | 1.12 | abc |
| Brianna | 2.4 | ab | 21.0 | abcde | 263.4 | bc | 12.5 | abc | 25.6 | cdef | 1.09 | bc |
| Edelweiss | 2.1 | b | 19.6 | bcdef | 225.2 | bcd | 11.3 | bcdef | 23.2 | defg | 1.12 | abc |
| E.S. 5-4-71 | 2.7 | ab | 18.6 | cdef | 232.4 | bcd | 12.5 | abcd | 24.6 | cdefg | 1.10 | abc |
| Frontenac | 3.0 | ab | 18.7 | def | 230.0 | bcd | 11.7 | bcde | 29.5 | bc | 1.11 | abc |
| Frontenac gris | 3.1 | ab | 23.1 | abcd | 273.6 | b | 11.7 | bcde | 30.9 | ab | 1.11 | abc |
| GR 7 | 2.9 | ab | 24.5 | abc | 258.2 | bc | 10.2 | efg | 22.3 | efg | 1.12 | abc |
| Kay Gray | 2.7 | ab | 21.1 | abcde | 228.1 | bcd | 10.7 | cdefg | 22.3 | efg | 1.12 | abc |
| La Crescent | 3.8 | a | 26.4 | a | 360.8 | a | 13.9 | a | 34.4 | a | 1.13 | abc |
| Laura’s Laughter | 2.5 | ab | 22.2 | abcd | 252.6 | bc | 11.3 | bcdef | 26.6 | bcde | 1.11 | abc |
| Léon Millot | 2.1 | b | 26.2 | ab | 255.1 | bc | 9.7 | fgh | 23.8 | defg | 1.12 | abc |
| Louise Swenson | 2.3 | ab | 14.5 | f | 148.4 | d | 10.1 | efg | 21.1 | efg | 1.07 | c |
| Maréchal Foch | 2.6 | ab | 24.4 | abcd | 196.1 | cd | 8.0 | h | 21.2 | fg | 1.15 | ab |
| Marquette | 3.1 | ab | 21.8 | cd | 204.6 | bcd | 9.2 | gh | 23.4 | efg | 1.10 | abc |
| Prairie Star | 3.2 | ab | 19.1 | cdef | 196.4 | bcd | 10.2 | efg | 19.6 | g | 1.11 | abc |
| Sabrevois | 3.7 | a | 20.3 | bcdef | 246.5 | bc | 11.9 | bcde | 28.6 | bcd | 1.11 | abc |
| Somerset Seedless | 2.9 | ab | 20.6 | bcdef | 225.0 | bcd | 10.6 | cdefg | 23.0 | efg | 1.16 | a |
| St. Croix | 3.2 | ab | 15.8 | ef | 205.8 | bcd | 12.7 | ab | 24.6 | def | 1.08 | c |
| Verona | 2.6 | ab | 23.9 | abcd | 251.7 | bc | 10.5 | defg | 25.8 | cdef | 1.13 | abc |
| F Ratio | 2.58 | 7.47 | 8.02 | 14.72 | 15.38 | 2.40 | ||||||
| p | 0.0006 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0009 | ||||||
| Genotype | Retained Suckers (No.) | Live Nodes (No./Sucker) | Live Sucker Length (cm/Sucker) | Internode Length (cm) | Sucker Circumference (mm) | Sucker Caliper Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adalmiina | 2.5 | ns 1 | 17.3 | b | 124.7 | b | 7.3 | b | 19.7 | ns | 1.06 | ns |
| Crimson Pearl | 3.0 | 24.1 | a | 198.5 | ab | 7.7 | b | 23.4 | 1.13 | |||
| Frontenac blanc | 2.4 | 24.5 | a | 178.7 | ab | 7.0 | b | 22.4 | 1.15 | |||
| Itasca | 2.9 | 22.5 | a | 177.3 | ab | 7.8 | b | 28.0 | 1.13 | |||
| Petite Pearl | 2.2 | 23.5 | a | 226.5 | a | 9.5 | a | 23.0 | 1.10 | |||
| F Ratio | 1.14 | 5.17 | 5.94 | 7.45 | 1.64 | 1.50 | ||||||
| p | 0.346 | 0.0006 | 0.0002 | <0.0001 | 0.1679 | 0.2055 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svyantek, A.; Köse, B.; Stenger, J.; Auwarter, C.; Hatterman-Valenti, H. Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota. Horticulturae 2020, 6, 75. https://doi.org/10.3390/horticulturae6040075
Svyantek A, Köse B, Stenger J, Auwarter C, Hatterman-Valenti H. Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota. Horticulturae. 2020; 6(4):75. https://doi.org/10.3390/horticulturae6040075
Chicago/Turabian StyleSvyantek, Andrej, Bülent Köse, John Stenger, Collin Auwarter, and Harlene Hatterman-Valenti. 2020. "Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota" Horticulturae 6, no. 4: 75. https://doi.org/10.3390/horticulturae6040075
APA StyleSvyantek, A., Köse, B., Stenger, J., Auwarter, C., & Hatterman-Valenti, H. (2020). Cold-Hardy Grape Cultivar Winter Injury and Trunk Re-Establishment Following Severe Weather Events in North Dakota. Horticulturae, 6(4), 75. https://doi.org/10.3390/horticulturae6040075