
Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis



Abstract
:
1. Introduction
2. Materials and Methods
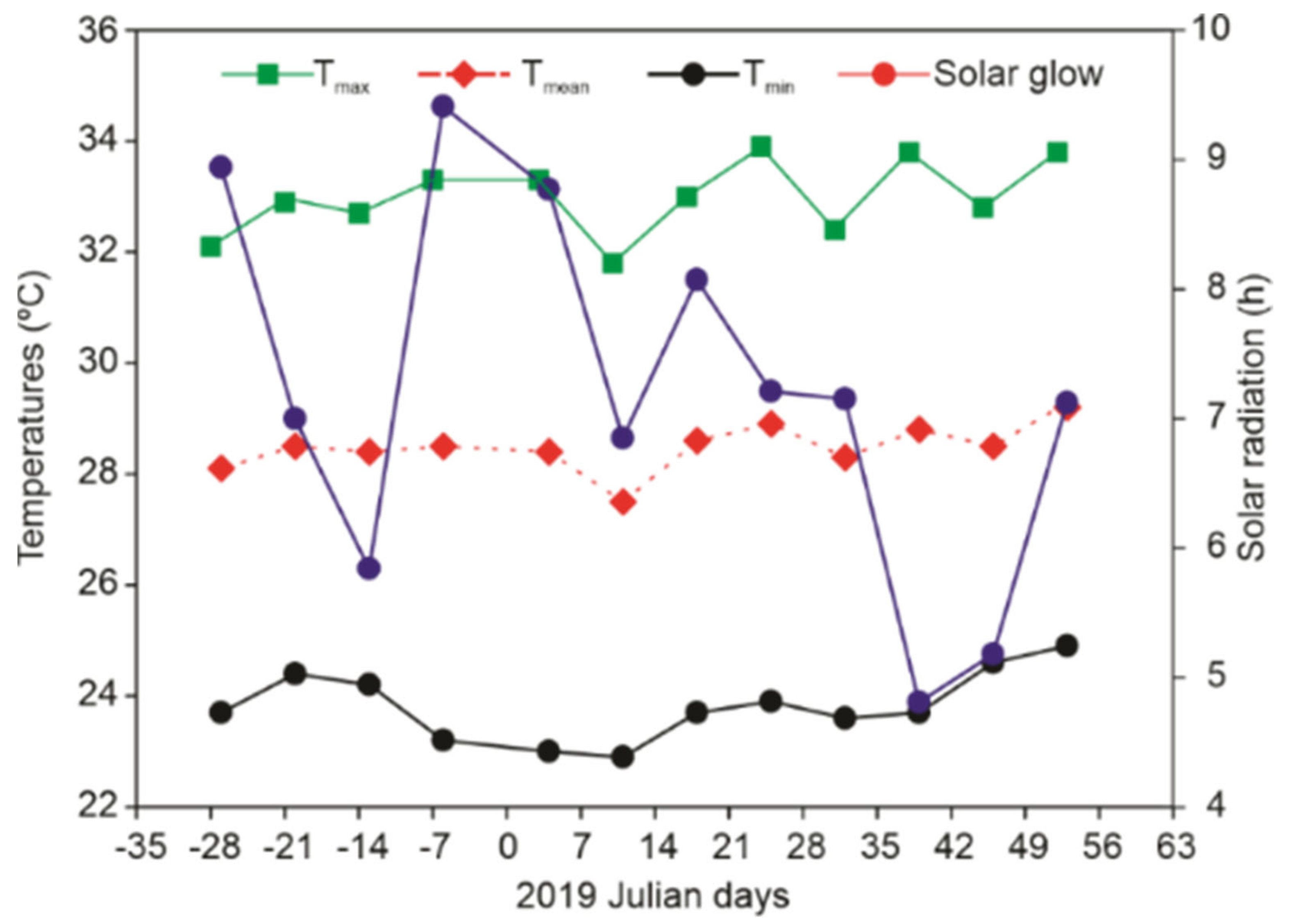
2.1. Plant Material and Environmental Conditions
2.2. Timeline of the Experiments, Leaf Harvesting, and Flower Induction
2.3. Morphological and Physiological Parameters
2.4. Analysis of Steviol Glycosides (SvGly)
2.5. Statistical Data Analysis
3. Results
3.1. Net Photosynthesis and Relative Growth
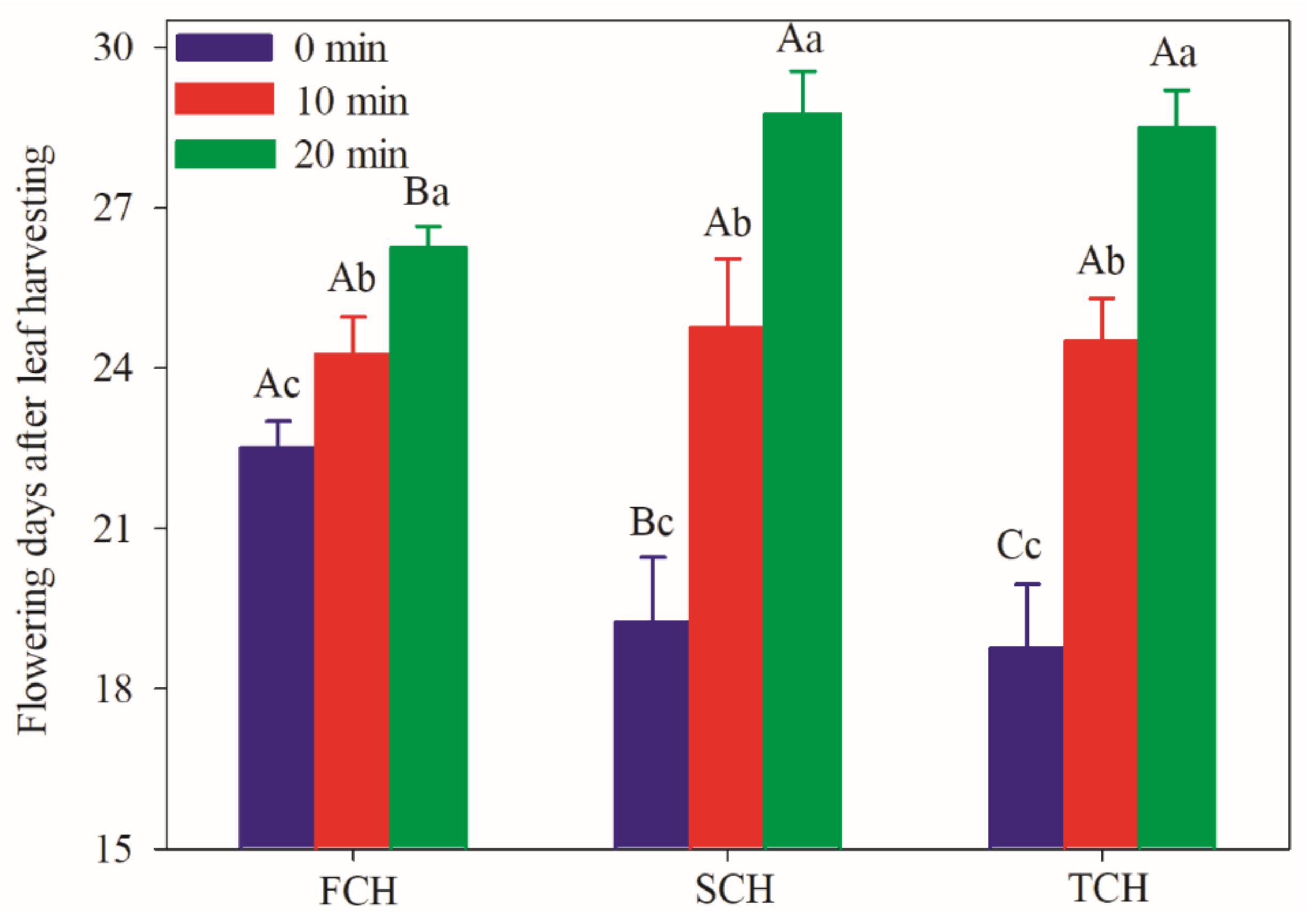
3.2. Flowering Delay
3.3. Plant Development
3.4. Steviol Glycosides
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Shaafi, B.; Mosavi, S.S.; Abdollahi, M.R.; Sarikhani, H. The optimized protocols for production, adaptation and keeping of the produced artificial seeds from encapsulated lateral buds in Stevia rebaudiana (Bertoni). Agrotech. Ind. Crop. 2021, 1, 24–35. [Google Scholar] [CrossRef]
- Ghaheri, M.; Kahrizi, D.; Bahrami, G.; Mohammadi-Motlagh, H.-R. Study of gene expression and steviol glycosides accumulation in Stevia rebaudiana Bertoni under various mannitol concentrations. Mol. Biol. Rep. 2019, 46, 7–16. [Google Scholar] [CrossRef]
- Akbari, F.; Arminian, A.; Kahrizi, D.; Fazeli, A.; Ghaheri, M. Effect of nitrogen sources on gene expression of Stevia rebaudiana (Bertoni) under in vitro conditions. Cell. Mol. Biol. 2018, 64, 11–16. [Google Scholar] [CrossRef]
- Yoneda, Y.; Shimizu, H.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Effects of light intensity and photoperiod on improving steviol glycosides content in Stevia rebaudiana (Bertoni) Bertoni while conserving light energy consumption. J. Appl. Res. Med. Aromat. Plants 2017, 7, 64–73. [Google Scholar] [CrossRef]
- Mahajan, M.; Anuradha; Pal, P.K. Attaining higher biomass and steviol glycosides yields of Stevia rebaudiana through adjustment of plant population and nitrogen rate. Ind. Crop. Prod. 2021, 165, 113426. [Google Scholar] [CrossRef]
- VMR. Global Rebaudioside A (Reb A) Market Market Size, Status and Forecast to 2028; Verified Market Research: New York, NY, USA, 2019; p. 94. [Google Scholar]
- Ceunen, S.; Werbrouck, S.; Geuns, J.M. Stimulation of steviol glycoside accumulation in Stevia rebaudiana by red LED light. J. Plant Physiol. 2012, 169, 749–752. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M.C. Steviol glycosides: Chemical diversity, metabolism, and function. J. Nat. Prod. 2013, 76, 1201–1228. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, G.J.M. Can Stevia reduce inflammation in COVID-19 disease? Arch. Food Sci. Nutr. Res. 2020, 1, 1001. [Google Scholar]
- Yadav, A.K.; Singh, S.; Dhyani, D.; Ahuja, P.S. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Espitia, M.C.; Montoya, R.B.; Atgencio, L.S. Rendimiento de Stevia rebaudiana Bert. bajo tres arreglos poblacionales en el sinú medio. Rev. UDCA Actual. Divulg. Cient. 2009, 12, 151–161. [Google Scholar]
- Ceunen, S.; Geuns, J.M. Influence of photoperiodism on the spatio-temporal accumulation of steviol glycosides in Stevia rebaudiana (Bertoni). Plant Sci. 2013, 198, 72–82. [Google Scholar] [CrossRef]
- Yoneda, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K.; Shimizu, H. Impact of blue, red, and far-red light treatments on gene expression and steviol glycoside accumulation in Stevia rebaudiana. Phytochemistry 2017, 137, 57–65. [Google Scholar] [CrossRef]
- Francisco, F.; Pereira, G.P.; Machado, M.P.; Kanis, L.A.; Deschamps, C. Characterization of Stevia rebaudiana Bertoni accessions cultived in Southern Brazil. J. Agric. Sci. 2018, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- RD Reporters and Data. Stevia Market by Form (Powdered, Liquid and Others), by Types (Reb A, Reb M, Red D and Others), by End Users, and by Distribution Channel (Online and Offline), and Segment Forecasts, 2020–2028; RD Reporters and Data: New York, NY, USA, 2021; Available online: https://www.reportsanddata.com/report-detail/stevia-market (accessed on 6 September 2021).
- Putnik, P.; Bezuk, I.; Barba, F.J.; Lorenzo, J.M.; Polunić, I.; Bursać, D.K. Sugar reduction: Stevia rebaudiana Bertoni as a natural sweetener. In Agri-Food Industry Strategies for Healthy Diets and Sustainability; Barba, F.J., Putnik, P., Kovačević, D.B., Eds.; Academic Press: San Diego, CA, USA, 2020; pp. 123–152. [Google Scholar]
- Truong, T.T.; Valíěek, P. Verification of growth and stevioside content of Stevia plants propagated by vegetative and generative method. Agric. Trop. Subtrop. 1999, 32, 79–84. [Google Scholar]
- Zaidan, L.B.P.; Dietrich, S.M.C.; Felippe, G.M. Effect of photoperiod on flowering and stevioside content in plants of Stevia rebaudiana bertoni. Jpn. J. Crop. Sci. 1980, 49, 569–574. [Google Scholar] [CrossRef]
- Idrees, M.; Sania, B.; Hafsa, B.; Kumari, S.; Khan, H.; Fazal, H.; Ahmad, I.; Akbar, F.; Ahmad, N.; Ali, S.; et al. Spectral lights trigger biomass accumulation and production of antioxidant secondary metabolites in adventitious root cultures of Stevia rebaudiana (Bert.). C. R. Biol. 2018, 341, 334–342. [Google Scholar] [CrossRef]
- Jarma-Orozco, A.; Combatt-Caballero, E.; Jaraba-Navas, J. Growth and development of Stevia rebaudiana Bert., in high and low levels of radiation. Curr. Plant Biol. 2020, 22, 100144. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Bautista-Aguilar, J.R. The effect of light quality on growth and development of in vitro plantlet of Stevia rebaudiana Bertoni. Sugar Tech 2016, 19, 331–336. [Google Scholar] [CrossRef]
- Pandey, H.; Pandey, P.; Pandey, S.S.; Singh, S.; Banerjee, S. Meeting the challenge of stevioside production in the hairy roots of Stevia rebaudiana by probing the underlying process. Plant Cell Tissue Organ Cult. 2016, 126, 511–521. [Google Scholar] [CrossRef]
- Pico-González, A.I.; Aramendiz-Tatis, H.; Pérez-Polo, D.J. Caracterización morfoagronómica de 25 clones de estevia (Stevia rebaudiana Bertoni.), en condiciones del valle del Sinú medio. Temas Agrários. 2020, 25, 106–116. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Mendes, K.R.; Ramos, M.V.; Santos, J.N.B.; Youssef, D.T.A.; Pereira, J.D.; Endres, L.; Jarma-Orozco, A.; Solano-Gomes, R.; Jarma-Arroyo, B.; et al. Mesophyll thickness and sclerophylly among Calotropis procera morphotypes reveal water-saved adaptation to environments. J. Arid Land 2019, 11, 795–810. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Sharma, S.; Sood, S. Yield components, light interception and marker compound accumulation of stevia (Stevia rebaudiana Bertoni) affected by planting material and plant density under western Himalayan conditions. Arch. Agron. Soil Sci. 2014, 60, 1731–1745. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Geuns, J.M.C. Steviol glycosides correlation to genes transcription revealed in gibberellin and paclobutrazol-treated Stevia rebaudiana. J. Plant Biochem. Biotechnol. 2017, 26, 387–394. [Google Scholar] [CrossRef]
- Ramesh, K.; Singh, V.; Megeji, N.W. Cultivation of Stevia [Stevia rebaudiana (Bert.) Bertoni]: A Comprehensive Review. Adv. Agron. 2006, 89, 137–177. [Google Scholar] [CrossRef]
- Hernández-Fernandéz, I.A.; Jarma-Orozco, A.; Pompelli, M.F. Allometric models for non-destructive leaf area measurement of stevia: An in depth and complete analysis. Hortic. Bras. 2021, 39, 205–215. [Google Scholar] [CrossRef]
- Vecteezy. Stevia Rebaudiana Vectors. Corel Draw 2021; Vecteezy Free License: New York, NY, USA, 2021. [Google Scholar]
- Santos, O.D.O.D.; Mendes, K.R.; Martins, S.V.C.; Batista-Silva, W.; dos Santos, M.A.; de Figueirôa, J.M.; de Souza, E.R.; Fernandes, D.; Araújo, W.L.; Pompelli, M.F. Physiological parameters and plasticity as key factors to understand pioneer and late successional species in the Atlantic Rainforest. Acta Physiol. Plant. 2019, 41, 145. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- St-Pierre, A.P.; Shikon, V.; Schneider, D.C. Count data in biology-Data transformation or model reformation? Ecol. Evol. 2018, 8, 3077–3085. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef]
- Durand, M.; Mainson, D.; Porcheron, B.; Maurousset, L.; Lemoine, R.; Pourtau, N. Carbon source–sink relationship in Arabidopsis thaliana: The role of sucrose transporters. Planta 2017, 247, 587–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Iersel, M. Carbon use efficiency depends on growth respiration, maintenance respiration, and relative growth rate. A case study with lettuce. Plant Cell Environ. 2003, 26, 1441–1449. [Google Scholar] [CrossRef]
- Nilwik, H.J.M. Growth analysis of sweet pepper (Capsicum annuum L.). Ann. Bot. 1981, 48, 137–146. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.B.; Kruger, E.L.; Reich, P.B. Relative growth rate in relation to physiological and morphological traits for northern hardwood tree seedlings: Species, light environment and ontogenetic considerations. Oecologia 1993, 96, 219–231. [Google Scholar] [CrossRef]
- Erwin, J.E.; Heins, R.D. Thermomorphogenic responses in stem and leaf development. HortScience 1995, 30, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, J.; Wulff, R.D. Light spectral quality, phytochrome and plant competition. Trends Ecol. Evol. 1993, 8, 47–51. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Ceunen, S.; Geuns, J.M.; Ende, W.V.D.; De Ley, M. UDP-dependent glycosyltransferases involved in the biosynthesis of steviol glycosides. J. Plant Physiol. 2011, 168, 1136–1141. [Google Scholar] [CrossRef]
- Zeevaart, J.; Gage, D.A. ent-kaurene biosynthesis is enhanced by long photoperiods in the long-day plants Spinacia oleracea L. and Agrostemma githago L. Plant Physiol. 1993, 101, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Hajihashemi, S.; Geuns, J.M.C.; Ehsanpour, A.A. Gene transcription of steviol glycoside biosynthesis in Stevia rebaudiana Bertoni under polyethylene glycol, paclobutrazol and gibberellic acid treatments in vitro. Acta Physiol. Plant. 2013, 35, 2009–2014. [Google Scholar] [CrossRef]
- Yoneda, Y.; Shimizu, H.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Effect of treatment with gibberellin, gibberellin biosynthesis inhibitor, and auxin on steviol glycoside content in Stevia rebaudiana Bertoni. Sugar Tech 2017, 20, 482–491. [Google Scholar] [CrossRef]
- Bayraktar, M.; Naziri, E.; Akgun, I.H.; Karabey, F.; Ilhan, E.; Akyol, B.; Bedir, E.; Gurel, A. Elicitor induced stevioside production, in vitro shoot growth, and biomass accumulation in micropropagated Stevia rebaudiana. Plant Cell Tissue Organ Cult. 2016, 127, 289–300. [Google Scholar] [CrossRef]
- Park, Y.G.; Muneer, S.; Soundararajan, P.; Manivnnan, A.; Jeong, B.R. Light quality during night interruption affects morphogenesis and flowering in Petunia hybrida, a qualitative long-day plant. Hortic. Environ. Biotechnol. 2016, 57, 371–377. [Google Scholar] [CrossRef]
- Somers, D.E.; Kim, W.-Y.; Geng, R. The F-Box protein ZEITLUPE confers dosage-dependent control on the circadian clock, photomorphogenesis, and flowering time [W]. Plant Cell 2004, 16, 769–782. [Google Scholar] [CrossRef] [Green Version]
- Serfaty, M.; Ibdah, M.; Fischer, R.; Chaimovitsh, D.; Saranga, Y.; Dudai, N. Dynamics of yield components and stevioside production in Stevia rebaudiana grown under different planting times, plant stands and harvest regime. Ind. Crop. Prod. 2013, 50, 731–736. [Google Scholar] [CrossRef]
- Kumar, R.; Sood, S.; Sharma, S.; Kasana, R.; Pathania, V.; Singh, B.; Singh, R. Effect of plant spacing and organic mulch on growth, yield and quality of natural sweetener plant Stevia and soil fertility in western Himalayas. Int. J. Plant Prod. 2014, 8, 311–334. [Google Scholar] [CrossRef]
- Gomes, E.N.; Moterle, D.; Biasi, L.A.; Koehler, H.S.; Kanis, L.A.; Deschamps, C. Plant densities and harvesting times on productive and physiological aspects of Stevia rebaudiana Bertoni grown in southern Brazil. An. Acad. Bras. Cienc. 2018, 90, 3249–3264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, P.K.; Prasad, R.; Pathania, V. Effect of decapitation and nutrient applications on shoot branching, yield, and accumulation of secondary metabolites in leaves of Stevia rebaudiana Bertoni. J. Plant Physiol. 2013, 170, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Mejeji, N.W.; Kumar, J.K.; Singh, V.; Kaul, V.K.; Ahuka, P.S. Introducing Stevia rebaudiana, a natural zerocalorie sweetener. Curr. Sci. 2005, 88, 801–804. [Google Scholar]
- González, H.R.; Luz, L.L.A.; Sosa, I.H.; Amita, M.M.R.; Ferradá, C.A.R.; Govín, E.S.; Figueredo, M.M. Comportamiento del cultivo de Stevia rebaudiana (Bertoni) Bertoni en Cuba. Rev. Cubana Plant. Med. 2007, 12, 1–5. [Google Scholar]
- Higuchi, Y.; Narumi, T.; Oda, A.; Nakano, Y.; Sumitomo, K.; Fukai, S.; Hisamatsu, T. The gated induction system of a systemic floral inhibitor, antiflorigen, determines obligate short-day flowering in chrysanthemums. Proc. Natl. Acad. Sci. USA 2013, 110, 17137–17142. [Google Scholar] [CrossRef] [Green Version]
- Sumitomo, K.; Higuchi, Y.; Aoki, K.; Miyamae, H.; Oda, A.; Ishiwata, M.; Yamada, M.; Nakayama, M.; Hisamatsu, T. Spectral sensitivity of flowering and FT- like gene expression in response to night-break light treatments in the chrysanthemum cultivar, ‘Reagan’. J. Hortic. Sci. Biotechnol. 2012, 87, 461–469. [Google Scholar] [CrossRef]
- Bernier, G.E. The control of floral evocation and morphogenesis. Annu. Rev. Plant Biol. 1988, 39, 175–219. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M. Spatio-temporal variation of the diterpene steviol in Stevia rebaudiana grown under different photoperiods. Phytochemistry 2013, 89, 32–38. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, S.; Ramesh, K.; Singh, B. Effects of shade regimes and planting geometry on growth, yield and quality of the natural sweetener plant stevia (Stevia rebaudiana Bertoni) in north-western Himalaya. Arch. Agron. Soil Sci. 2013, 59, 963–979. [Google Scholar] [CrossRef]
- Hernández, J.L.; Combatt, E.M.; Jarma-Orozco, A.; Polo, J.S.; Rodrígues, L.P. Rendimiento y calidad de hojas de Stevia rebaudiana bert. bajo la oferta edafológica y dos niveles de radiación en cinco regiones de Colombia. Rev. UDCA Actual. Divulg. Cient. 2016, 19, 77–85. [Google Scholar]
- Bae, G.; Choi, G. Decoding of light signals by plant phytochromes and their interacting proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef] [Green Version]
- Jarma-Orozco, A.; Ayala, C.C.; Herrera, C.F. Temperature and radiation effect on steviol glycosides production in Stevia rebaudiana in the Colombian humid Caribbean region. Rev. UDCA Actual. Divulg. Cient. 2012, 15, 339–347. [Google Scholar]
- Pompelli, M.F.; Martins, S.C.; Antunes, W.C.; Chaves, A.R.; DaMatta, F.M. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, C.; Marín, L.; Habeych, D. Obtención de edulcorantes de Stevia rebaudiana Bertoni. Rev. CENIC Cienc. Biol. 2005, 36, 3–10. [Google Scholar]
- Parris, C.A.; Shock, C.C.; Qian, M. Dry Leaf and Steviol Glycoside Productivity of Stevia rebaudiana in the Western United States. HortScience 2016, 51, 1220–1227. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Storck, L.; Lopes, S.J.; Martin, T.N.; Bisognin, D.A. Dry biomass and glycosides yield from Stevia rebaudiana leaves under different harvesting times. Biosci. J. 2016, 32, 1462–1471. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, Y.; Hou, M.; Huang, X.; Zhang, T.; Huang, S.; Xu, X.; Yuan, H. Optimized nitrogen topdressing strategies enhance steviol glycoside productivity in stevia (Stevia rebaudiana Bertoni) Plants. J. Soil Sci. Plant Nutr. 2020, 20, 1133–1143. [Google Scholar] [CrossRef]
- Benhmimou, A.; Ibriz, M.; Al Faïz, C.; Gaboun, F.; Douaik, A.; Amchra, F.Z.; Khiraoui, A.; Lage, M. Effects of planting density and harvesting time on productivity of natural sweetener plant (Stevia rebaudiana Bertoni.) in Larache Region, Morocco. Int. J. Plant Res. 2017, 7, 83–89. [Google Scholar] [CrossRef]
- Shulgina, A.; Kalashnikova, E.; Tarakanov, I.; Kirakosyan, R.; Cherednichenko, M.; Polivanova, O.; Baranova, E.; Khaliluev, M. Influence of light conditions and medium composition on morphophysiological characteristics of Stevia rebaudiana Bertoni in vitro and in vivo. Horticulturae 2021, 7, 195. [Google Scholar] [CrossRef]
- Nissim-Levi, A.; Kitron, M.; Nishri, Y.; Ovadia, R.; Forer, I.; Oren-Shamir, M. Effects of blue and red LED lights on growth and flowering of Chrysanthemum morifolium. Sci. Hortic. 2019, 254, 77–83. [Google Scholar] [CrossRef]
- Hasanaklou, H.T.; Ghadim, A.E.K.; Moradi, F.; Ghodehkahriz, S.J.; Gholipouri, A. The effects of NH4+ and NO3− and plant growth regulators on the accumulation of nutrients, carbohydrates and secondary metabolites of Stevia rebaudiana bertoni. Sugar Tech 2020, 23, 65–76. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Area (cm2) | ||||||
|---|---|---|---|---|---|---|
| PA | Genotype 2 | Genotype 4 | ||||
| 0 min | 10 min | 20 min | 0 min | 10 min | 20 min | |
| 29 | 140.2 ± 29.3 Af | 112.9 ± 41.3 Af | 109.4 ± 33.8 Ag | 99.8 ± 28.1 Ah | 55.7 ± 31.8 Af | 58.9 ± 14.8 Ag |
| 36 | 321.0 ± 24.1 Ad | 374.5 ± 60.5 Ad | 330.6 ± 56.2 Ae | 265.4 ± 12.8 Ag | 236.9 ± 20.2 Ae | 175.2 ± 30.4 Bf |
| 43 | 273.2 ± 44.5 Ae * | 261.3 ± 39.0 Ae * | 237.7 ± 10.9 Af * | 496.3 ± 74.3 Af | 405.6 ± 52.9 Ad | 490.0 ± 33.3 Ae |
| 50 | 466.6 ± 45.2 Ad * | 371.2 ± 54.1 Ad * | 381.9 ± 38.5 Ad * | 856.0 ± 22.3 Be | 930.8 ± 32.5 Ac | 897.8 ± 75.4 Ad |
| 57 | 498.1 ± 127.2 Bc * | 704.7 ± 126.7 Ac * | 622.1 ± 108.0 Ac * | 1144.0 ± 209.1 Ad | 1222.5 ± 172.9 Ab | 923.3 ± 143.3 Ac |
| 64 | 946.1 ± 169.2 Bb * | 1305.0 ± 202.4 Aab | 1363.3 ± 175.2 Ab* | 1953.9 ± 111.5 Ac | 2151.8 ± 551.2 Aab | 2131.5 ± 165.5 Ab |
| 71 | 1152.2 ± 327.0 Ba * | 1146.2 ± 288.4 Bb * | 1558.0 ± 367.6 Aa * | 2800.9 ± 454.1 Aa | 2554.8 ± 389.6 Aa | 2679.9 ± 344.9 Aa |
| 78 | 1676.8 ± 299.8 Aa * | 1546.8 ± 452.5 Aa * | 1363.4 ± 214.4 Bb * | 2484.0 ± 184.4 Bb | 2999.8 ± 709.2 Aa | 2982.5 ± 470.5 Aa |
| 85 | 1906.0 ± 148.5 Bb * | 2993.4 ± 558.2 Aa | 2375.2 ± 552.3 Aa | 2843.3 ± 226.7 Bb | 3904.4 ± 609.5 Aa | 2488.2 ± 357.7 Bb |
| PA | Specific leaf weight (g m−2) | |||||
| 0 min | 10 min | 20 min | 0 min | 10 min | 20 min | |
| 29 | 36.4 ± 2.6 Ac | 38.5 ± 4.1 Ab * | 41.4 ± 10.2 Ac | 44.5 ± 3.5 Ad | 19.0 ± 2.4 Cd | 30.2 ± 4.6 Bc |
| 36 | 49.7 ± 7.7 Ab | 43.6 ± 7.2 Ab | 52.9 ± 15.8 Aa | 60.6 ± 11.2 Ac | 51.3 ± 7.7 Ab | 53.1 ± 8.6 Ab |
| 43 | 40.6 ± 1.1 Bb | 47.0 ± 14.3 Aa | 39.7 ± 0.4 Bc * | 44.4 ± 2.9 Ad | 40.5 ± 2.0 Cc | 46.4 ± 2.9 Bb |
| 50 | 44.0 ± 2.8 Ab | 47.5 ± 4.9 Aa | 44.4 ± 1.7 Ab | 50.6 ± 1.2 Ac | 45.3 ± 2.0 Bc | 45.3 ± 0.6 Bb |
| 57 | 62.1 ± 3.6 Aa | 39.4 ± 2.1 Bb | 58.7 ± 14.1 Aa | 50.3 ± 7.0 Ac | 61.9 ± 11.8 Ab | 52.4 ± 20.4 Ab |
| 64 | 46.3 ± 8.9 Bb * | 50.6 ± 5.1 Aa | 38.9 ± 5.1 Bc | 61.7 ± 4.2 Ac | 54.3 ± 7.4 ABb | 48.5 ± 13.2 Bbc |
| 71 | 43.7 ± 7.0 Ab | 39.3 ± 2.6 Ab | 45.1 ± 5.7 Aa | 50.6 ± 2.7 Ac | 51.2 ± 6.3 Ab | 77.1 ± 16.0 Aa |
| 78 | 52.7 ± 6.0 Aa * | 49.6 ± 6.5 Ba * | 52.4 ± 7.7 Aa | 94.8 ± 5.1 Aa | 80.7 ± 4.2 Ba | 72.0 ± 10.9 Ba |
| 85 | 46.8 ± 3.7 Ab * | 39.4 ± 4.17 Bb * | 38.6 ± 7.3 Bc * | 75.2 ± 5.8 Ab | 53.8 ± 2.1 Bb | 70.0 ± 5.9 Aa |
| PA | Leaf area ratio (cm2 g−1) | |||||
| 0 min | 10 min | 20 min | 0 min | 10 min | 20 min | |
| 29 | 4.0 ± 1.0 Ae | 3.2 ± 1.2 Ae | 3.5 ± 1.5 Ae | 2.4 ± 0.7 Ag | 2.5 ± 1.1 Af | 1.9 ± 0.3 Af |
| 36 | 6.7 ± 0.6 Bd | 8.6 ± 0.5 Ad * | 7.5 ± 1.8 ABcd | 4.7 ± 0.7 Af | 5.0 ± 1.0 Ae | 3.4 ± 0.7 Ae |
| 43 | 6.8 ± 1.3 Ad | 12.3 ± 8.3 Ac | 6.0 ± 0.3 Ad * | 11.4 ± 1.9 Ae | 10.3 ± 1.8 Ad | 10.6 ± 0.8 Ad |
| 50 | 10.9 ± 1.6 Ac * | 8.0 ± 1.4 Ad * | 8.7 ± 1.0 Ac * | 17.0 ± 0.8 Bd | 20.7 ± 1.1 Acd | 19.8 ± 1.8 Ac |
| 57 | 7.8 ± 2.6 Bd | 18.3 ± 3.9 Ac | 14.5 ± 5.6 ABc | 27.7 ± 7.0 Abc | 21.4 ± 5.8 ABc | 14.0 ± 5.4 Bcd |
| 64 | 24.5 ± 2.6 Bb | 27.6 ± 6.5 ABb | 37.7 ± 8.1 Aab | 31.9 ± 1.8 Ab | 27.4 ± 2.5 Abc | 36.5 ± 8.7 Ab |
| 71 | 31.0 ± 11.8 Ab | 30.7 ± 9.1 Ab | 37.3 ± 11.8 Aab | 56.1 ± 9.6 Aa | 58.6 ± 7.0 Aa | 49.7 ± 4.2 Aa |
| 78 | 34.9 ± 9.2 Aab | 38.4 ± 18.5 Ab | 29.3 ± 8.3 Ab | 26.6 ± 2.9 Bc | 27.7 ± 3.2 Bb | 47.4 ± 13.8 Aab |
| 85 | 42.3 ± 6.9 Ba | 117.1 ± 5.0 Aa * | 38.7 ± 2.7 Ba | 39.3 ± 6.5 Bb | 64.2 ± 11.1 Aa * | 37.0 ± 8.0 Bb |
| Genotype 2 | Genotype 4 | ||||
|---|---|---|---|---|---|
| Stevioside (g kg−1 DW) | |||||
| 0 min | 10 min | 20 min | 0 min | 10 min | 20 min |
| 109.63 ± 1.80 Bb | 112.83 ± 3.05 Bb | 128.89 ± 1.28 Ba | 124.81 ± 1.94 Ac | 135.32 ± 0.51 Ab | 155.55 ± 2.22 Aa |
| Rebaudioside A (g kg−1 DW) | |||||
| 83.67 ± 1.06 Bc | 89.75 ± 1.10 Bb | 100.73 ± 1.31 Ba | 102.85 ± 1.52 Ac | 112.86 ± 0.56 Ab | 132.07 ± 1.27 Aa |
| Rebaudioside A/Stevioside ratio | |||||
| 0.77 ± 0.02 Ba | 0.80 ± 0.02 Aa | 0.78 ± 0.01 Ba | 0.83 ± 0.02 Aa | 0.83 ± 0.01 Aa | 0.85 ± 0.02 Aa |
| Stevioside yield (kg ha−1) | |||||
| 986.68 ± 16.22 Bb | 1015.51 ± 27.42 Bb | 1160.00 ± 17.46 Ba | 1123.32 ± 17.46 Ac | 1217.85 ± 4.60 Ab | 1399.93 ± 20.02 Aa |
| Rebaudioside A yield (kg ha−1) | |||||
| 752.99 ± 9.57 Bc | 807.74 ± 9.89 Bb | 906.55 ± 11.79 Ba | 925.65 ± 13.65 Ac | 1015.78 ± 5.01 Ab | 1178.64 ± 11.46 Aa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Avilez, J.A.; Jarma-Orozco, A.; Pompelli, M.F. Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis. Horticulturae 2021, 7, 543. https://doi.org/10.3390/horticulturae7120543
Rivera-Avilez JA, Jarma-Orozco A, Pompelli MF. Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis. Horticulturae. 2021; 7(12):543. https://doi.org/10.3390/horticulturae7120543
Chicago/Turabian StyleRivera-Avilez, Jesús Antonio, Alfredo Jarma-Orozco, and Marcelo F. Pompelli. 2021. "Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis" Horticulturae 7, no. 12: 543. https://doi.org/10.3390/horticulturae7120543
APA StyleRivera-Avilez, J. A., Jarma-Orozco, A., & Pompelli, M. F. (2021). Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis. Horticulturae, 7(12), 543. https://doi.org/10.3390/horticulturae7120543