
Deciphering Resistance to Root-Knot Nematodes in Prunus for Rootstock Breeding: Sources, Genetics and Characterization of the Ma Locus


Abstract
:1. Introduction
2. Root-Knot Nematodes
2.1. Biology and Life Cycle
2.2. The RKNs Affecting Prunus Species
3. Diversity of Prunus Species and Rootstocks
- The subgenus Amydalus includes diploid species (2n = 2x = 16) such as the cultivated peach (Prunus persica), wild peaches P. davidiana and P. kansuensis, the cultivated almond (P. dulcis) and the wild almond P. webbii.
- The subgenus Prunophora or Prunus senso stricto includes hexaploid (P. domestica, P. insititia; 2n = 6x = 48), tetraploid (P. spinosa; 2n = 4x = 32) and diploid (P. cerasifera, P. salicina; 2n = 2x = 16) plums as well as the apricots (P. armeniaca, P. mume, P. brigantina; 2n = 2x = 16).
4. Control of RKNs in Prunus: The Resistance Strategy
4.1. Resistance Sources
4.2. Genetics and Mapping of RKN Resistance
4.3. Histological Mechanisms of RKN Resistance
4.4. Other Features of Resistance in Prunus: Durability, Stability at High Temperature and Relationship with Other Pests and Pathogens
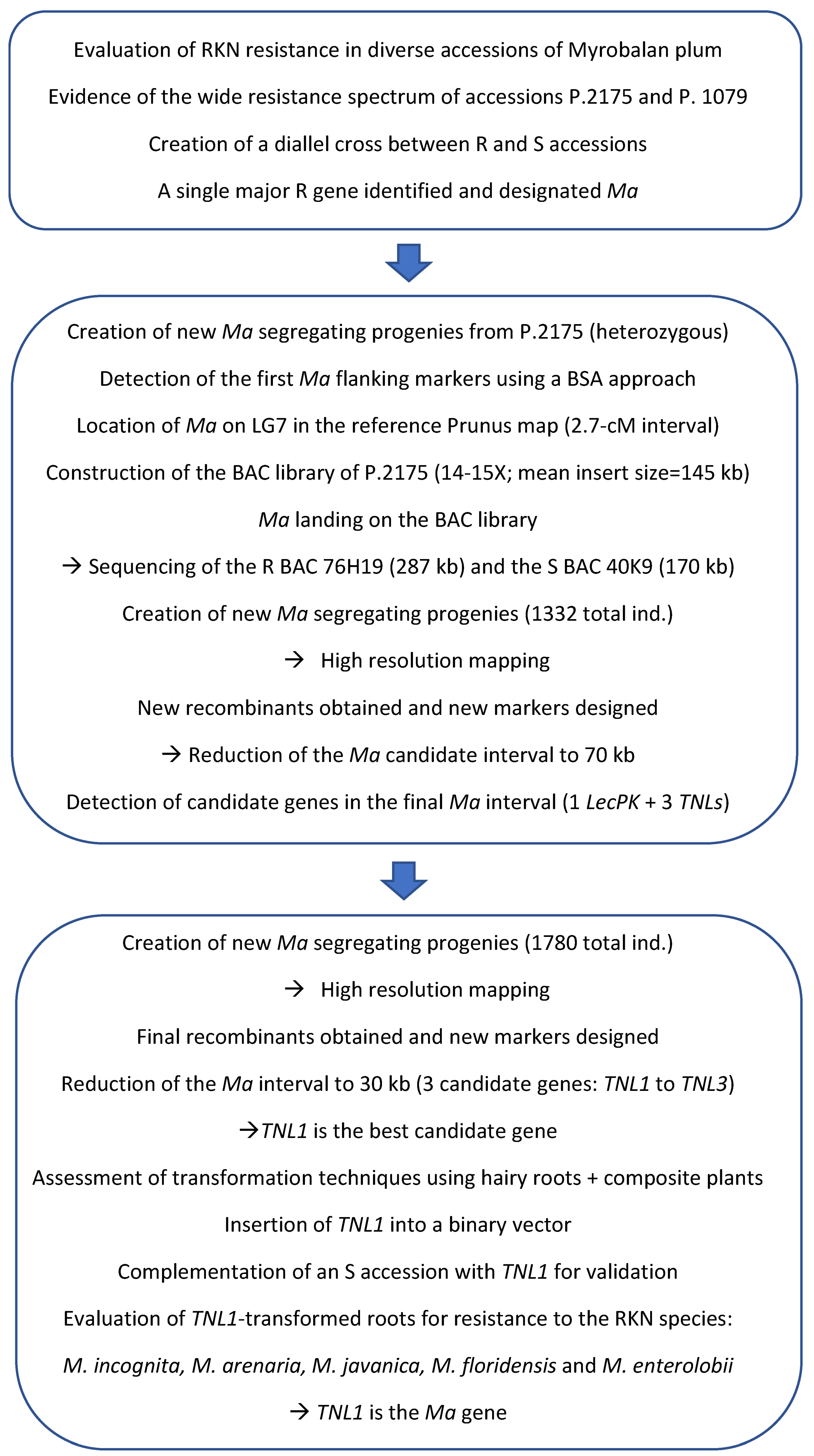
5. High-Resolution Mapping and Cloning of the Ma Locus
5.1. Positional Cloning and Functional Characterization of the Ma Gene
5.1.1. High-Resolution Mapping, BAC Library Construction and Chromosome Landing
5.1.2. Evidence of the TNL Cluster Containing Ma
5.1.3. Validation of the Final Candidate TNL1 and Evidence of the Remarkable Structure of the Ma Gene
5.2. High-Resolution Mapping of the Almond RMja Gene, a Ma Orthologue
6. Conclusions and Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nyczepir, A.P.; Halbrendt, J.M. Nematode pests of deciduous fruit and nut trees. In Plant Parasitic Nematodes in Temperate Agriculture; Evans, K., Trudgill, D.L., Webster, J.M., Eds.; CABI: Wallingford, UK, 1993; pp. 381–425. [Google Scholar]
- Reighard, G.L. Current Directions of Peach Rootstock Programs Worldwide. In Proceedings of the 5th International Peach Symposium, Davis, CA, USA, 8–11 July 2021; pp. 421–427. [Google Scholar]
- Nyczepir, A.; Esmenjaud, D. Peach nematodes. In The Peach: Botany, Production and Uses; Wayne, D.R., Bassi, D., Eds.; CAB Editions: New York, NY, USA, 2007; pp. 505–535. [Google Scholar]
- Reighard, G. Breeding Stone Fruit Rootstocks for High-efficiency Orchards. Hortscience 2016, 51, S86. [Google Scholar]
- Lamberti, F. Economic importance of Meloidogyne spp. In subtropical and mediterranean climates. In Root-Knot Nematodes (Meloidogyne Species): Systematics, Biology and Control; Lamberti, F., Taylor, C.E., Eds.; Academic Press: New York, NY, USA, 1979; pp. 341–357. [Google Scholar]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-Lopez, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Nyczepir, A.P.; Miller, R.W.; Beckman, T.G. Root-knot nematodes on peach in the southeastern United States: An update and advances. Afr. Plant Protect. 1997, 3, 115. [Google Scholar]
- De Guiran, G.; Netscher, G. Les nématodes du genre Meloidogyne parasites des cultures tropicales. Cah. ORSTOM Ser. Biol. 1970, 3, 151–185. [Google Scholar]
- Abad, P.; Williamson, V.M. Plant nematode interaction: A sophisticated dialogue. In Advances in Botanical Research; Kader, J.C., Delseny, M., Eds.; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2010; Volume 53, pp. 147–192. [Google Scholar]
- Esmenjaud, D.; Minot, J.C.; Voisin, R.; Pinochet, J.; Salesses, G. Inter- and intraspecific resistance variability in Myrobalan plum, peach and peach-almond rootstocks using 22 root-knot nematode populations. J. Am. Soc. Hortic. Sci. 1994, 119, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, R.H.; Hesse, C.O.; Lownsbery, B.A.; Perry, V.G.; Hansen, C.J. Breeding peaches for root-knot nematode resistance. J. Am. Soc. Hortic. Sci. 1969, 94, 209–212. [Google Scholar]
- Handoo, Z.A.; Nyczepir, A.P.; Esmenjaud, D.; van der Beek, J.G.; Castagnone-Sereno, P.; Carta, L.K.; Skantar, A.M.; Higgins, J.A. Morphological, molecular, and differential-host characterization of Meloidogyne floridensis n. sp (Nematoda: Meloidogynidae), a root-knot nematode parasitizing peach in Florida. J. Nematol. 2004, 36, 20–35. [Google Scholar]
- Saucet, S.B.; Van Ghelder, C.; Abad, P.; Duval, H.; Esmenjaud, D. Resistance to root-knot nematodes Meloidogyne spp. in woody plants. New Phytol. 2016, 211, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Nyczepir, A.P.; Esmenjaud, D.; Eisenback, J.D. Pathogenicity of Meloidogyne sp. (FL-isolate) on Prunus in the southeastern United States and France. J. Nematol. 1998, 30, 509. [Google Scholar]
- Stanley, J.D.; Brito, J.A.; Kokalis-Burelle, N.; Frank, J.H.; Dickson, D.W. Biological evaluation and comparison of four Florida isolates of Meloidogyne floridensis. Nematropica 2009, 39, 255–271. [Google Scholar]
- Reighard, G.L.; Henderson, W.G.; Scott, S.O.; Subbotin, S.A. First Report of the Root-Knot Nematode, Meloidogyne floridensis Infecting Guardian (R) Peach Rootstock in South Carolina, USA. J. Nematol. 2019, 51, e2019-61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez, J.; Forghani, F.; Hajihassani, A. The occurrence of Meloidogyne floridensis on collard greens in Georgia, USA. Phytopathology 2020, 110, 9. [Google Scholar]
- Marquez, J.; Forghani, F.; Hajihassani, A. First Report of the Root-Knot Nematode, Meloidogyne floridensis, on Tomato in Georgia, USA. Plant Dis. 2021, 105, 1228. [Google Scholar] [CrossRef]
- Westphal, A.; Maung, Z.T.Z.; Doll, D.A.; Yaghmour, M.A.; Chitambar, J.J.; Subbotin, S.A. First Report of the Peach Root-Knot Nematode, Meloidogyne floridensis Infecting Almond on Root-Knot Nematode Resistant ‘Hansen 536’ and ‘Bright’s Hybrid 5’ Rootstocks in California, USA. J. Nematol. 2019, 51, e2019-02. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnone-Sereno, P. Meloidogyne enterolobii (=M. mayaguensis): Profile of an emerging, highly pathogenic, root-knot nematode species. Nematology 2012, 14, 133–138. [Google Scholar] [CrossRef]
- Rammah, A.; Hirschmann, H. Meloidogyne mayaguensis n. sp. (Meloidogynidae), a root-knot nematode from Puerto-Rico. J. Nematol. 1988, 20, 58–69. [Google Scholar]
- Yang, B.; Eisenback, J.D. Meloidogyne enterolobii n.sp. (Meloidogynidae), a root-knot nematode parasitizing pacara earpot tree in China. J. Nematol. 1983, 15, 381–391. [Google Scholar]
- Rammah, A.; Hirschmann, H. Meloidogyne morocciensis n. sp. (Meloidogyninae), a root-knot nematode from Morocco. J. Nematol. 1990, 22, 279–291. [Google Scholar]
- Hirschmann, H. Meloidogyne hispanica nsp. (Nematoda, Meloidogynidae), the Seville root-knot nematode. J. Nematol. 1986, 18, 520–532. [Google Scholar]
- Meza, P.; Soto, B.; Rojas, L.; Esmenjaud, D. Identification of Meloidogyne Species from the Central Valley of Chile and Interaction with Stone Fruit Rootstocks. Plant Dis. 2016, 100, 1358–1363. [Google Scholar] [CrossRef] [Green Version]
- Esbenshade, P.R.; Triantaphyllou, A.C. Isozyme phenotypes for the identification of Meloidogyne species. J. Nematol. 1990, 22, 10–15. [Google Scholar] [PubMed]
- Adam, M.A.M.; Phillips, M.S.; Blok, V.C. Molecular diagnostic key for identification of single juveniles of seven common and economically important species of root-knot nematode (Meloidogyne spp.). Plant Pathol. 2007, 56, 190–197. [Google Scholar] [CrossRef]
- Rehder, A. Manual of Cultivated Trees and Shrubs; Macmillan Company: New York, NY, USA, 1947. [Google Scholar]
- Chin, S.W.; Shaw, J.; Haberle, R.; Wen, J.; Potter, D. Diversification of almonds, peaches, plums and cherries—Molecular systematics and biogeographic history of Prunus (Rosaceae). Mol. Phylogenet. Evol. 2014, 76, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Eremin, V.G. Genetic potential of species Prunus cerasifera Ehr. and its use in breeding. Acta Hortic. 1978, 74, 61–65. [Google Scholar] [CrossRef]
- Dosba, F.; Bernhard, R.; Zanetto, A. Importance des ressources génétiques des Prunus. C.-R. Acad. Agric. Fr. 1994, 80, 45–57. (In French) [Google Scholar]
- Khallouk, S.; Voisin, R.; Van Ghelder, C.; Engler, G.; Amiri, S.; Esmenjaud, D. Histological Mechanisms of the Resistance Conferred by the Ma Gene Against Meloidogyne incognita in Prunus spp. Phytopathology 2011, 101, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Voisin, R.; Rubio-Cabetas, M.J.; Minot, J.C.; Esmenjaud, D. Penetration, development and emigration of juveniles of the nematode Meloidogyne arenaria in Myrobalan plum (Prunus cerasifera) clones bearing the Ma resistance genes. Eur. J. Plant Pathol. 1999, 105, 103–108. [Google Scholar] [CrossRef]
- Abbott, A.G.; Arus, P.; Scorza, R. Genetic engineering and genomics. In The Peach: Botany, Production and Uses; Wayne, D.R., Bassi, D., Eds.; CAB Editions: New York, NY, USA, 2008; pp. 85–105. [Google Scholar]
- Layne, R.E.C. Peach rootstocks. In Rootstocks for Fruit Crops; Rom, R.C., Carlson, R.F., Eds.; John Wiley and Sons, Inc.: New York, NY, USA, 1987; pp. 185–216. [Google Scholar]
- Yamamoto, T.; Hayashi, T. New root-knot nematode resistance genes and their STS markers in peach. Sci. Hortic. 2002, 96, 81–90. [Google Scholar] [CrossRef]
- Tufts, W.P. Nematode resistance of certain peach seedlings. Proc. Am. Soc. Hortic. Sci. 1929, 26, 98–110. [Google Scholar]
- La Massese, C.S.; Grasselly, C.; Minot, J.C.; Voisin, R. Différence de comportement de 23 clones et hybrides de Prunus à l’égard de quatre espèces de Meloidogyne. Rev. Nematol. 1984, 7, 265–270. [Google Scholar]
- Esmenjaud, D.; Voisin, R.; Van Ghelder, C.; Bosselut, N.; Lafargue, B.; Di Vito, M.; Dirlewanger, E.; Poessel, J.L.; Kleinhentz, M. Genetic dissection of resistance to root-knot nematodes Meloidogyne spp. in plum, peach, almond, and apricot from various segregating interspecific Prunus progenies. Tree Genet. Genomes 2009, 5, 279–289. [Google Scholar] [CrossRef]
- Duval, H.; Van Ghelder, C.; Portier, U.; Confolent, C.; Meza, P.; Esmenjaud, D. New Data Completing the Spectrum of the Ma, RMia, and RMja Genes for Resistance to Root-Knot Nematodes (Meloidogyne spp.) in Prunus. Phytopathology 2019, 109, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, J.H. Plums. In Advances in Fruit Breeding; Janick, J., Moore, J.N., Eds.; Perdue University Press: West Lafayette, IN, USA, 1975; pp. 336–347. [Google Scholar]
- Okie, W.R. Plum rootstocks. In Rootstocks for Fruit Crops; Rom, R.C., Carlson, R.F., Eds.; John Wiley and Sons, Inc.: New York, NY, USA, 1987; pp. 321–360. [Google Scholar]
- Salesses, G.; Grasselly, C.; Renaud, R.; Claverie, J. Les porte-greffe des espèces fruitières à noyau du genre Prunus. In Amélioration des Espèces Végétales Cultivées: Objectifs et Critères de Sélection; Gallais, A., Bannerot, H., Eds.; INRA Editions: Paris, France, 1992; pp. 605–650. [Google Scholar]
- Day, L.H.; Tufts, W.P. Further notes on nematode-resistant rootstocks for deciduous fruit trees. Proc. Am. Soc. Hortic. Sci. 1939, 37, 327–329. [Google Scholar]
- Esmenjaud, D.; Minot, J.C.; Voisin, R.; Pinochet, J.; Simard, M.H.; Salesses, G. Differential response to root-knot nematodes in Prunus species and correlative genetic implications. J. Nematol. 1997, 29, 370–380. [Google Scholar]
- Esmenjaud, D.; Minot, J.C.; Voisin, R.; Bonnet, A.; Salesses, G. Inheritance of resistance to the root-knot nematode Meloidogyne arenaria in Myrobalan plum. Theor. Appl. Genet. 1996, 92, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Lecouls, A.C.; Salesses, G.; Minot, J.C.; Voisin, R.; Bonnet, A.; Esmenjaud, D. Spectrum of the Ma genes for resistance to Meloidogyne spp in Myrobalan plum. Theor. Appl. Genet. 1997, 95, 1325–1334. [Google Scholar] [CrossRef]
- Rubio-Cabetas, M.J.; Minot, J.C.; Voisin, R.; Esmenjaud, D.; Salesses, G.; Bonnet, A. Resistance response of the Ma genes from ‘Myrobalan’ plum to Meloidogyne hapla and M-mayaguensis. Hortscience 1999, 34, 1266–1268. [Google Scholar] [CrossRef] [Green Version]
- Claverie, M.; Bosselut, N.; Lecouls, A.C.; Voisin, R.; Lafargue, B.; Poizat, C.; Kleinhentz, M.; Laigret, F.; Dirlewanger, E.; Esmenjaud, D. Location of independent root-knot nematode resistance genes in plum and peach. Theor. Appl. Genet. 2004, 108, 765–773. [Google Scholar] [CrossRef]
- Verde, I.; Abbott, A.G.; Scalabrin, S.; Jung, S.; Shu, S.Q.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.T.; Grimwood, J.; Cattonaro, F.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–U47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochba, J.; Spiegel-Roy, P. Inheritance of resistance to the root-knot nematode (Meloidogyne iavanica Chitwood) in bitter almond progenies. Euphytica 1975, 24, 453–457. [Google Scholar] [CrossRef]
- Van Ghelder, C.; Lafargue, B.; Dirlewanger, E.; Ouassa, A.; Voisin, R.; Polidori, J.; Kleinhentz, M.; Esmenjaud, D. Characterization of the RMja gene for resistance to root-knot nematodes in almond: Spectrum, location, and interest for Prunus breeding. Tree Genet. Genomes 2010, 6, 503–511. [Google Scholar] [CrossRef]
- Van Ghelder, C.; Esmenjaud, D.; Callot, C.; Dubois, E.; Mazier, M.; Duval, H. Ma Orthologous Genes in Prunus spp. Shed Light on a Noteworthy NBS-LRR Cluster Conferring Differential Resistance to Root-Knot Nematodes. Front. Plant Sci. 2018, 9, 01269. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Wang, L.R.; Zhu, G.R.; Fang, W.C.; Chen, C.W.; Zhao, P. Construction of a Linkage Map and Identification of Resistance Gene Analog Markers for Root-knot Nematodes in Wild Peach, Prunus kansuensis. J. Am. Soc. Hortic. Sci. 2011, 136, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Cao, K.; Wang, L.R.; Zhao, P.; Zhu, G.R.; Fang, W.C.; Chen, C.W.; Wang, X.W. Identification of a candidate gene for resistance to root-knot nematode in a wild peach and screening of its polymorphisms. Plant Breed. 2014, 133, 530–535. [Google Scholar] [CrossRef]
- Maquilan, M.A.D.; Olmstead, M.A.; Dickson, D.W.; Chaparro, J.X. Inheritance of resistance to the peach root-knot nematode (Meloidogyne floridensis) in interspecific crosses between peach (Prunus persica) and its wild relative (Prunus kansuensis). Plant Breed. 2018, 137, 805–813. [Google Scholar] [CrossRef]
- Maquilan, M.A.D.; Olmstead, M.A.; Olmstead, J.W.; Dickson, D.W.; Chaparro, J.X. Genetic analyses of resistance to the peach root-knot nematode (Meloidogyne floridensis) using microsatellite markers. Tree Genet. Genomes 2018, 14, 47. [Google Scholar] [CrossRef]
- Zhu, X.; Xiao, K.; Cui, H.; Hu, J. Overexpression of the Prunus sogdiana NBS-LRR Subgroup Gene PsoRPM2 Promotes Resistance to the Root-Knot Nematode Meloidogyne incognita in Tobacco. Front. Microbiol. 2017, 8, 2113. [Google Scholar] [CrossRef]
- Meyer, A.J. Why is ‘Nemaguard’ rootstock resistant to root-knot nematodes (Meloidogyne spp.) and ‘Kakamas’ susceptible? Deciduous Fruit Grow. 1978, 28, 66–72. [Google Scholar]
- Marull, J.; Pinochet, J.; Felipe, A.; Cenis, J.L. Resistance verification in Prunus selections to a mixture of 13 Meloidogyne isolates and resistance mechanisms of a peach-almond hybrid to M. javanica. Fundam. Appl. Nematol. 1994, 17, 85–92. [Google Scholar]
- Malo, S.E. Nature of resistance of ‘Okinawa’ and ‘Nemaguard’ peach to the root-knot nematode Meloidogyne javanica. Proc. Am. Soc. Hortic. Sci. 1967, 90, 39–46. [Google Scholar]
- Johnson, R. Durable resistance: Definition of genetic control and attainment in plant breeding. Phytopathology 1981, 71, 567–568. [Google Scholar] [CrossRef]
- Khallouk, S.; Voisin, R.; Portier, U.; Polidori, J.; Van Ghelder, C.; Esmenjaud, D. Multiyear Evaluation of the Durability of the Resistance Conferred by Ma and RMia Genes to Meloidogyne incognita in Prunus under Controlled Conditions. Phytopathology 2013, 103, 833–840. [Google Scholar] [CrossRef] [Green Version]
- Esmenjaud, D.; Minot, J.C.; Voisin, R. Effects of durable inoculum pressure and high temperature on root galling, nematode numbers and survival of Myrobalan plum genotypes (Prunus cerasifera Ehr) highly resistant to Meloidogyne spp. Fundam. Appl. Nematol. 1996, 19, 85–90. [Google Scholar]
- Rubio-Cabetas, M.J.; Minot, J.C.; Voisin, R.; Esmenjaud, D. Interaction of root-knot nematodes (RKN) and the bacterium Agrobacterium tumefaciens in roots of Prunus cerasifera: Evidence of the protective effect of the Ma RKN resistance genes against expression of crown gall symptoms. Eur. J. Plant Pathol. 2001, 107, 433–441. [Google Scholar] [CrossRef]
- Stalin, N.; Salesses, G.; Pinochet, J.; Minot, J.C.; Voisin, R.; Esmenjaud, D. Comparative host suitability to Meloidogyne spp. and Pratylenchus vulnus in Myrobalan plum (Prunus cerasifera). Plant Pathol. 1998, 47, 211–215. [Google Scholar] [CrossRef]
- Esmenjaud, D.; Srinivasan, C. Molecular Breeding. In Genetics, Genomics and Breeding of Stone Fruits; Kole, C., Abbott, A.G., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 158–211. [Google Scholar]
- Horn, R.; Sajer, O.; Esmenjaud, D.; Claverie, M.; Dirlewanger, E. Map-Based Cloning of Single Gene Traits and Quantitative Traits. In Genetics, Genomics and Breeding of Stone Fruits; Kole, C., Abbott, A.G., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 212–243. [Google Scholar]
- Lecouls, A.C.; Bergougnoux, V.; Rubio-Cabetas, M.J.; Bosselut, N.; Voisin, R.; Poessel, J.L.; Faurobert, M.; Bonnet, A.; Salesses, G.; Dirlewanger, E.; et al. Marker-assisted selection for the wide-spectrum resistance to root-knot nematodes conferred by the Ma gene from Myrobalan plum (Prunus cerasifera) in interspecific Prunus material. Mol. Breed. 2004, 13, 113–124. [Google Scholar] [CrossRef]
- Lecouls, A.C.; Rubio-Cabetas, M.J.; Minot, J.C.; Voisin, R.; Bonnet, A.; Salesses, G.; Dirlewanger, E.; Esmenjaud, D. RAPD and SCAR markers linked to the Ma1 root-knot nematode resistance gene in Myrobalan plum (Prunus cerasifera Ehr.). Theor. Appl. Genet. 1999, 99, 328–335. [Google Scholar] [CrossRef]
- Claverie, M.; Dirlewanger, E.; Cosson, P.; Bosselut, N.; Lecouls, A.C.; Voisin, R.; Kleinhentz, M.; Lafargue, B.; Caboche, M.; Chalhoub, B.; et al. High-resolution mapping and chromosome landing at the root-knot nematode resistance locus Ma from Myrobalan plum using a large-insert BAC DNA library. Theor. Appl. Genet. 2004, 109, 1318–1327. [Google Scholar] [CrossRef]
- Claverie, M.; Dirlewanger, E.; Bosselut, N.; Van Ghelder, C.; Voisin, R.; Kleinhentz, M.; Lafargue, B.; Abad, P.; Rosso, M.-N.; Chalhoub, B.; et al. The Ma Gene for Complete-Spectrum Resistance to Meloidogyne Species in Prunus Is a TNL with a Huge Repeated C-Terminal Post-LRR Region. Plant Physiol. 2011, 156, 779–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosselut, N.; Van Ghelder, C.; Claverie, M.; Voisin, R.; Onesto, J.P.; Rosso, M.N.; Esmenjaud, D. Agrobacterium rhizogenes-mediated transformation of Prunus as an alternative for gene functional analysis in hairy-roots and composite plants. Plant Cell Rep. 2011, 30, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Van Ghelder, C.; Esmenjaud, D. TNL genes in peach: Insights into the post-LRR domain. BMC Genom. 2016, 17, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mota, A.P.Z.; Fernandez, D.; Arraes, F.B.M.; Petitot, A.S.; de Melo, B.P.; de Sa, M.E.L.; Grynberg, P.; Saraiva, M.A.P.; Guimaraes, P.M.; Brasileiro, A.C.M.; et al. Evolutionarily conserved plant genes responsive to root-knot nematodes identified by comparative genomics. Mol. Genet. Genom. 2020, 295, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Steuernagel, B.; Witek, K.; Jones, J.D.G.; Wulff, B.B.H. MutRenSeq: A Method for Rapid Cloning of Plant Disease Resistance Genes. In Wheat Rust Diseases: Methods and Protocols; Periyannan, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1659, pp. 215–229. [Google Scholar]
- Van Eck, L.; Bradeen, J.M. Hunting for novel Disease Resistance Genes: Observations and Opportunities from the Rosaceae. In Proceedings of the 7th International Symposium on Rose Research and Cultivation, Angers, France, 2–7 July 2017; pp. 125–133. [Google Scholar]
- Saucet, S.B.; Esmenjaud, D.; Van Ghelder, C. Integrity of the Post-LRR Domain Is Required for TIR-NB-LRR Function. Mol. Plant-Microb. Interact. 2021, 34, 286–296. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Root-Knot Nematode Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Resistance Gene | Plant Species | Resistance Source | M. arenaria | M. incognita | M. javanica | M. ethiopica | M. floridensis | M. enterolobii |
| Ma | Plum—P. cerasifera | P.2175 & P.2980 | R | R | R | R | R | R |
| RMia | Peach—P. persica | Nemared | R | R | S | R | S | S |
| RMja | Almond—P. dulcis | Alnem 1 | R | S | R | R | S | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esmenjaud, D. Deciphering Resistance to Root-Knot Nematodes in Prunus for Rootstock Breeding: Sources, Genetics and Characterization of the Ma Locus. Horticulturae 2021, 7, 564. https://doi.org/10.3390/horticulturae7120564
Esmenjaud D. Deciphering Resistance to Root-Knot Nematodes in Prunus for Rootstock Breeding: Sources, Genetics and Characterization of the Ma Locus. Horticulturae. 2021; 7(12):564. https://doi.org/10.3390/horticulturae7120564
Chicago/Turabian StyleEsmenjaud, Daniel. 2021. "Deciphering Resistance to Root-Knot Nematodes in Prunus for Rootstock Breeding: Sources, Genetics and Characterization of the Ma Locus" Horticulturae 7, no. 12: 564. https://doi.org/10.3390/horticulturae7120564
APA StyleEsmenjaud, D. (2021). Deciphering Resistance to Root-Knot Nematodes in Prunus for Rootstock Breeding: Sources, Genetics and Characterization of the Ma Locus. Horticulturae, 7(12), 564. https://doi.org/10.3390/horticulturae7120564