
Antifreeze Protein Improves the Cryopreservation Efficiency of Hosta capitata by Regulating the Genes Involved in the Low-Temperature Tolerance Mechanism


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. AFP Treatment
2.3. RNA Extraction and Gene Expression Analysis
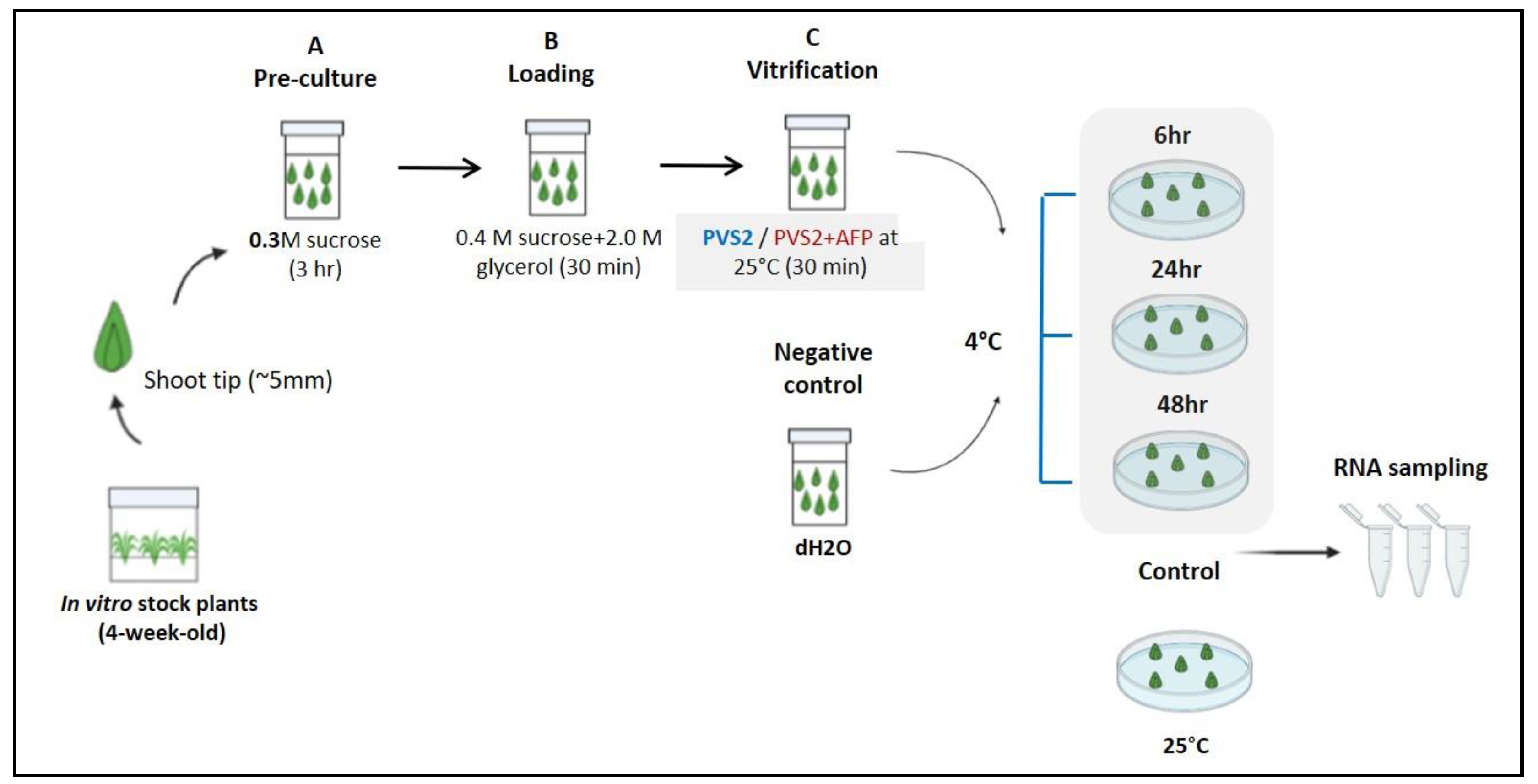
2.4. Cryopreservation
2.5. Recovery of Cryopreserved Meristems
2.6. Statistical Analysis
3. Results
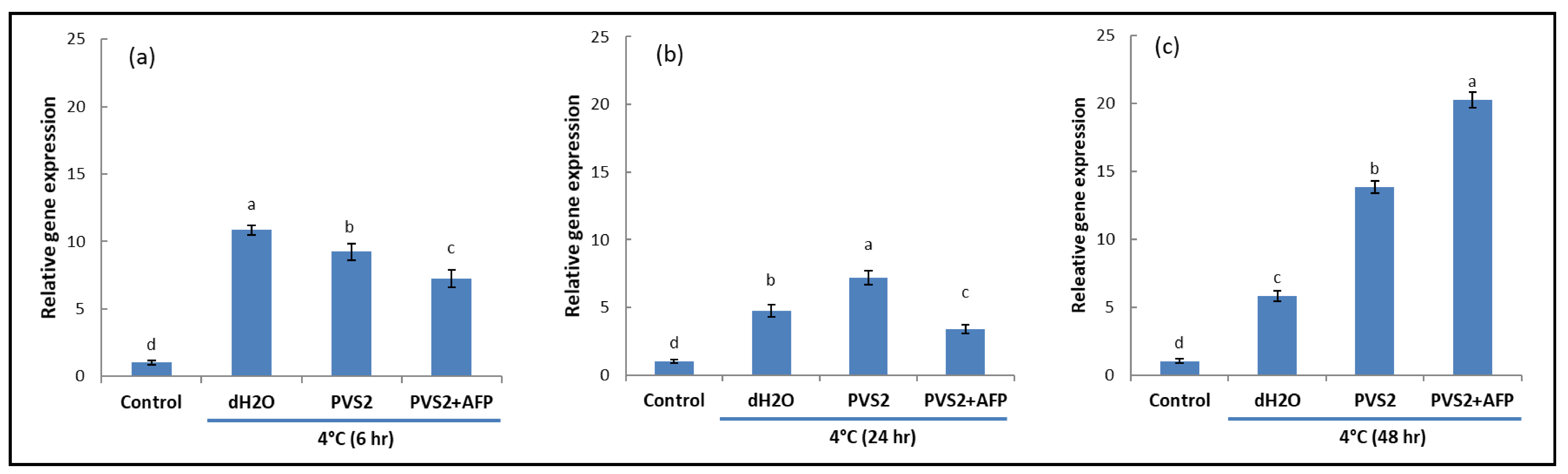
3.1. Antifreeze Protein Regulates Genes Involved in the Low Temperature Tolerance Mechanism
3.2. Influence of Antifree Protein on Cryopreservation Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pe, P.P.W.; Kyu, S.W.; Naing, A.H.; Park, K.I.; Chung, M.Y.; Kim, C.K. Antifreeze proteins promote the germination of low temperature-treated petunia seeds via regulation of antioxidant- and proline-related genes. J. Plant Biotechnol. 2020, 47, 203–208. [Google Scholar] [CrossRef]
- Ryu, K.H.; Park, M.H.; Lee, J.S. Occurrence of mosaic disease of Hosta plants caused by Hosta virus X. Plant Pathol. 2002, 18, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Helliot, B.; Panis, B.; Poumay, Y.; Swennen, R.; Lepoivre, P.; Frison, E. Cryopreservation for the elimination of cucumber mosaic and banana streak viruses from banana (Musa spp.). Plant Cell Rep. 2002, 20, 1117–1122. [Google Scholar] [CrossRef]
- Wang, Q.C.; Mawassi, M.; Li, P.; Gafny, R.; Sela, I.; Tanne, E. Elimination of grapevine virus A (GVA) by cryopreservation of in vitro-grown shoot tips of Vitis vinifera L. Plant Sci. 2003, 165, 321–327. [Google Scholar] [CrossRef]
- Wang, Q.C.; Valkonen, J.P.T. Elimination of two viruses which interact synergistically from sweetpotato by shoot tip culture and cryotherapy. J. Virol. Methods 2008, 154, 135–145. [Google Scholar] [CrossRef]
- Chapsky, L.; Rubinsky, B. Kinetics of antifreeze protein-induced ice growth inhibition. FEBS Lett. 1997, 412, 241–324. [Google Scholar] [CrossRef] [Green Version]
- Arav, A.; Shehu, D.; Mattioli, M. Osmotic and cytotoxic study of vitrification of immature bovine oocytes. J. Reprod Fertil. 1993, 99, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.; Feeney, R.E. Antifreeze proteins: Structures and mechanisms of function. Chem. Rev. 1996, 96, 601–618. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. A brief review of applications of antifreeze proteins in cryopreservation and metabolic genetic engineering. 3 Biotech 2019, 9, 329. [Google Scholar] [CrossRef]
- Wang, J.H.; Bian, H.W.; Zhang, Y.X.; Cheng, H.P. The dual effect of antifreeze protein on cryopreservation of rice (Oryza sativa l.) embryogenic suspension cells. Cryo Lett. 2001, 22, 175–182. [Google Scholar]
- Jeon, S.M.; Naing, A.H.; Park, K.I.; Kim, C.K. The effect of antifreeze protein on the cryopreservation of chrysanthemums. Plant Cell Tissue Organ Cult. 2015, 123, 665–671. [Google Scholar] [CrossRef]
- Seo, J.H.; Naing, A.H.; Jeon, S.M.; Kim, C.K. Anti-freezing-protein type III strongly infuences the expression of relevant genes in cryopreserved potato shoot tips. Plant Mol. Biol. 2018, 97, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Pe, P.P.W.; Naing, A.H.; Park, K.I.; Chung, M.Y.; Kim, C.K. The role of antifreeze proteins in the regulation of genes involved in the response of Hosta capitata to cold. 3 Biotech 2019, 9, 335. [Google Scholar] [CrossRef]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol. 2008, 59, 21. [Google Scholar] [CrossRef]
- Neill, S.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Wilson, I. Nitric oxide evolution and perception. J. Exp. Bot. 2008, 59, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrel, C.; Vazquez, T.; Puyaubert, J.; Rezé, N.; Lesch, M.; Kaiser, W.M.; Dutilleul, C.; Guillas, I.; Zachowski, A.; Baudouin, E. Nitric oxide participates in cold-responsive phosphosphingolipid formation and gene expression in Arabidopsis thaliana. New Phytol. 2011, 189, 415–427. [Google Scholar] [CrossRef]
- Shimoda, Y.; Nagata, M.; Suzuki, A.; Abe, M.; Sato, S.; Kato, T.; Tabata, S.; Higashi, S.; Uchiumi, T. Symbiotic rhizobium and nitric oxide induce gene expression of non-symbiotic hemoglobin in Lotus japonicas. Plant Cell Physiol. 2005, 46, 99–107. [Google Scholar] [CrossRef]
- Corpas, F.J.; Chaki, M.; Fernández-Ocan, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; A del Río, L.; Barroso, J.B. Metabolism of reactive nitrogen species in Pea plants under abiotic stress conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hirai, D.; Sakai, A. Cryopreservation of in vitro-grown axillary shoot-tip meristems of mint (Mentha spicata L.) by encapsulation vitrification. Plant Cell Rep. 1999, 19, 15–155. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. Var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Naing, A.H.; Park, K.I.; Ai, T.N.; Chung, M.Y.; Han, J.S.; Kang, Y.W.; Kim, C.K. Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halmagyi, A.; Deliu, C. Cryopreservation of carnation (Dianthus caryophyllus L.) shoot tips by encapsulation-vitrification. Sci Hort. 2007, 113, 300–306. [Google Scholar] [CrossRef]
- Nikishina, T.V.; Popova, E.V.; Vakhrameeva, M.G.; Varlygina, T.I.; Kolomeitseva, G.L.; Burov, A.V.; Popovich, E.A.; Shirokov, A.I.; Shumilov, V.Y.; Popov, A.S. Cryopreservation of seeds and protocorms of rare temperate orchids. Russ. J. Plant Physiol. 2007, 54, 121–127. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, Y.; Chang, Y.; Reed, B.M. Cryopreservation of Fruit and Ornamental Trees. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Le Bras, C.; Le Besnerais, P.-H.; Hamama, L.; Grapin, A. Cryopreservation of ex-vitro-grown Rosa chinensis ‘Old Blush’ buds using droplet-vitrification and encapsulation-dehydration. Plant Cell Tiss Organ Cult. 2014, 116, 235–242. [Google Scholar] [CrossRef]
- Kyu, S.W.; Naing, A.H.; Pe, P.P.W.; Park, K.I.; Kim, C.K. Tomato seeds pretreated with Antifreeze protein type I (AFP I) promotes the germination under cold stress by regulating the genes involved in germination process. Plant Signal. Behav. 2019, 14, e1682796. [Google Scholar] [CrossRef]
- Cutler, A.J.; Saleem, M.; Kendall, E.; Gusta, L.V.; Georges, F.; Fletcher, G.L. Winter flounder antifreeze protein improves the cold hardiness of plant tissues. J. Plant Physiol. 1989, 135, 351–354. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Cheng, H.; Roberts, S.M.; Yang, Q.; Rhee, S.Y.; Stockinger, E.J. The low temperature-responsive, Solanum CBF1 genes maintain high identity in their upstream regions in a genomic environment undergoing gene duplications, deletions, and rearrangements. Plant Mol. Biol. 2008, 67, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Skinner, J.S.; Meng, R.; Chang, Y.; Bhalerao, N.P.A.; Finn, C.E.; Chen, T.H.H.; Hurry, V. The CBF1-dependent low temperature signalling pathway, regulon and increase in freeze tolerance are conserved in Populus spp. Plant Cell Environ. 2006, 29, 1259–1272. [Google Scholar] [CrossRef]
- Choi, D.W.; Rodriguez, E.M.; Close, T.J. Barley CBF3 gene identification, expression pattern, and map location. Plant Physiol. 2002, 129, 1781–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant J. 2004, 39, 905–919. [Google Scholar] [CrossRef]
- Bhattarai, T.; Fettig, S. Isolation and characterization of a dehydrin gene from Cicer pinnatifidum, a drought-resistant wild relative of chickpea. Physiol. Plant. 2005, 123, 452–458. [Google Scholar] [CrossRef]
- Welling, A.; Rinne, P.; Viherä-Aarnio, A.; Kontunen-Soppela, S.; Heino, P.; Palva, E.T. Photoperiod and temperature differentially regulate the expression of two dehydrin genes during overwintering of birch (Betula pubescens Ehrh.). J. Exp. Bot. 2004, 55, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Baudo, M.M.; Meza-Zepeda, L.A.; Palva, E.T.; Heino, P. Induction of homologous low temperature and ABA-responsive genes in frost-resistant (Solanum commersonii) and frost-sensitive (Solanum tuberosum cv. Bintje) potato species. Plant Mol. Biol. 1996, 30, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′→ 3′) | PCR Condition |
|---|---|---|
| CBF1 | F- AGGGTCAAAGGACACACGTC R- GGGAGCACACGGAGTTTTTG | 95 °C (10 min) → [95 °C (15 s) → 59 °C (30 s)] × 40 cycles → 95 °C (15 s) → 58 °C (1 min) → 95 °C (15 s) |
| DHN1 | F- CAGTCTGACATCACCAGGGTA R- GGAGTTATAGGAGAGACAGATT | 95 °C (10 min) → [95 °C (15 s) → 59 °C (30 s)] × 40 cycles → 95 °C (15 s) → 57 °C (1 min) → 95 °C (15 s) |
| Nia1 | F- CCACCAGGAGAAACCGAACA R- TTGAAAGACTCGTCCCAGGC | 95 °C (10 min) → [95 °C (15 s) → 59 °C (30 s)] × 40 cycles → 95 °C (15 s) → 55 °C (1 min) → 95 °C (15 s) |
| Nia2 | F- CCTTCTTTGGTAGACGCCGA R- TCGTGACATGGCGTCGTAAT | 95 °C (10 min) → [95 °C (15 s) → 59 °C (30 s)] × 40 cycles → 95 °C (15 s) → 53 °C (1 min) → 95 °C (15 s) |
| Treatment | Survival Rate (%) | Shoot Regeneration Rate (%) |
|---|---|---|
| Control | 0c | 0c |
| PVS2 | 33.3b | 17.8b |
| PVS2+AFP | 46.7a | 28.9a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pe, P.P.W.; Naing, A.H.; Kim, C.K.; Park, K.I. Antifreeze Protein Improves the Cryopreservation Efficiency of Hosta capitata by Regulating the Genes Involved in the Low-Temperature Tolerance Mechanism. Horticulturae 2021, 7, 82. https://doi.org/10.3390/horticulturae7040082
Pe PPW, Naing AH, Kim CK, Park KI. Antifreeze Protein Improves the Cryopreservation Efficiency of Hosta capitata by Regulating the Genes Involved in the Low-Temperature Tolerance Mechanism. Horticulturae. 2021; 7(4):82. https://doi.org/10.3390/horticulturae7040082
Chicago/Turabian StylePe, Phyo Phyo Win, Aung Htay Naing, Chang Kil Kim, and Kyeung Il Park. 2021. "Antifreeze Protein Improves the Cryopreservation Efficiency of Hosta capitata by Regulating the Genes Involved in the Low-Temperature Tolerance Mechanism" Horticulturae 7, no. 4: 82. https://doi.org/10.3390/horticulturae7040082
APA StylePe, P. P. W., Naing, A. H., Kim, C. K., & Park, K. I. (2021). Antifreeze Protein Improves the Cryopreservation Efficiency of Hosta capitata by Regulating the Genes Involved in the Low-Temperature Tolerance Mechanism. Horticulturae, 7(4), 82. https://doi.org/10.3390/horticulturae7040082