
A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and FA Treatment
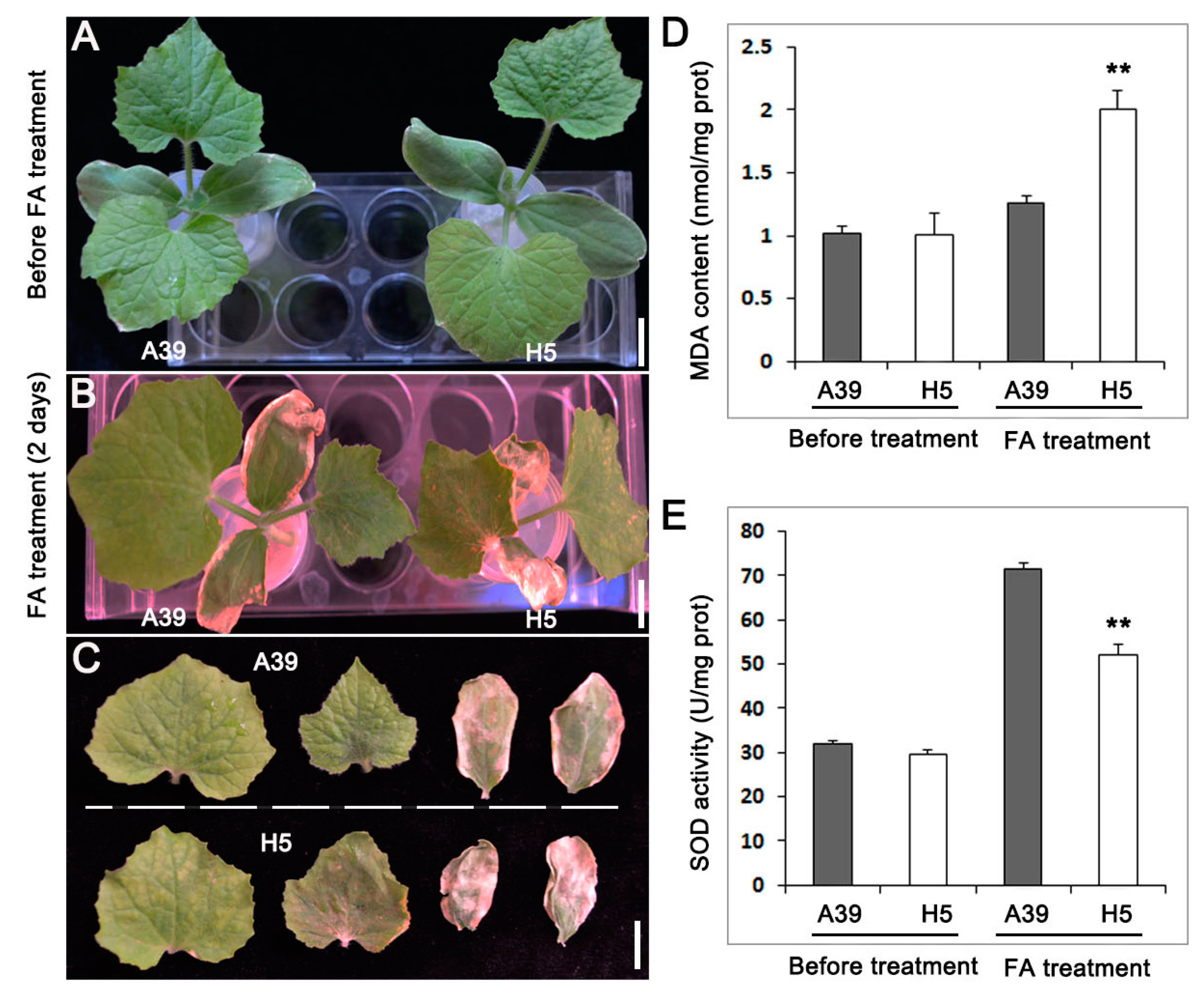
2.2. Measurement of Malondialdehyde (MDA) Content and Superoxide Dismuatse (SOD) Activity
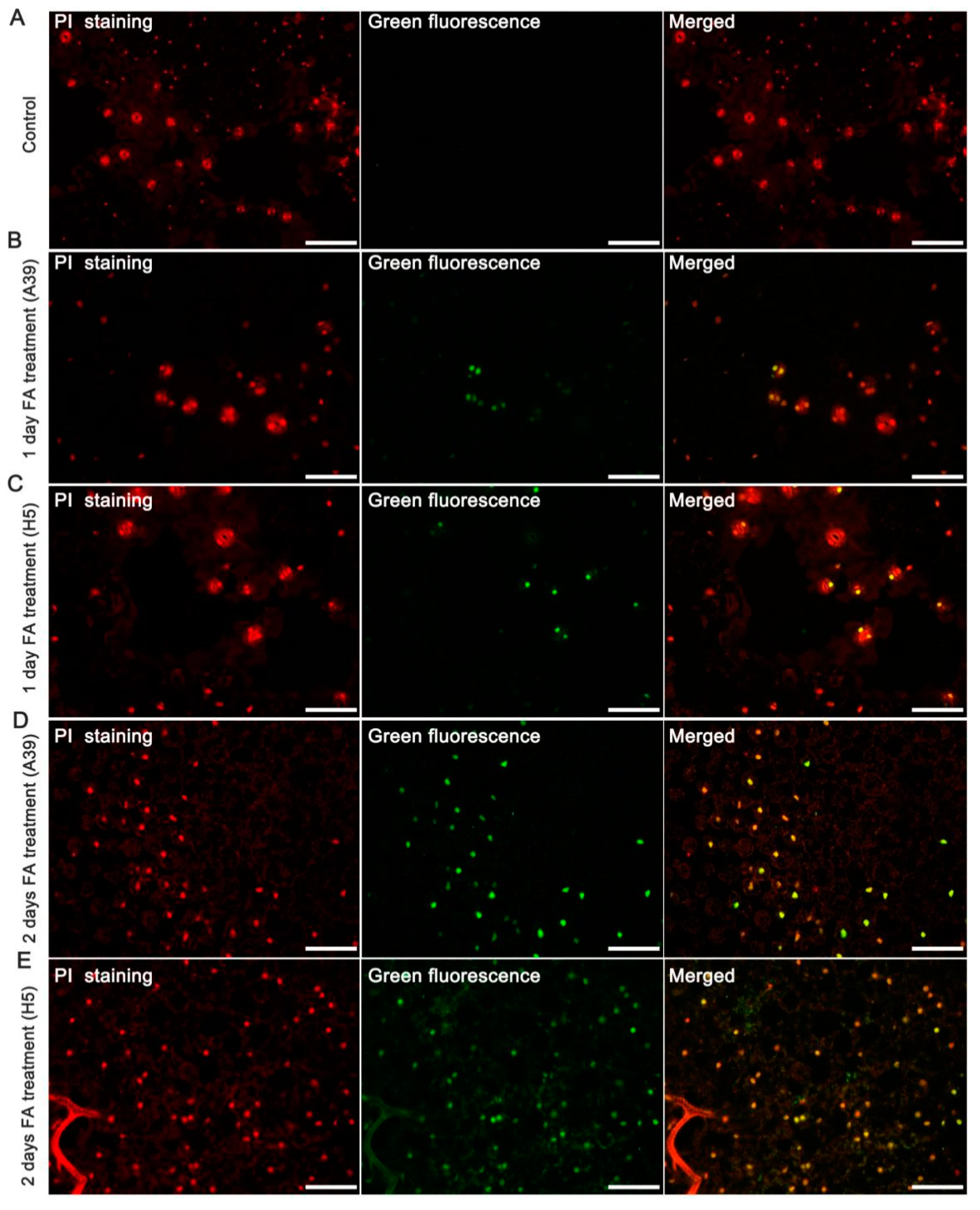
2.3. Terminal Deoxynucleotidyl Transferase-Mediated UTP Nick-End Labeling (TUNEL) Assay
2.4. Transcriptome Sequencing
2.5. Screening and Significant Test for DEGs
2.6. Quantitative Real-Time PCR Analysis
2.7. Statistical Analyses
3. Results
3.1. Phenotype of ‘A39’ and ‘H5’ upon FA Stress
3.2. Cell Death under FA Stress
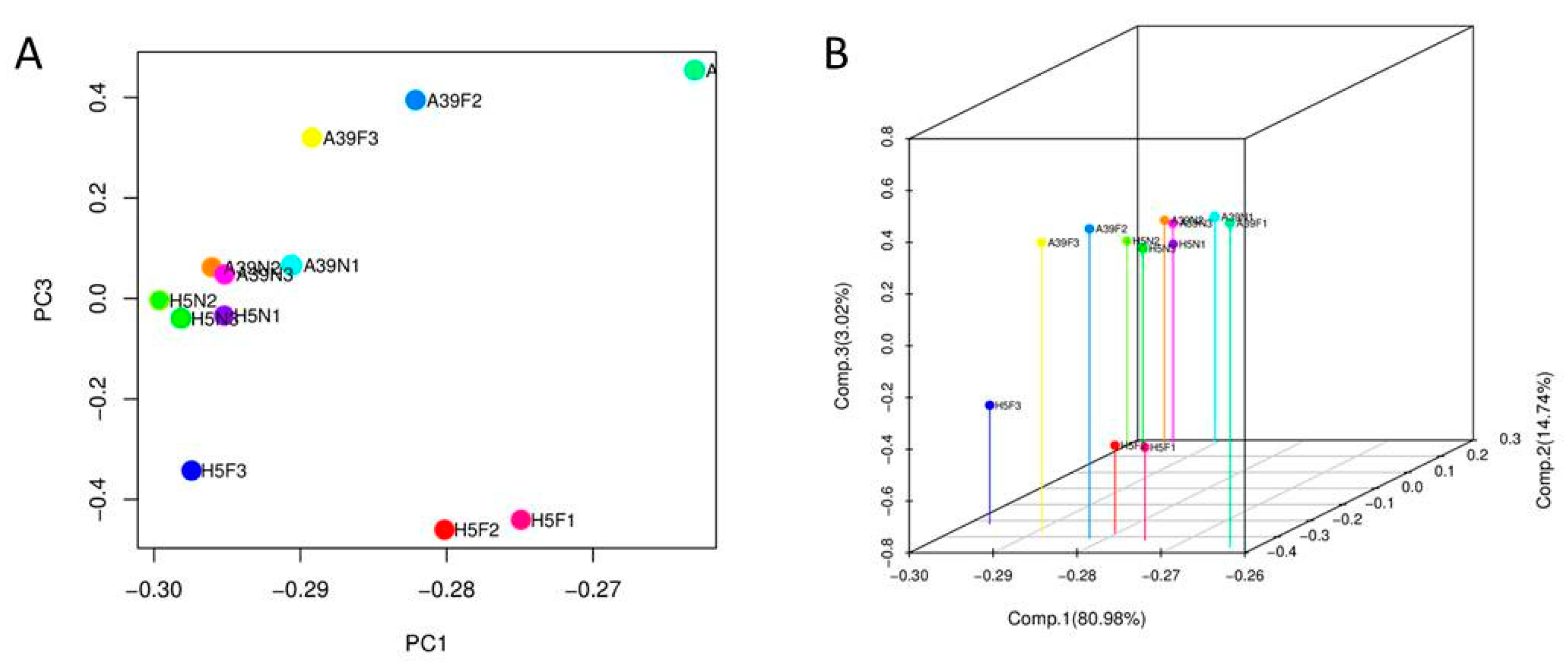
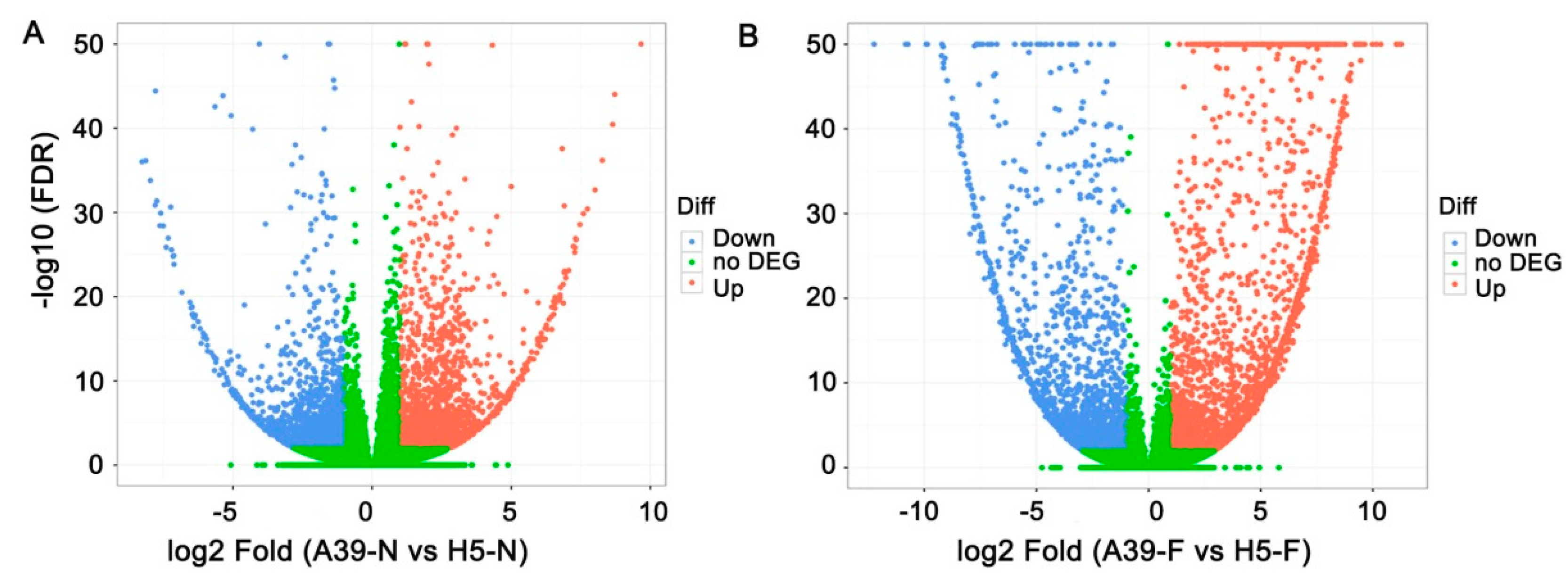
3.3. Transcripts Assembly and SNP Analysis
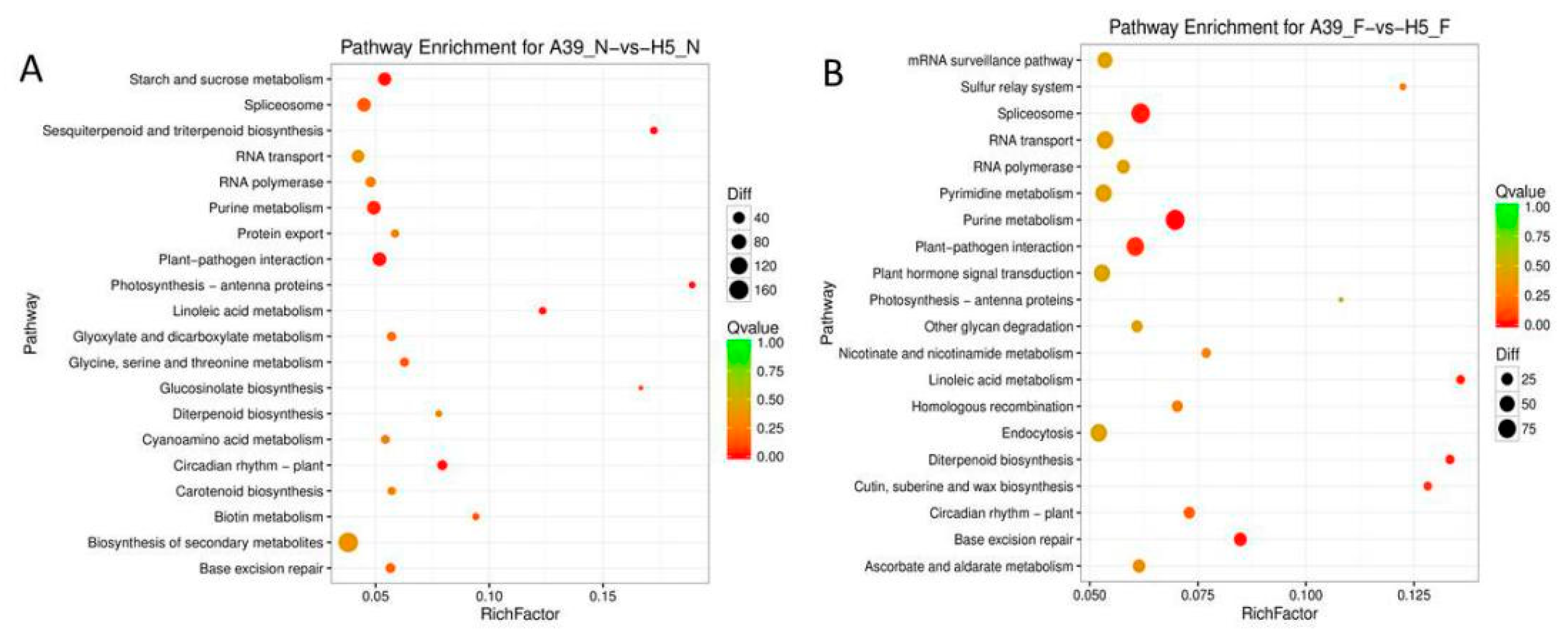
3.4. Functional Classification of FA Stress Response Genes
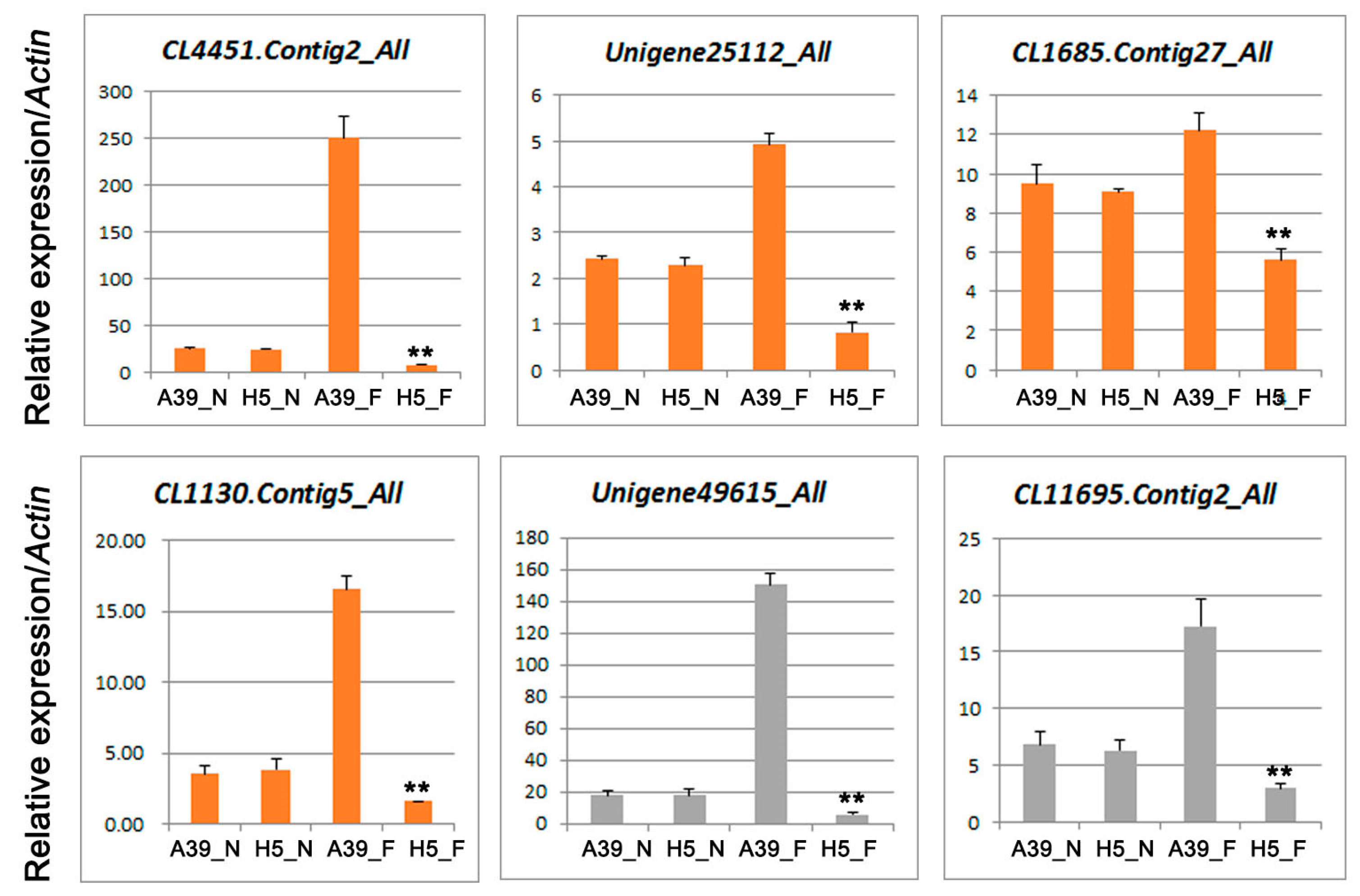
3.5. Expression of Genes Encoding Pathogen-Related Proteins
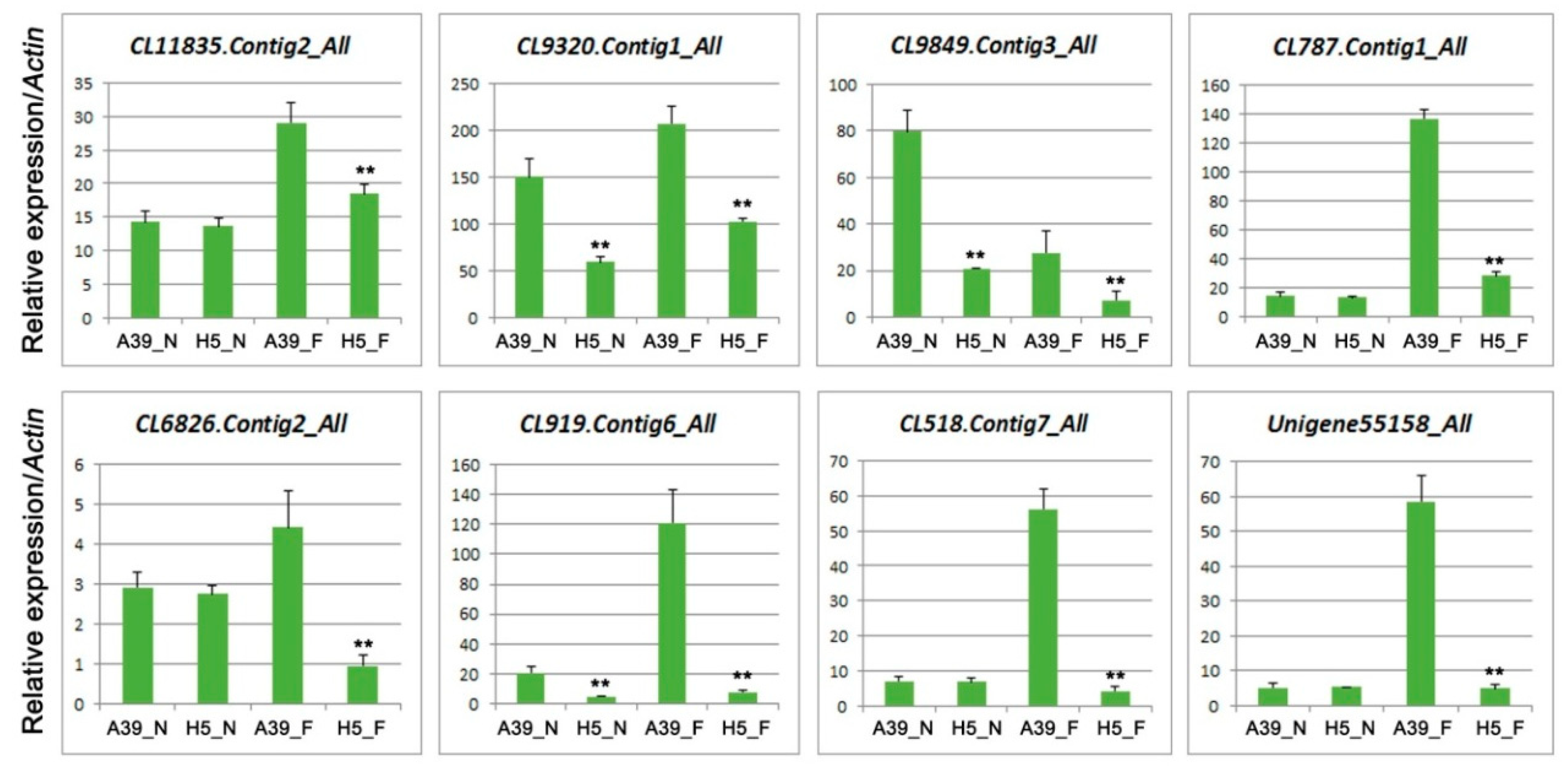
3.6. Expression of Genes Related to Ethylene (ET)
4. Discussion
4.1. More Apoptosis were Detected in ‘H5’
4.2. Analysis of Pathogen-Related (PR) Protein under FA Stress
4.3. Analysis of Genes Related to Enzymes under FA Stress
4.4. Analysis of Genes Related to Hormones under FA Stress
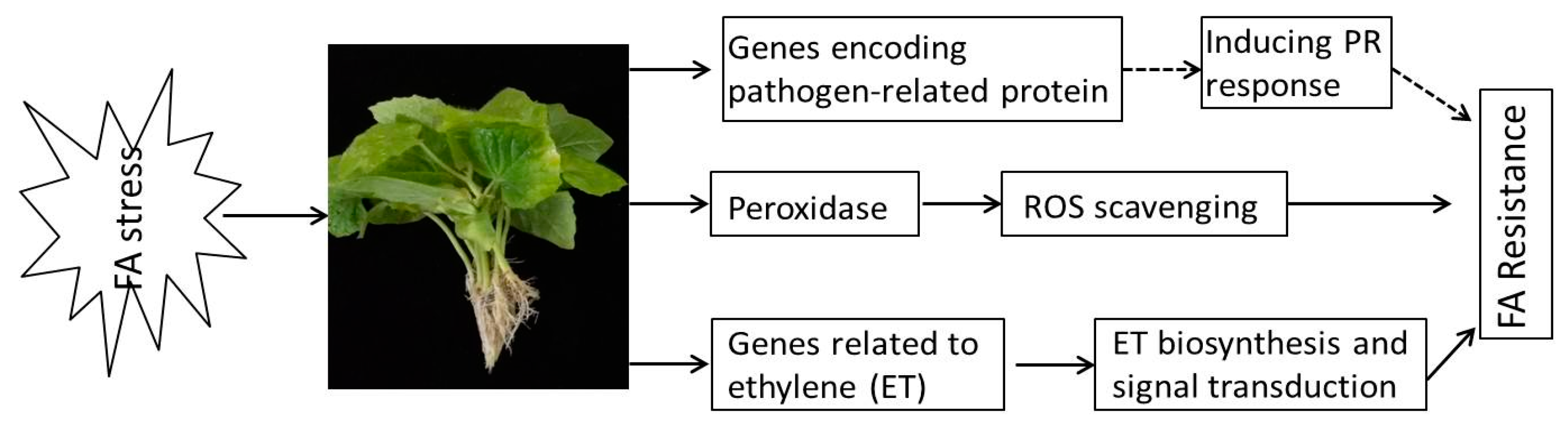
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, X.M.; Xie, D.S.; Chen, Q.H.; Peng, Q.W. Chieh-qua biotechnology, progress and prospects. Asia and Aust. J. Sci. Biotechnol. 2007, 1, 19–22. [Google Scholar]
- Xie, D.S.; Xu, Y.C.; Wang, J.P.; Liu, W.R.; Zhou, Q.; Luo, S.B.; Huang, W.; He, X.M.; Li, Q.; Peng, Q.W.; et al. The wax gourd genomes offer insights into the genetic diversity and ancestral cucurbit karyotype. Nat. Commun. 2019. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Deng, M.G.; Zhu, T.S.; Luo, F.F.; Xie, S.D. Study on the regularity and comprehensive control of the fusarium wilt in chieh-qua. Guangdong Agric. Science 1994, 6, 31–34. (In Chinese) [Google Scholar]
- Owen, J.H. Fusarium wilt of cucumber. Phytopathology 1995, 45, 435–439. [Google Scholar]
- Gäumann, E. Fusaric acid as a wilt toxin. Phytopathology 1957, 47, 342–357. [Google Scholar]
- Gäumann, E. The mechanism of fusaric acid injury. Phytopathology 1958, 48, 670–686. [Google Scholar]
- Harborne, J.B. Introduction to Ecological Biochemistry; Academic Press: London, UK, 1989; Volume 338. [Google Scholar]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.; Guo, S. Fusaric acid is acrucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef]
- Liu, X.J.; Huang, B.B.; Lin, J.; Fei, J.; Chen, Z.; Pang, Y.; Sun, X.; Tang, K. A novel pathogenesis-related protein (SsPR10) from Solanum surattense with ribonucleolytic and antimicrobial activity is stress and pathogen-inducible. J. Plant Physiol. 2006, 163, 546–556. [Google Scholar] [CrossRef]
- Sapko, O.A.; Utarbaeva, A.S.; Makulbek, S. Effect of fusaric acid on prooxidantand antioxidant properties of the potato cell suspension culture. Russ. J. Plant Physiol. 2011, 58, 828–835. [Google Scholar]
- Wu, H.; Yin, X.; Liu, D.; Ling, N.; Bao, W.; Ying, R.; Zhu, Y.; Guo, S.; Shen, Q. Effect offungal fusaric acid on the root and leaf physiology of watermelon (Citrullus lanatus) seedlings. Plant Soil. 2008, 308, 255–266. [Google Scholar] [CrossRef]
- He, X.; Xie, D.; Peng, Q.; Mu, L. Studies on the somaclonalvariants with resistance to fusaric acid in Chieh-qua (Benincasa hispida cogn. var. chieh-qua how). Chin. Agric. Sci Bull. 2009, 25, 172–175. (In Chinese) [Google Scholar]
- Mao, Y.Z.; Jiang, B.; Peng, Q.W.; Liu, W.R.; Lin, Y.; Xie, D.S.; He, X.M.; Li, S.S. Cloning and characterization of WRKY gene homologs in Chieh-qua (Benincasa hispida Cogn. var. Chieh-qua How) and their expression in response to fusaric acid treatment. Biotech 2017, 7, 86. [Google Scholar]
- Martínez-Medina, A.; Pascual, J.A.; Francisco, P.; Pérez-Alfocea, F.; Albacete, A.; Roldán, A. Trichoderma harzianum and Glomus intraradices modify the hormone disruption induced by Fusarium oxysporum infection in melon plants. Phytopathology 2010, 100, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.Y.; Deng, G.M.; Yang, J.; Altus, V.; Jin, Y.; Kuang, R.B.; Zou, C.W.; Lv, Z.C.; Yang, Q.S.; Sheng, O.; et al. Transcriptome profiling of resistant and susceptible Cavendish banana roots following inoculation with Fusarium oxysporum f. sp. Cubensetropical race 4. BMC Genom. 2012, 13, 374. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.P.; Miao, H.; Yang, Y.H.; Xie, B.Y.; Wang, Y.; Gu, X.F. A major quantitative trait locus conferring resistance to fusarium wilt was detected in cucumber by using recombinant inbred lines. Mol. Breed. 2014, 34, 1805–1815. [Google Scholar] [CrossRef]
- Xu, Y.; Ouyang, X.X.; Zhang, H.Y.; Kang, G.B.; Wang, Y.L.; Chen, H. Identification of a RAPD marker linked to Fusarium wilt resistant gene in wild watermelon germplasm (Citrullus lanatus var. citroides). Acta Bot. Sin. 1999, 49, 952–955. [Google Scholar]
- Zhang, Y.; Zhang, H.Y.; Guo, S.G.; Ren, Y.; Zhang, J.; Geng, L.H.; Liang, Z.H.; Xu, Y. Developments of molecular markers tightly linked to Fon-1 for resistance to Fusarium oxysporum f.sp.niveum Race 1 in watermelon. Sci. Agric. Sin. 2013, 46, 2085–2093. [Google Scholar]
- Xie, D.S.; He, X.M.; Peng, Q.W.; Li, H.P. Identification of RAMP marker linked to the blight resistance gene in winter Gourd and its application. Mol. Plant Breed. 2009, 7, 928–933. (In Chinese) [Google Scholar]
- Xie, D.S.; He, X.M.; Peng, Q.W.; Li, H.P. Inheritance of resistance to fusariumwilt in winter melon. Chin. J. Trop. Crop 2009, 30, 1005–1008. (In Chinese) [Google Scholar]
- Li, Z.L.; Qiao, Y.C.; Lin, J.Y.; Li, G.G. Studies on molecular markers to fusarium wilt resistance gene in chieh-qua. Genom. Appl. Biol. 2015, 34, 001946–1949. (In Chinese) [Google Scholar]
- Xing, M.M.; Lv, H.H.; Ma, J.; Xu, D.H.; Li, H.L.; Yang, L.M.; Kang, J.G.; Wang, X.W.; Fang, Z.Y.; Yu, J.H. Transcriptome profiling of resistance to Fusarium oxysporum f. sp. conglutinans in cabbage (Brassica oleracea) roots. PloS ONE 2016, 11, e0148048. [Google Scholar] [CrossRef]
- Gaige, A.R.; Ayella, A.; Bin, S. Methyl jasmonate and ethylene induce partial resistance in Medicago truncatula against the charcoal rot pathogen Macrophomina phaseolina. Physiol. Mol. Plant Pathol. 2010, 74, 412–418. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, P.; Cui, F.; Zhang, F.; Luo, X.; Xie, J. Transcriptome analysis of salt stress responsiveness in the seedlings of Dongxiang wild rice (Oryza rufipogon Griff.). PLoS ONE 2016, 11, e0146242. [Google Scholar]
- Xiao, J.; Jin, X.H.; Jia, X.P.; Wang, H.Y.; Cao, A.Z.; Zhao, W.P.; Pei, H.Y.; Xue, Z.K.; He, L.Q.; Chen, Q.G.; et al. Transcriptome-based discovery of pathwaysand genes related to resistance against Fusarium head blight in wheat landrace Wangshuibai. BMC Genom. 2013, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Lyons, R.; Stiller, J.; Powell, J.; Rusu, A.; Manners, J.M.; Kazan, K. Fusariumoxysporum triggers tissue-specific transcriptional reprogramming in Arabidopsis thaliana. PLoS ONE 2015, 10, 1–23. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, B.; Liu, W.R.; Lin, Y.E.; Liang, Z.J.; He, X.M.; Peng, Q.W. Transcriptome analyses provide novel insights into heat stress responses in chieh-qua (Benincasa hispida Cogn. var. Chieh-Qua How). Int. J. Mol. Sci. 2019, 20, 883. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; He, X.M.; Jiang, B.; Liu, W.R.; Lin, Y.E.; Xie, D.S.; Liang, Z.J.; Chen, L.H.; Peng, Q.W. Transcriptome analysis in different chieh-qua cultivars provides new insights into drought-stress response. Plant Biotechnol. Rep. 2019. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. Wego, a web tool for plotting go annotations. Nucleic Acids Res. 2006, 34, W293. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. Kegg for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accuratenormalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal controlgenes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuc, J. Induced immunity to plant disease. Bioscience 1982, 32, 854–860. [Google Scholar]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487. [Google Scholar] [CrossRef]
- Ouyang, B.; Chen, Y.H.; Li, H.X.; Qian, C.J.; Huang, S.L.; Ye, Z.B. Transformation of tomatoes with osmotin and chitinase genes and their resistance to\r, Fusarium\r, wilt. J. Hortic. Sci. Biotechnol. 2005, 80, 517–522. [Google Scholar] [CrossRef]
- O’Donnell, P.J.; Schmelz, E.; Block, A.; Miersch, O.; Wasternack, C.; Jones, J.B.; Klee, H.J. Multiple hormones act sequentially to mediate a susceptible tomato pathogen defense response. Plant Physiol. 2003, 133, 1181–1189. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P.; Penninckx, I.A.; Cammue, B.P.; Broekaert, W.F. The complexity of disease signaling in Arabidopsis. Curr. Opin. Immunol. 2001, 13, 63–68. [Google Scholar] [CrossRef]
- Jiang, B.; Liu, W.R.; Xie, D.S.; Peng, Q.W.; He, X.M.; Lin, Y.E.; Liang, Z.J. High-density genetic map construction and gene mapping of pericarp color in wax gourd using specific-locus amplified fragment (SLAF) sequencing. BMC Genom. 2015, 16, 1035. [Google Scholar] [CrossRef] [Green Version]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1989, 186, 421–431. [Google Scholar]
- Galindo-Gonzalez, L.; Deyholos, M.K. RNA-seq Transcriptome response of flax (Linum Usitatissimum, L.) to the pathogenic fungus Fusarium oxysporum f. Sp. lini. Front Plant Sci. 2016, 7, 1766. [Google Scholar] [CrossRef] [Green Version]
- Deslandes, L.; Olivier, J.; Theulieres, F.; Hirsch, J.; Feng, D.X.; Bittner-Eddy, P. Resistance to Ralstonia solanacearum in Arabidopsis thaliana is conferred by the recessive RRS1-R gene, a member of a novel family of resistance genes. Proc. Natl. Acad. Sci. USA 2002, 99, 2404–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Park, C.J.; Kim, K.J.; Shin, R.; Park, J.M.; Shin, Y.C.; Paek, K.H. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant J. 2004, 37, 186–198. [Google Scholar] [CrossRef]
- Hashimoto, M.; Kisseleva, L.; Sawa, S.; Furukawa, T.; Komatsu, S.; Koshiba, T. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway. Plant Cell Physiol. 2004, 45, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadha, P.; Das, R.H. A pathogenesis related protein, AhPR10 from peanut, an insight of its mode of antifungal activity. Planta 2006, 225, 213–222. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Ron, M. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.H.; Wang, M.; Hu, J.B.; Wang, W.; Fu, X.Z.; Liu, J.H. PtrABF of Poncirus trifoliata functions in dehydration tolerance by reducing stomatal density and maintaining reactive oxygen species homeostasis. J. Exp. Bot. 2015, 19, 19. [Google Scholar]
- Wei, T.L.; Wang, Y.; Xie, Z.Z.; Guo, D.Y.; Chen, C.W.; Fan, Q.J.; Deng, X.D.; Liu, J.H. Enhanced ROS scavenging and sugar accumulation contribute to drought tolerance of naturally occurring autotetraploids in Poncirus trifoliata. Plant Biotechnol. J. 2019, 17, 1394–1407. [Google Scholar] [CrossRef] [Green Version]
- Hiraga, S. A large family of class III plant peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindschedler, L.V.; Dewdney, J.; Blee, K.A.; Stone, J.M.; Asai, T.; Plotnikov, J.; Denoux, C.; Hayes, T.; Gerrish, C.; Davies, D.R.; et al. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. 2006, 47, 851–863. [Google Scholar] [CrossRef] [Green Version]
- Johrde, A.; Schweizer, P. A class III peroxidase specifically expressed in pathogen-attacked barley epidermis contributes to basal resistance. Mol. Plant Pathol. 2008, 9, 687–696. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Kaleem, F.; Shabir, G.; Aslam, K.; Rasul, S.; Manzoor, H.; Shah, S.M.; Khan, A.R. An overview of the genetics of plant response to salt stress: Present status and the way forward. Appl. Biochem. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 10, 2839–2856. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.N. SA, JA, ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1998, 1, 316–323. [Google Scholar] [CrossRef]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [Green Version]
- Broekaert, W.F.; Delauré, S.L.; DeBolle, M.F.; Cammue, B.P. The role of ethylene in host-pathogen interactions. Annu. Rev. Phytopathol. 2006, 44, 393–416. [Google Scholar] [CrossRef]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Qi, L.; Liu, X.; Cai, S.; Zhang, Z. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen rhizoctonia cerealis and freezing stresses. Plant Physiol. 2014, 164, 1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; De Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.W.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar] [PubMed] [Green Version]
- Penninckx, I.A.; Thomma, B.P.; Buchala, A.; Métraux, J.P.; Broekaert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 10, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano, R.; Stepanova, A.; Chao, Q.; Ecker, J.R. Nuclear events in ethylene signaling: A transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENERESPONSE FACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | log2 Fold | p Value | Diff | Swissprot-Annotation |
|---|---|---|---|---|
| CL4451.Contig2 | 6.815745035 | 1.04 × 10−86 | Up | Pathogen-related protein |
| Unigene25112 | 2.735795316 | 4.43 × 10−8 | Up | Pathogen-related protein-PRB1-2 |
| CL1685.Contig27 | 1.178656927 | 0.0000999 | Up | Pathogen-related protein PR-4A |
| CL1130.Contig5 | 1.550394333 | 0.0003625 | Up | Pathogen-related protein |
| Unigene49615 | 1.312624296 | 8.91 × 10−8 | Up | Peroxidase |
| CL11695.Contig2 | 2.4597268 | 0.00000841 | Up | Peroxidase |
| Gene ID | log2 Fold | p Value | Diff | Nr-Annotation |
|---|---|---|---|---|
| CL11835.Contig2_All | 1.032445303 | 4.54 × 10−5 | Up | salicylic acid-binding protein |
| CL9320.Contig1_All | 5.243998483 | 1.15 × 10−145 | Up | ethylene-responsive transcription factor RAP2-4-like |
| CL9849.Contig3_All | 2.13936557 | 1.16 × 10−20 | Up | ETHYLENE INSENSITIVE 3-like 1 protein isoform X1 |
| CL787.Contig1_All | 1.23793966 | 7.8 × 10−18 | Up | ACC oxidase |
| CL6826.Contig2_All | 1.241786521 | 1.34 × 10−13 | Up | AP2-like ethylene-responsive transcription factor AIL1 |
| CL919.Contig6_All | 4.333608032 | 2.53 × 10−13 | Up | ethylene-responsive transcription factor SHINE 2-like |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Liu, W.; Yan, J.; Sun, P.; Chen, F.; Jiang, B.; Xie, D.; Lin, Y.; Peng, Q.; He, X. A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress. Horticulturae 2021, 7, 88. https://doi.org/10.3390/horticulturae7040088
Wang M, Liu W, Yan J, Sun P, Chen F, Jiang B, Xie D, Lin Y, Peng Q, He X. A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress. Horticulturae. 2021; 7(4):88. https://doi.org/10.3390/horticulturae7040088
Chicago/Turabian StyleWang, Min, Wenrui Liu, Jinqiang Yan, Piaoyun Sun, Feng Chen, Biao Jiang, Dasen Xie, Yu’e Lin, Qingwu Peng, and Xiaoming He. 2021. "A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress" Horticulturae 7, no. 4: 88. https://doi.org/10.3390/horticulturae7040088
APA StyleWang, M., Liu, W., Yan, J., Sun, P., Chen, F., Jiang, B., Xie, D., Lin, Y., Peng, Q., & He, X. (2021). A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress. Horticulturae, 7(4), 88. https://doi.org/10.3390/horticulturae7040088