
Superior Line from Anther Culture of Dendrocalamus latiflorus Selected after Field Trial


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Materials
2.3. Field Experiment
2.4. Nutrient Comparison of Bamboo Shoots
2.5. Anatomical Comparison of Bamboo Culms
2.6. Detection of Chromosome Ploidy in Plant Materials
2.7. Data Statistics and Analysis
3. Results
3.1. Survival Rate of Regenerated Lines in Field
3.2. Difference of Growth Characters in Regenerated Lines after One Year of Planting
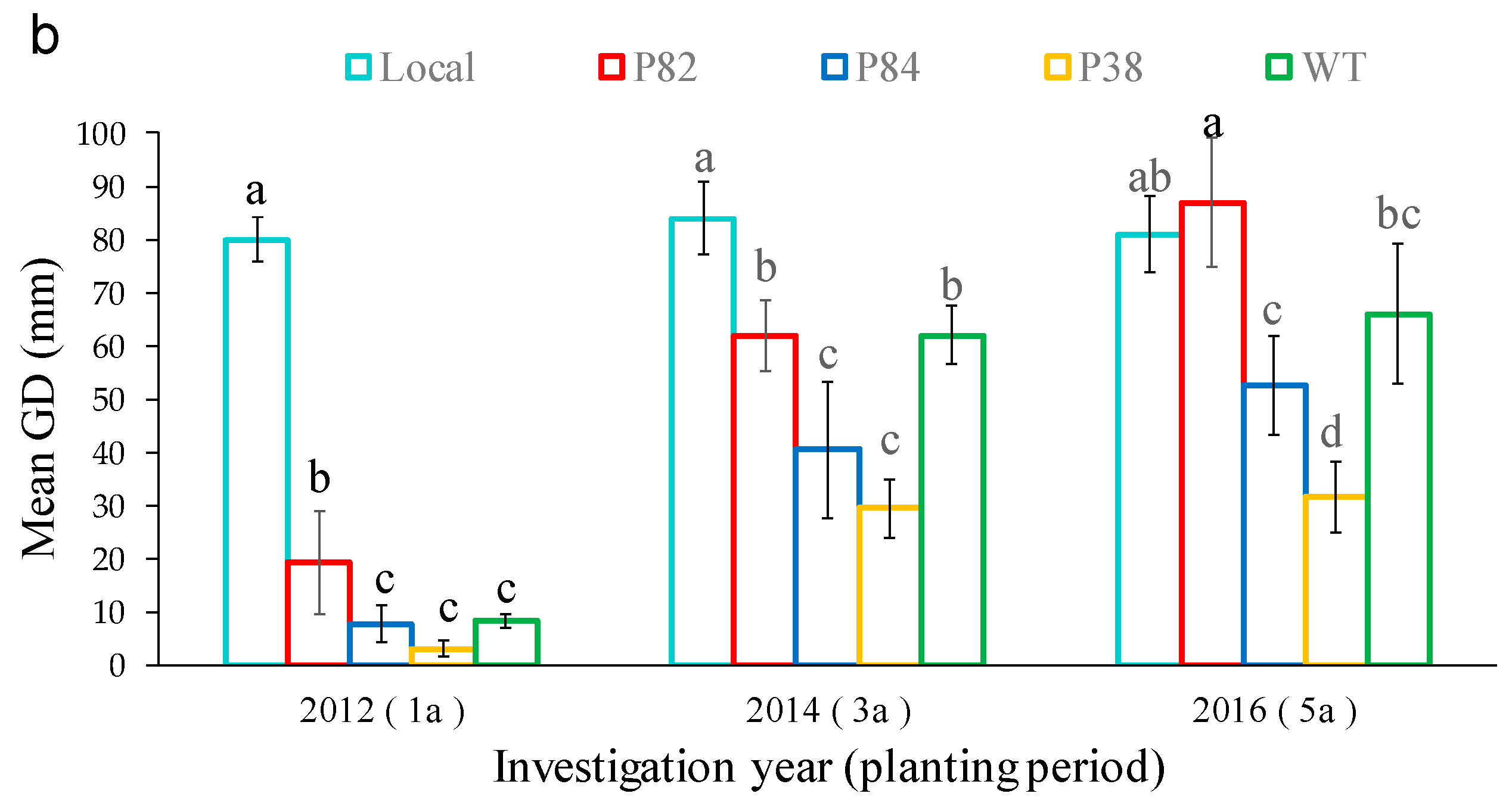
3.3. Differences in Growth Characters of Regenerated Lines after three Years of Growth and Selection for Superior Regenerated Lines
3.4. Comparison the Shoot’ Yield of Superior Lines
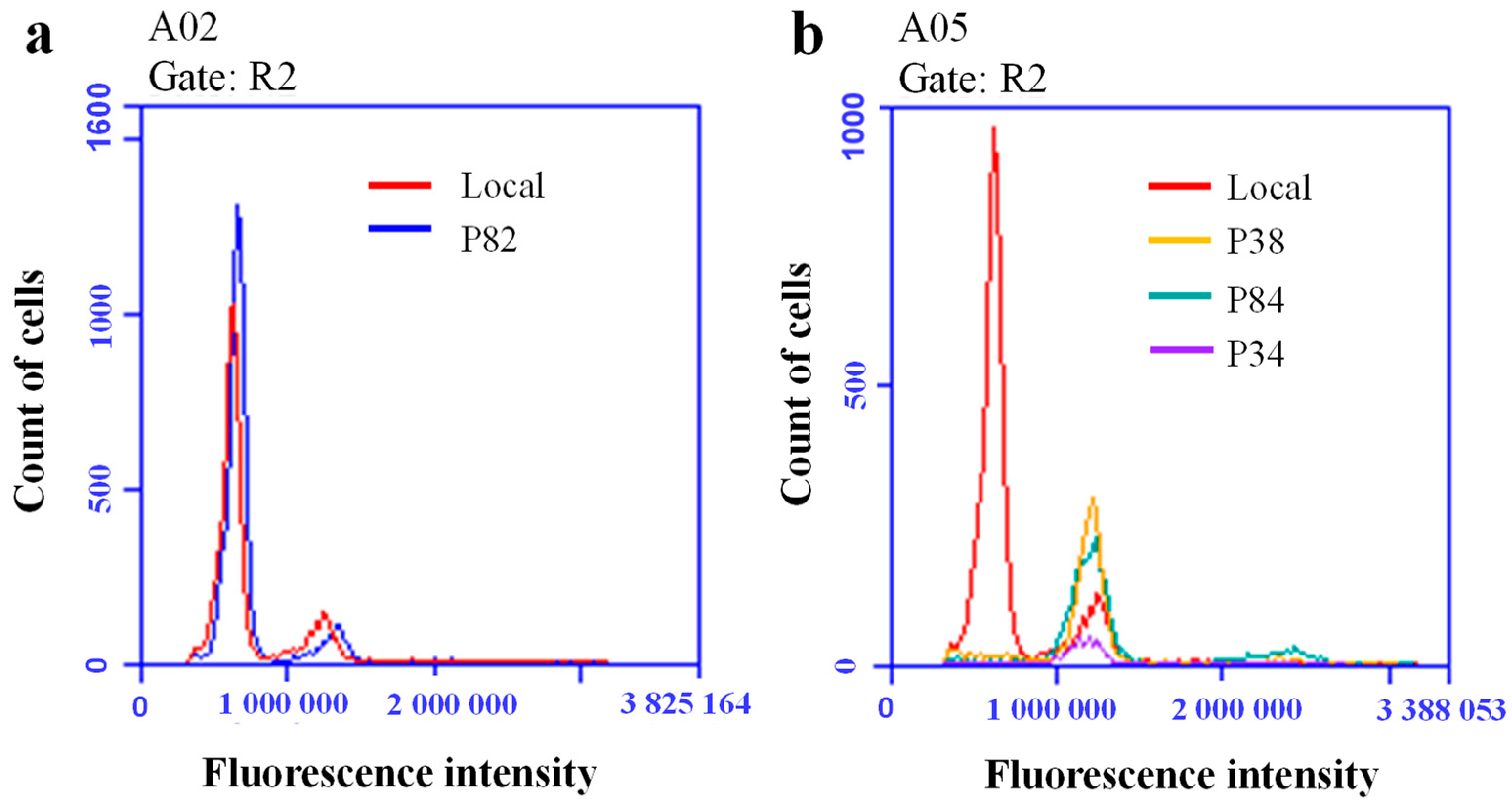
3.5. Ploidy Level Test of Superior Lines P82
3.6. Comparison the Shoot’ Nutrient of Superior Lines
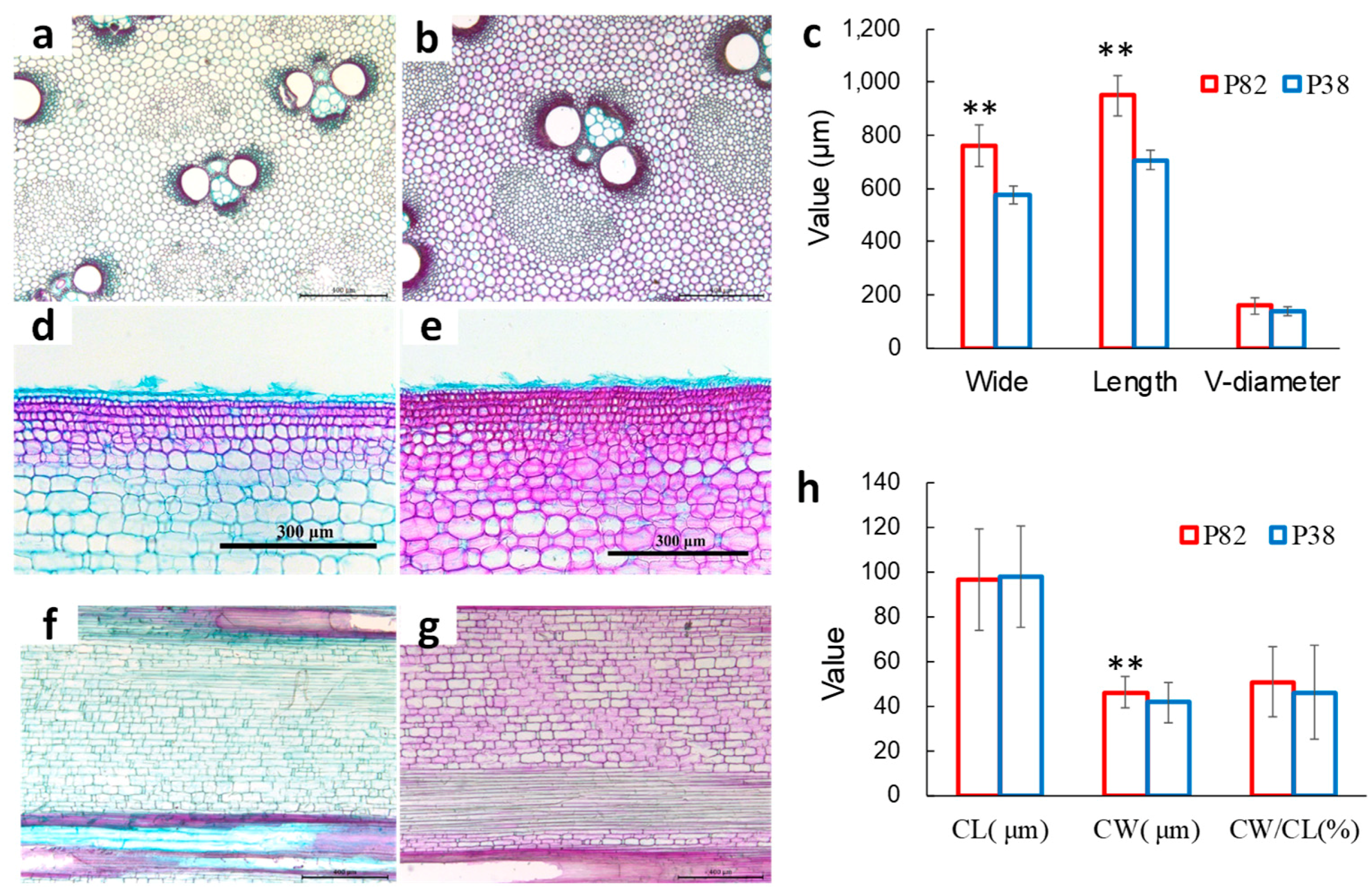
3.7. Culm Anatomical Properties of Superior Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nadgauda, R.S.; Parasharami, V.A.; Mascarenhas, A.F. Precocious Flowering and Seeding Behaviour in Tissue-cultured Bamboos. Nature 1990, 344, 335–336. [Google Scholar] [CrossRef]
- Chen, G.C.; Ma, N.X. Advances in studies on Geneties and Breeding of Bamboos. For. Res. 2005, 6, 749–754. [Google Scholar]
- Xie, N.; Chen, L.N.; Dong, Y.R.; Yang, H.Q. Mixed Mating System and Variable Mating Patterns in Tropical Woody Bamboos. BMC Plant Biol. 2019, 19, 418. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.T.; Fu, M.Y.; Xiao, X.T.; Wang, P.H.; Zhuang, J.K.; Lin, Y.H. Sowing Properties of Controlled Pollinated Seeds and the Growth of Young Seedlings of Dendrocalamus latiflorus Munro. J. Beijing For. Univ. 2004, 1, 10–13. [Google Scholar]
- Xing, X.T.; Fu, M.Y.; Xiao, X.T. Biological Characteristics of Flowering and Controlled Pollination of Dendrocalamus latiflorus Munro. J. Beijing For. Univ. 2005, 6, 103–107. [Google Scholar]
- Yuan, J.L.; Fu, M.Y.; Zhuang, J.K.; Xiao, X.T.; Wang, P.H. The Characteristics of Flowering and Pollinating of Several Sympodial Bamboos and Elementary Selection of Dendrocalamus latiflorus Seedlings. J. Bamboo Res. 2005, 3, 9–13. [Google Scholar]
- Yang, X.Y.; Fu, M.Y.; Xing, X.T. Study on Genetic Variations and Selections of Controlling Pollination Progeny at Seedling Stage of Dendrocalamus latiflorus. World Bamboo Ratt. 2008, 2, 11–13. [Google Scholar]
- Yang, X.Y.; Fu, M.Y.; Yang, X.S.; Fei, X.Q. Variation of Bamboo Shoot Nutrient Components of the Hybrids of Dendrocalamus latiflorus Munro. J. Huazhong Agric. Univ. 2010, 1, 101–106. [Google Scholar]
- Guangchu, Z.; Fuzhu, C. Studies on the crossbreeding of bamboos. Guangdong For. Sci. Technol. 1986, 3, 1–5. [Google Scholar]
- Qirong, G. Rare New varieties of moso bamboo-Phyllostachys edulis Pachyloen. World Bamboo Ratt. 2003, 1, 40. [Google Scholar]
- Zhang, G.C. The Recent Situation of Bamboo Breeding. J. Bamboo Res. 2000, 3, 13–15. [Google Scholar]
- Ning, C.Q.; Dai, Q.H. Research on cross breeding of Bambusa pervariabilis × Dendrocalamopsis grandis. Guangxi For. Sci. 1995, 24, 167–168. [Google Scholar]
- Huang, B.H.; Hua, X.Q.; Chen, B.X. Evaluation and arrangement of High quality and high-yield shoot use bamboo species. J. Bamboo Res. 1995, 14, 18–29. [Google Scholar]
- Qiao, G.R.; Li, H.Y.; Liu, M.Y.; Jiang, J.; Yin, Y.F.; Zhang, L.; Zhuo, R.Y. Callus induction and plant regeneration from anthers of Dendrocalamus latiflorus Munro. In Vitro Cell. Dev. Biol.–Plant 2013, 49, 375–382. [Google Scholar] [CrossRef]
- Bradstreet, R.B. Kjeldahl Method for Organic Nitrogen. Anal. Chem. 1954, 26, 185–187. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, P.; Chen, L.; Wu, Z.; Wu, D. Effect of Fertilization on Nutrient Components of Bamboo Shoot and Leaves Nutrient Status Shoot Emergence Stages in Dendrocalamopsis oldhami. Sci. Silvae Sin. 2004, 6, 79–84. [Google Scholar]
- Virtanen, A.; Laine, T.; Toivonen, T. Estimation of certain amino acids by the Ninhydrin method. Biol. Chem. 1940, 266, 193–204. [Google Scholar]
- Wang, Y.J.; Zhan, H.; Ding, Y.J. Variability of anatomical and chemical properties with age and height in Dendrocalamus brandisii. BioResources 2016, 11, 1202–1213. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Wan, S.B.; Dai, L.X.; Song, W.W.; Chen, J.; Shi, Y.Q. Estimating and screening of drought resistance indexes of peanut. Chin. J. Plant Ecol. 2011, 1, 100–109. [Google Scholar] [CrossRef]
- Guo, Z.H.; Ma, P.F.; Yang, G.Q.; Hu, J.Y.; Liu, Y.L.; Xia, E.H.; Zhong, M.C.; Zhao, L.; Sun, G.L.; Xu, Y.X.; et al. Genome Sequences Provide Insights into the Reticulate Origin and Unique Traits of Woody Bamboos. Mol. Plant 2019, 12, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Lin, S.; Fu, H.; Wan, Y.; Ding, Y. The Bamboo Flowering Cycle Sheds Light on Flowering Diversity. Front. Plant Sci. 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Cistué, L.; Echávarri, B.; Batlle, F.; Soriano, M.; Castillo, A.; Vallés, M.P.; Romagosa, I. Segregation distortion for agronomic traits in doubled haploid lines of barley. Plant Breed. 2005, 124, 546–550. [Google Scholar] [CrossRef]
- Meng, Y.J.; Li, J.H.; Liu, J.J.; Hu, H.X.; Li, W.E.I.; Liu, W.X. Ploidy effect and genetic architecture exploration of stalk traits using DH and its corresponding haploid populations in maize. BMC Plant Biol. 2016, 16, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cistué, L.; Soriano, M.; Castillo, A.; Vallés, M.P.; Sanz, J.M.; Echavarri, B. Production of doubled haploids in durum wheat (Triticum turgidum L.) through isolated microspore culture. Plant Cell Rep. 2006, 25, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Maheshwari, S.C. In vitro production of embryos from anthers of Datura. Nature 1964, 212, 97–98. [Google Scholar] [CrossRef]
- Te Beest, M.; Le Roux, J.J.; Richardson, D.M.; Brysting, A.K.; Suda, J.; Kubešová, M.; Pyšek, P. The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot. 2012, 109, 19–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.X.; Zhang, G.C.; Li, X.W. An Evaluation on Shoot number of Sympodial Bamboo Species and Their Hybrids. J. Bamboo Res. 2005, 4, 39–44. [Google Scholar]
- Deng, L.; Li, P.C.; Chu, C.H.; Ding, Y.L.; Wang, S.G. Symplasmic phloem unloading and post-phloem transport during bamboo internode elongation. Tree Physiol. 2020, 40, 391–412. [Google Scholar] [CrossRef] [PubMed]







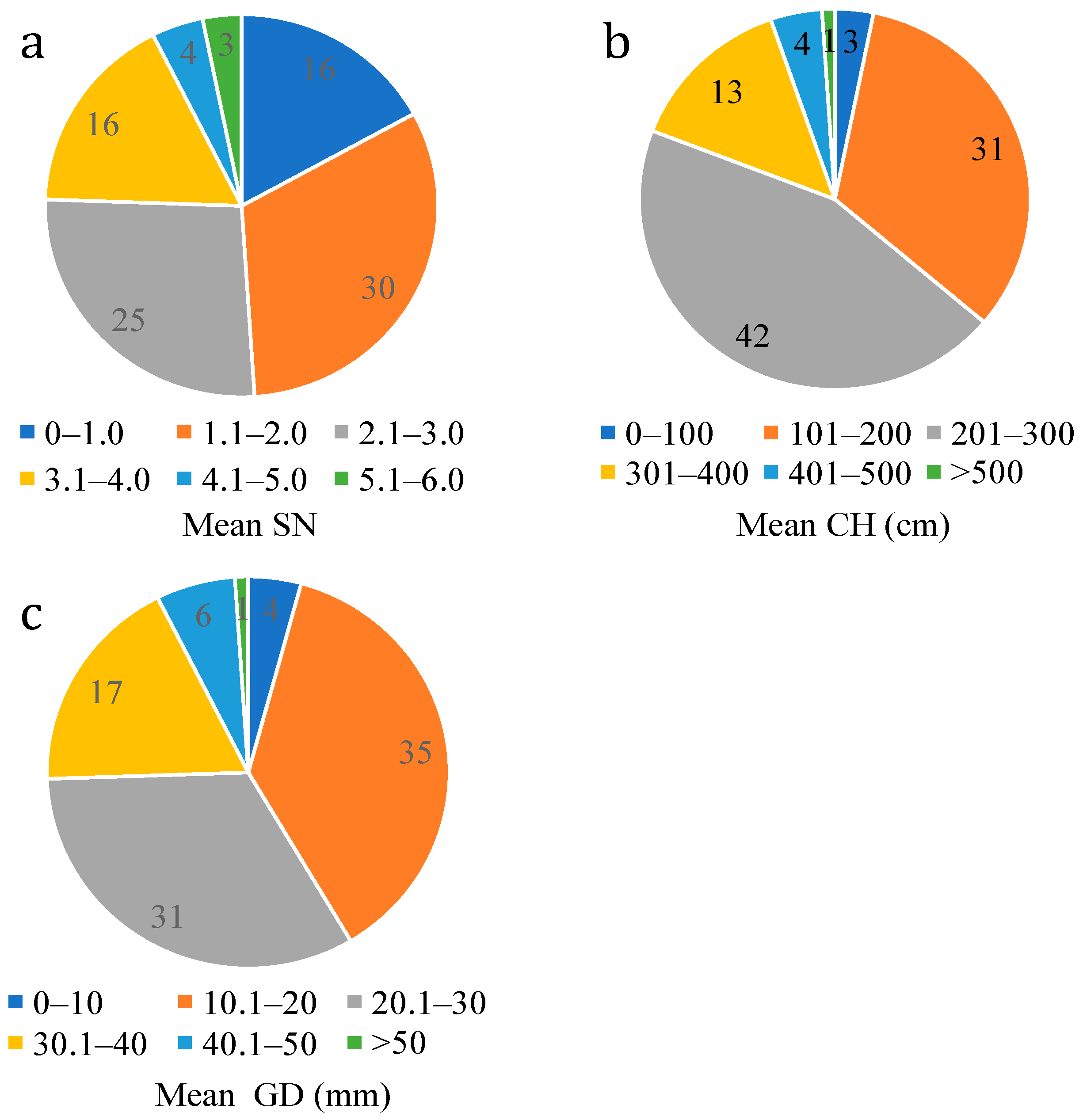{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate and Altitude Conditions | Value | Soil Conditions | Value |
|---|---|---|---|
| Altitude | 155 m | Thickness | >1 m |
| Mean annual temperature | 17.3 °C | pH | 5.32 |
| Maximum temperature | 39.5 °C | Organic matter | 37.89 g·kg−1 |
| Minimum temperature | −3.7 °C | Available nitrogen | 233.84 mg·kg−1 |
| Mean annual frost-free period | 320 d | Available phosphorus | 50.01 mg·kg−1 |
| Mean annual precipitation | 1448–2023 mm | Available potassium | 19.32 mg·kg−1 |
| Mean | Maximum | Minimum | Maximum Difference | Coefficient of Variation % | |
|---|---|---|---|---|---|
| CH of mother regenerated lines (cm) | 19.5 | 35.0 | 3.0 | 32.0 | 28.5 |
| CH of first shoot (cm) | 50.1 | 184.0 | 12.0 | 172.0 | 52.4 |
| SN per cluster | 2.9 | 12.0 | 0 | 12.0 | 45.1 |
| Plant Label | SN | CH (cm) | GD (mm) | Mean MFV | |||
|---|---|---|---|---|---|---|---|
| Mean ± SD | MFV | Mean ± SD | MFV | Mean ± SD | MFV | ||
| P82 | 3.7 ± 0.5 | 0.576 | 697.3 ± 87.2 | 1.000 | 61.9 ± 6.8 | 1.000 | 0.859 |
| P84 | 2.5 ± 1.5 | 0.364 | 420.3 ± 100.1 | 0.614 | 40.5 ± 12.8 | 0.571 | 0.516 |
| P38 | 4.7 ± 0.9 | 0.758 | 269.7 ± 16.2 | 0.416 | 29.5 ± 5.5 | 0.338 | 0.504 |
| P34 | 5.0 ± 0.0 | 0.818 | 260.0 ± 39.6 | 0.304 | 23.4 ± 5.8 | 0.322 | 0.482 |
| P03 | 3.0 ± 1.0 | 0.455 | 379.7 ± 9.9 | 0.445 | 31.2 ± 0.8 | 0.508 | 0.469 |
| P20 | 1.0 ± 0.1 | 0.091 | 459.7 ± 106.7 | 0.681 | 44.2 ± 6.3 | 0.633 | 0.468 |
| P18 | 3.3 ± 1.9 | 0.515 | 314.7 ± 18.4 | 0.458 | 31.9 ± 1.9 | 0.408 | 0.460 |
| P109 | 2.5 ± 0.5 | 0.364 | 335.3 ± 34.5 | 0.563 | 37.7 ± 4.0 | 0.439 | 0.455 |
| P37 | 3.0 ± 1.6 | 0.455 | 330.3 ± 36.3 | 0.477 | 32.9 ± 5.6 | 0.431 | 0.454 |
| P116 | 1.0 ± 0.2 | 0.091 | 450.0 ± 39.8 | 0.652 | 42.6 ± 4.6 | 0.617 | 0.453 |
| P22 | 1.0 ± 0.1 | 0.091 | 375.0 ± 144.7 | 0.762 | 48.7 ± 20.1 | 0.501 | 0.451 |
| P40 | 3.7 ± 0.5 | 0.576 | 264.7 ± 44.1 | 0.429 | 30.3 ± 6.1 | 0.330 | 0.445 |
| P45 | 2.3 ± 0.9 | 0.333 | 368.7 ± 121.2 | 0.493 | 33.8 ± 9.8 | 0.490 | 0.439 |
| P33 | 1.7 ± 0.5 | 0.212 | 378.0 ± 70.4 | 0.586 | 39.0 ± 5.8 | 0.506 | 0.435 |
| P16 | 1.0 ± 0.1 | 0.091 | 403.0 ± 73.4 | 0.640 | 41.9 ± 6.8 | 0.543 | 0.425 |
| P56 | 4.5 ± 0.5 | 0.727 | 235.3 ± 14.8 | 0.229 | 19.2 ± 3.0 | 0.283 | 0.413 |
| P14 | 1.5 ± 0.5 | 0.182 | 329.7 ± 10.2 | 0.618 | 40.8 ± 0.9 | 0.431 | 0.410 |
| P69 | 4.0 ± 0.0 | 0.636 | 225.0 ± 54.9 | 0.308 | 23.6 ± 9.6 | 0.268 | 0.404 |
| P28 | 3.0 ± 0.0 | 0.455 | 310.3 ± 121.6 | 0.352 | 26.0 ± 6.5 | 0.400 | 0.402 |
| P94 | 2.5 ± 0.5 | 0.364 | 275.3 ± 5.2 | 0.495 | 33.9 ± 1.4 | 0.345 | 0.401 |
| Plant Label | Ash (g·100 g−1) | Soluble Sugar (%) | Free Amino Acid (g·kg−1 FW) | Protein (g·kg−1 FW) | Hydrolyzed Amino Acids (g·kg−1 FW) | Tannin (g·kg−1 FW) | Oxalic Acid (g·kg−1 FW) |
|---|---|---|---|---|---|---|---|
| P38 | 0.8 ± 0.08 a | 1.5 ± 0.3 a | 3.2 ± 0.2 a | 16.4 ± 2.8 a | 9.4 ± 2.1 a | 2.5 ± 0.2 a | 1.8 ± 0.1 a |
| P84 | 0.6 ± 0.07 b | 1.0 ± 0.1 b | 3.6 ± 0.3 a | 17.0 ± 2.3 a | 10.4 ± 2.3 a | 2.2 ± 0.4 ab | 2.1 ± 0.3 a |
| P82 | 0.6 ± 0.05 b | 1.4 ± 0.1 ab | 3.5 ± 0.5 a | 14.8 ± 1.4 a | 9.1 ± 2.3 a | 2.0 ± 0.4 ab | 1.8 ± 0.2 a |
| WT | 0.5 ± 0.03 b | 1.1 ± 0.1 b | 1.8 ± 0.1 b | 9.6 ± 1.5 b | 6.4 ± 0.6 b | 1.6 ± 0.2 b | 2.1 ± 0.3 a |
| Local | 0.6 ± 0.04 b | 1.2 ± 0.3 ab | 3.5 ± 0.5 a | 17.2 ± 3.4 a | 11.9 ± 1.7 a | 2.4 ± 0.5 ab | 1.0 ± 0.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Wang, Y.; Qiao, G.; Fan, H.; Jin, K.; Huang, B.; Qiu, W.; Zou, Y.; Xie, J.; Zhuo, R. Superior Line from Anther Culture of Dendrocalamus latiflorus Selected after Field Trial. Horticulturae 2021, 7, 98. https://doi.org/10.3390/horticulturae7050098
Zhang W, Wang Y, Qiao G, Fan H, Jin K, Huang B, Qiu W, Zou Y, Xie J, Zhuo R. Superior Line from Anther Culture of Dendrocalamus latiflorus Selected after Field Trial. Horticulturae. 2021; 7(5):98. https://doi.org/10.3390/horticulturae7050098
Chicago/Turabian StyleZhang, Wei, Yujun Wang, Guirong Qiao, Huijin Fan, Kangming Jin, Biyun Huang, Wenmin Qiu, Yueguo Zou, Jinzhong Xie, and Renying Zhuo. 2021. "Superior Line from Anther Culture of Dendrocalamus latiflorus Selected after Field Trial" Horticulturae 7, no. 5: 98. https://doi.org/10.3390/horticulturae7050098
APA StyleZhang, W., Wang, Y., Qiao, G., Fan, H., Jin, K., Huang, B., Qiu, W., Zou, Y., Xie, J., & Zhuo, R. (2021). Superior Line from Anther Culture of Dendrocalamus latiflorus Selected after Field Trial. Horticulturae, 7(5), 98. https://doi.org/10.3390/horticulturae7050098