
Characterization of Japanese Apricot (Prunus mume) Floral Bud Development Using a Modified BBCH Scale and Analysis of the Relationship between BBCH Stages and Floral Primordium Development and the Dormancy Phase Transition


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Evaluation of Growth/Dormancy Status
2.3. Estimation of the CR for Endodormancy Release and the Heat Requirement (HR) for Blooming
2.4. Analyses of Bud Phenology and Internal Floral Primordium Development
3. Results
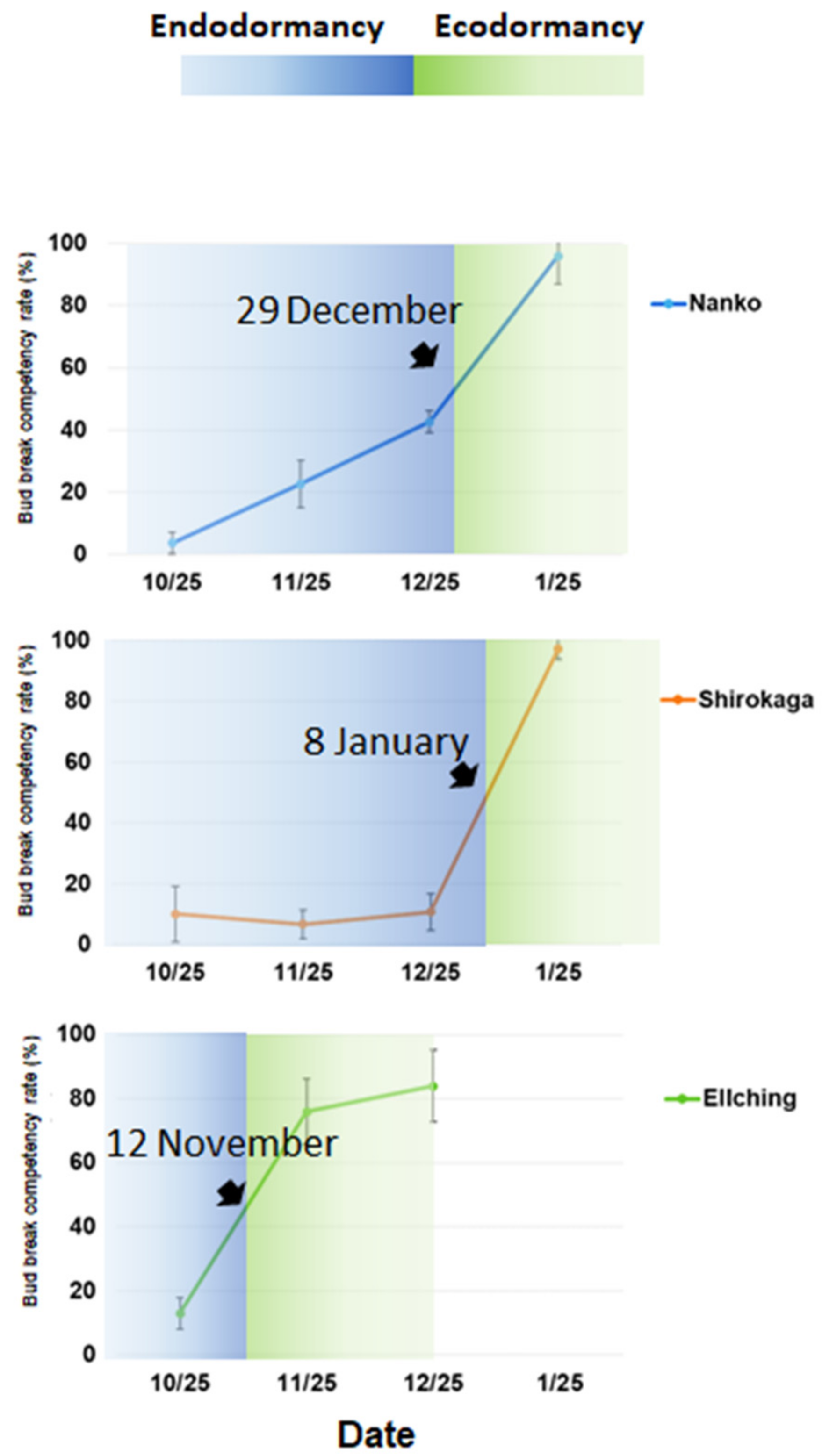
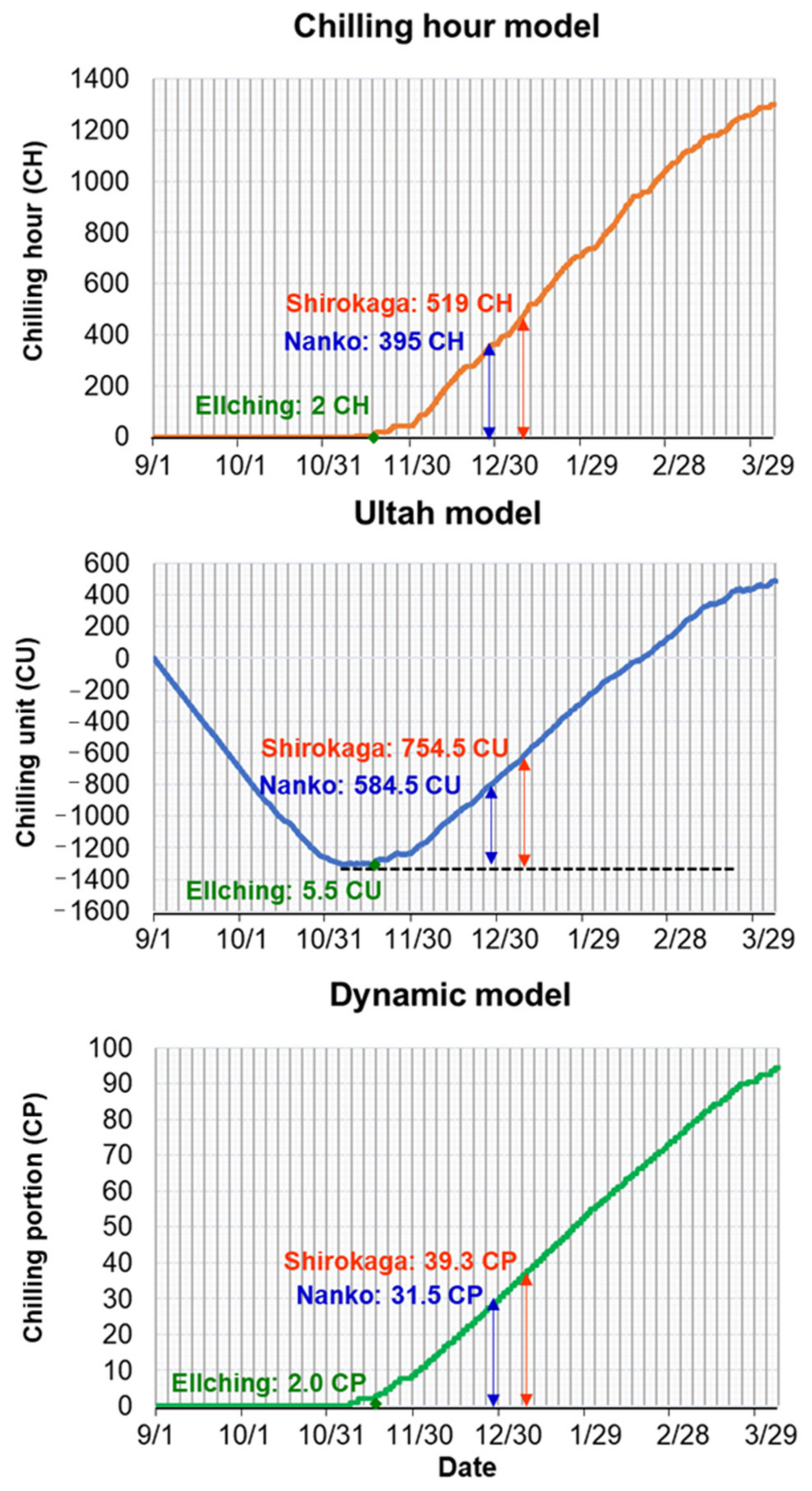
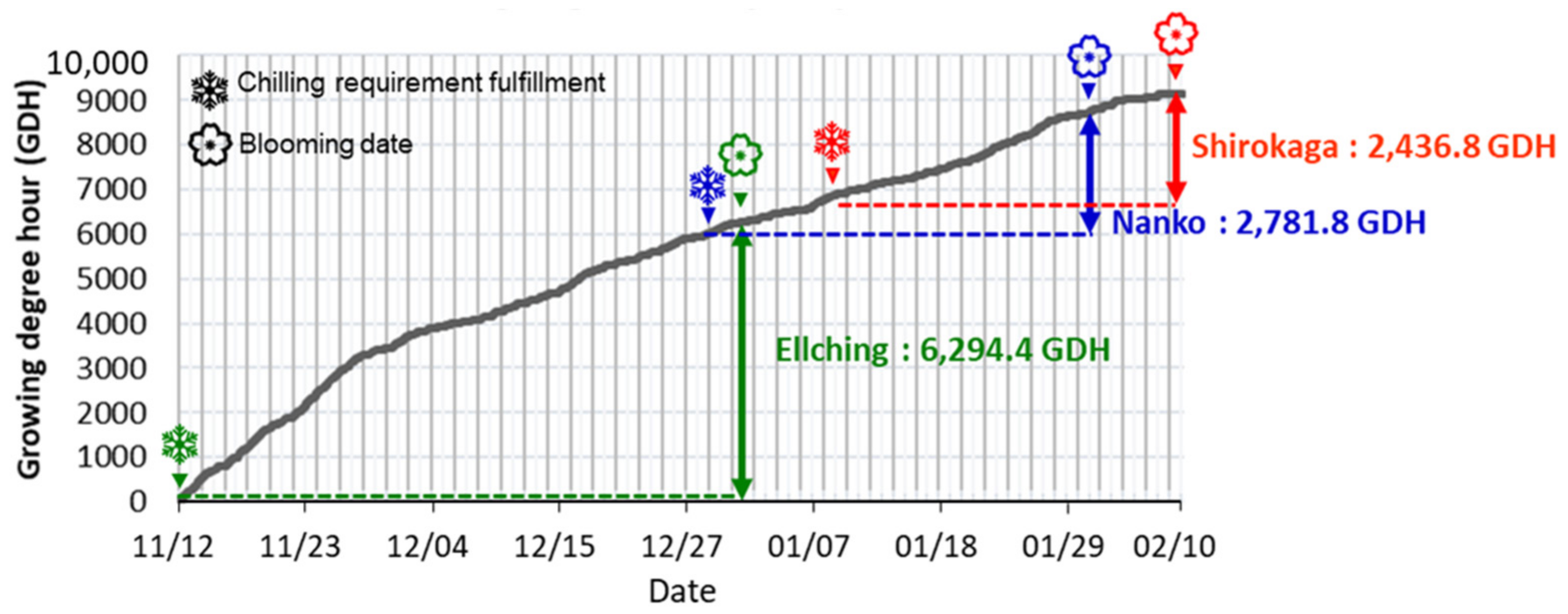
3.1. Contrasting the CR and HR of Three Cultivars
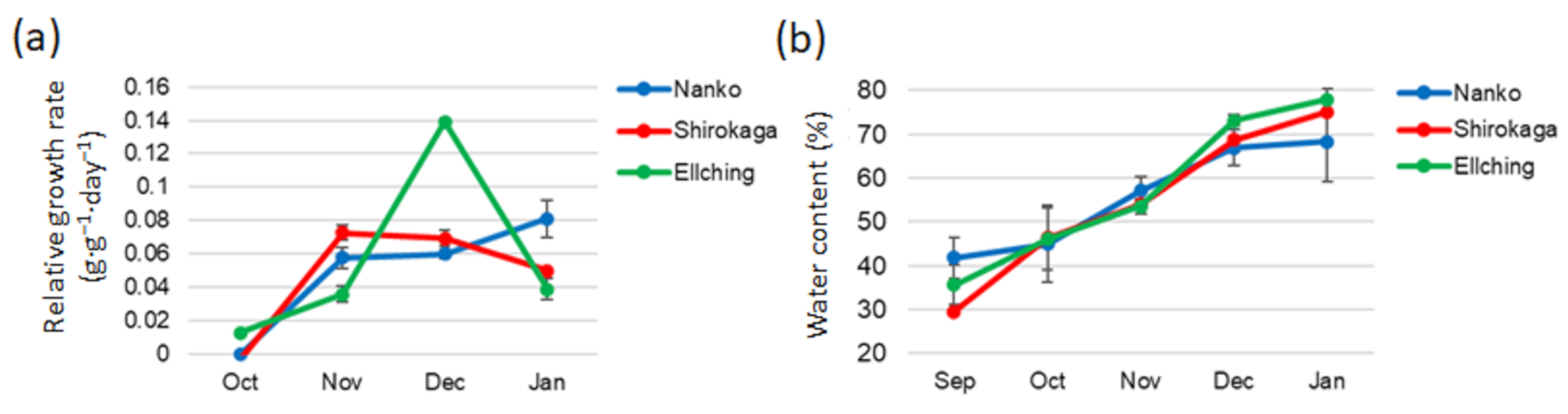
3.2. Seasonal Changes in the RGR and Water Content of the Low-Chill and High-Chill Cultivars
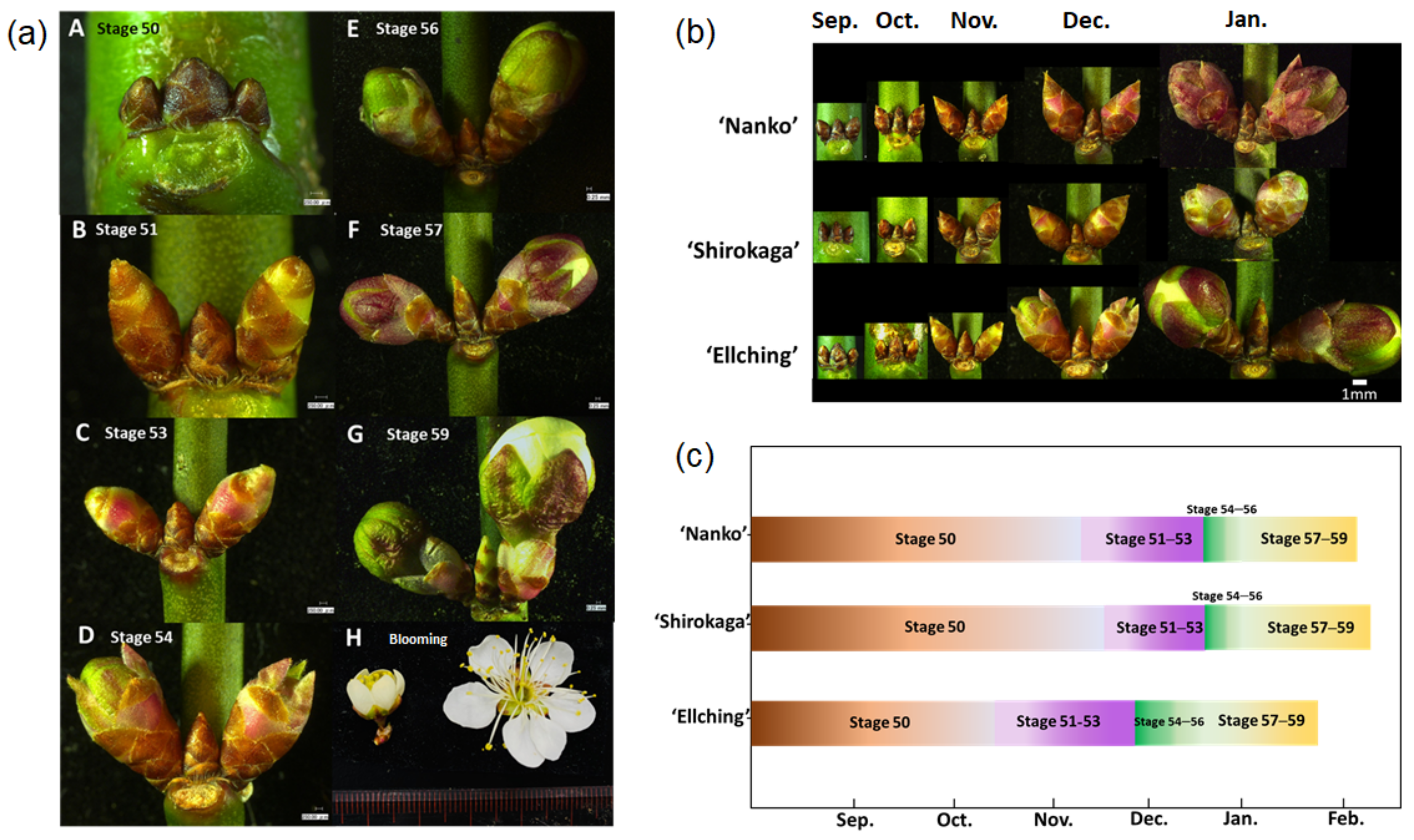
3.3. Establishment of a Modified BBCH Scale for Japanese Apricot Floral Buds
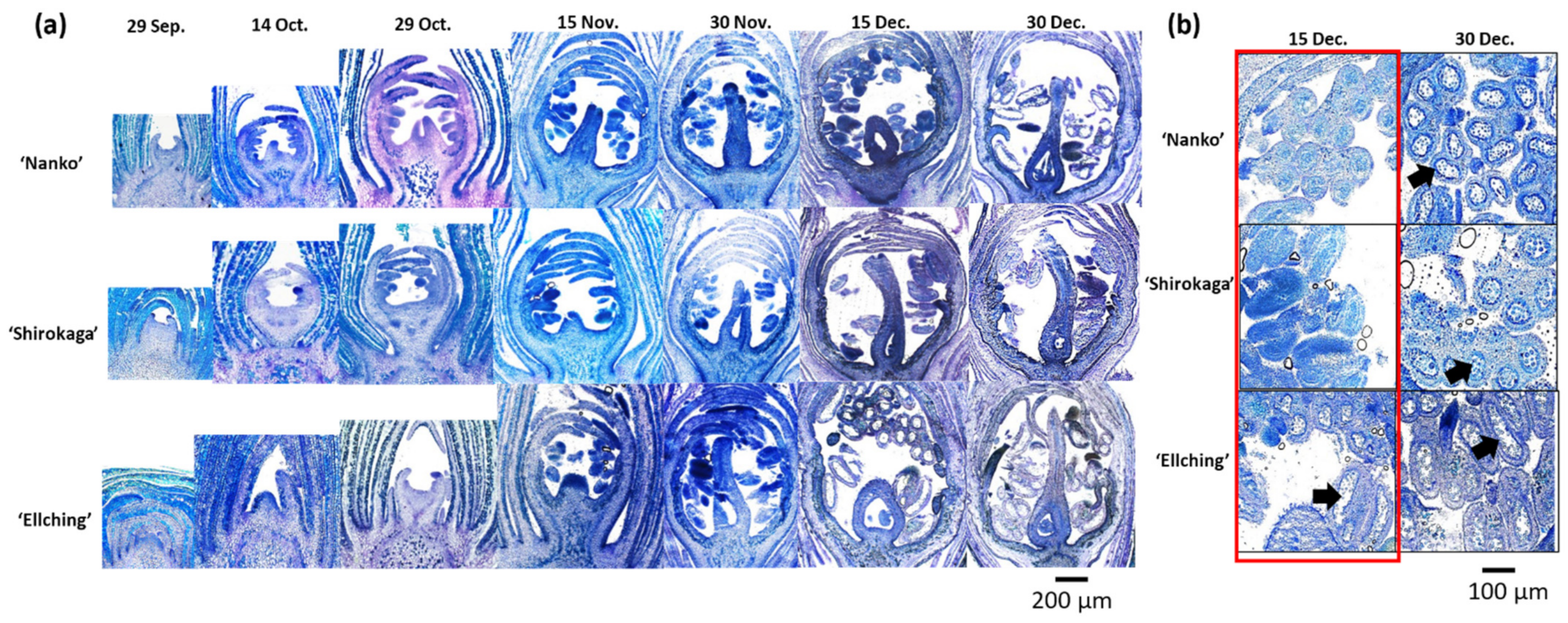
3.4. Comparison of Seasonal Floral Primordium Development among Cultivars with Contrasting CRs
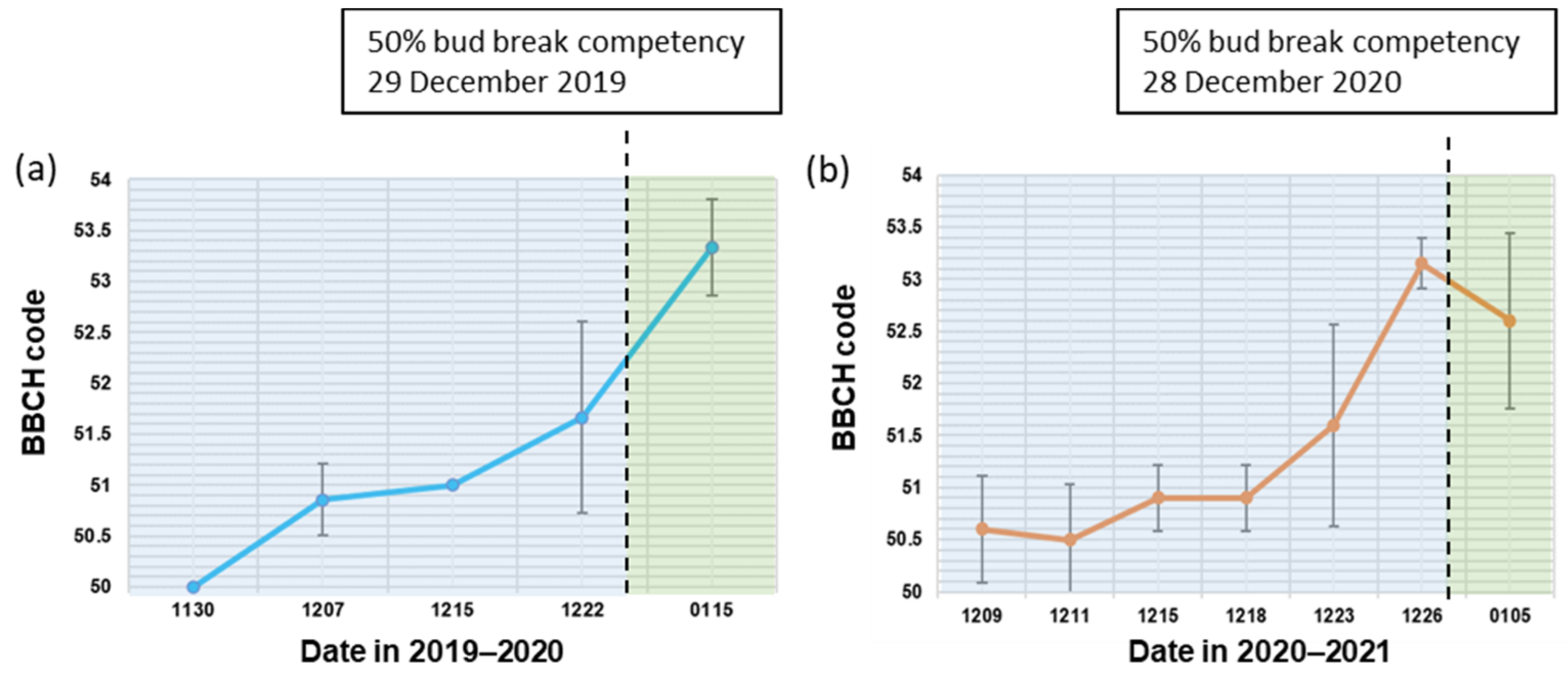
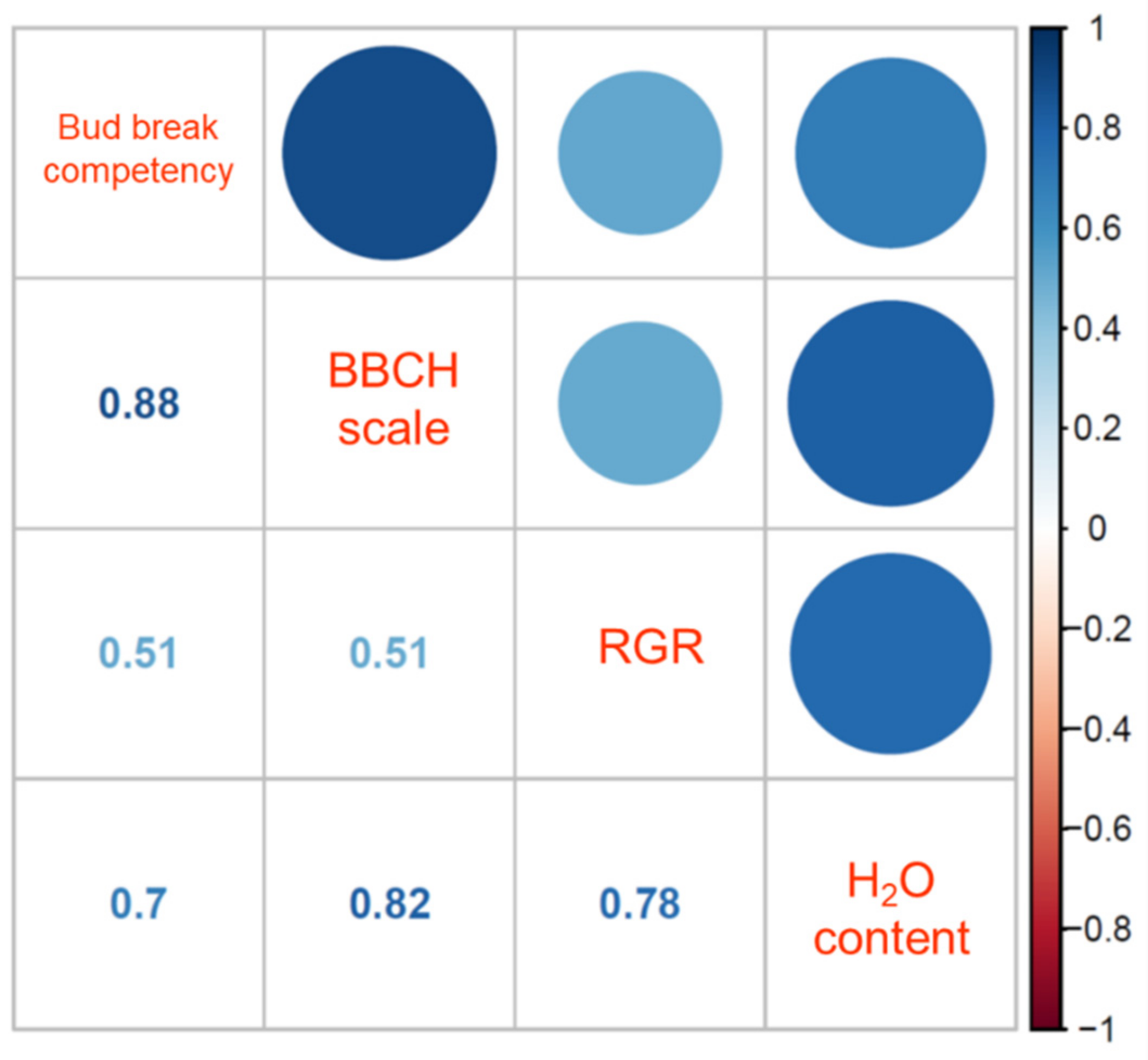
3.5. Correlation between BBCH Stages and the Floral Bud Physiological Dormancy Status
4. Discussion
4.1. Comparison of the Japanese Apricot Floral Bud Dormancy Progression of Cultivars with Different Blooming Dates
4.2. Calculated CRs and HRs for Japanese Apricot Floral Buds
4.3. Relationship between the Dormancy Phase Transition and BBCH Stages in Japanese Apricot
4.4. Connection between Pollen Maturation and the Dormancy Status in Japanese Apricot
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Hertogh, A. Principles for forcing tulips, hyacinths, daffodils, easter lilies and dutch irises. Sci. Hortic. 1974, 2, 313–355. [Google Scholar] [CrossRef]
- Fadón, E.; Herrero, M.; Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- Hsiang, T.-F.; Chen, W.; Yamane, H. The MADS-box gene family involved in the regulatory mechanisms of dormancy and flowering in Rosaceae fruit trees. Annu. Plant Rev. Online 2021, in press. [Google Scholar]
- Julian, C.; Rodrigo, J.; Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Ann. Bot. 2011, 108, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower development in sweet cherry framed in the BBCH scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Herrero, M.; Rodrigo, J. Anther and pollen development in sweet cherry (Prunus avium L.) in relation to winter dormancy. Protoplasma 2019, 256, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Luedeling, E.; Behrend, D.; Van De Vliet, S.; Kunz, A.; Fadón, E. Mild water stress makes apple buds more likely to flower and more responsive to artificial forcing—Impacts of an unusually warm and dry summer in germany. Agronomy 2020, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- Penso, G.A.; Citadin, I.; Scariotto, S.; Magalhães dos Santos, C.E.; Junior, A.W.; Bruckner, C.H.; Rodrigo, J. Development of peach flower buds under low winter chilling conditions. Agronomy 2020, 10, 428. [Google Scholar] [CrossRef] [Green Version]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants; Federal Biological Research Centre for Agriculture and Forestry: Kleinmachnow, Germany, 2001; p. 158. [Google Scholar]
- Pérez-Pastor, A.; Ruiz-Sánchez, M.; Domingo, R.; Torrecillas, A. Growth and phenological stages of Búlida apricot trees in south-east Spain. Agronomie 2004, 24, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Sakar, E.H.; El Yamani, M.; Boussakouran, A.; Rharrabti, Y. Codification and description of almond (Prunus dulcis) vegetative and reproductive phenology according to the extended BBCH scale. Sci. Hortic. 2019, 247, 224–234. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Szalay, L.; Froemel-Hajnal, V.; Bakos, J.; Ladányi, M. Changes of the microsporogenesis process and blooming time of three apricot genotypes (Prunus armeniaca L.) in central hungary based on long-term observation (1994–2018). Sci. Hortic. 2019, 246, 279–288. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Herrero, M.; Rodrigo, J. Male meiosis in sweet cherry is constrained by the chilling and forcing phases of dormancy. Tree Physiol. 2020, 41, 619–630. [Google Scholar] [CrossRef]
- McPherson, H.G.; Snelgar, W.P.; Manson, P.J.; Snowball, A.M. Bud respiration and dormancy of kiwifruit (Actinidia deliciosa). Ann. Bot. 1997, 80, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and heat requirements of temperate stone fruit trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for redhaven and elberta peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Anderson, J.L.; Ashcroft, G.L. Pheno-climatography of spring peach bud development. HortScience 1975, 10, 236–237. [Google Scholar]
- Anderson, J.L.; Richardson, E.A. The Utah chill unit/flower bud phenology models for deciduous fruit: Their implication for production in subtropical areas. Acta Hortic. 1987, 199, 45–50. [Google Scholar] [CrossRef]
- Götz, K.P.; Chmielewski, F.M.; Homann, T.; Huschek, G.; Matzneller, P.; Rawel, H.M. Seasonal changes of physiological parameters in sweet cherry (Prunus avium L.) buds. Sci. Hortic. 2014, 172, 183–190. [Google Scholar] [CrossRef]
- Saito, T.; Tuan, P.A.; Katsumi-Horigane, A.; Bai, S.; Ito, A.; Sekiyama, Y.; Ono, H.; Moriguchi, T. Development of flower buds in the Japanese pear (Pyrus pyrifolia) from late autumn to early spring. Tree Physiol. 2015, 35, 653–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, R.R.; Akemi, K.H.; Yoshida, M.; Sekozawa, Y.; Sugaya, S.; Gemma, H. Floral primordia necrosis” Incidence in mixed buds of Japanese pear (Pyrus pyrifolia (Burm.) Nakai var. Culta) ‘Housui’ grown under mild winter conditions and the possible relation with water dynamics. J. Jpn. Soc. Hortic. Sci. 2010, 79, 246–257. [Google Scholar]
- Yamane, H.; Kashiwa, Y.; Kakehi, E.; Yonemori, K.; Mori, H.; Hayashi, K.; Iwamoto, K.; Tao, R.; Kataoka, I. Differential expression of dehydrin in flower buds of two Japanese apricot cultivars requiring different chilling requirements for bud break. Tree Physiol. 2006, 26, 1559–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, Y.; Yamane, H.; Yukimori, A.; Shimo, H.; Numaguchi, K.; Tao, R. Blooming date predictions based on Japanese apricot ‘Nanko’ flower bud responses to temperatures during dormancy. HortScience 2017, 52, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Prudencio, A.S.; Martínez-Gómez, P.; Dicenta, F. Evaluation of breaking dormancy, flowering and productivity of extra-late and ultra-late flowering almond cultivars during cold and warm seasons in south-east of Spain. Sci. Hortic. 2018, 235, 39–46. [Google Scholar] [CrossRef]
- Okie, W.R.; Blackburn, B. Increasing chilling reduces heat requirement for floral budbreak in peach. HortScience 2011, 46, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Heide, O.M. Interaction of photoperiod and temperature in the control of growth and dormancy of Prunus species. Sci. Hortic. 2008, 115, 309–314. [Google Scholar] [CrossRef]
- Renaut, J.; Hausman, J.F.; Bassett, C.; Artlip, T.; Cauchie, H.M.; Witters, E.; Wisniewski, M. Quantitative proteomic analysis of short photoperiod and low-temperature responses in bark tissues of peach (Prunus persica L. Batsch). Tree Genet. Genomes 2008, 4, 589–600. [Google Scholar] [CrossRef]
- Wisniewski, M.E.; Bassett, C.L.; Renaut, J.; Farrell, R.; Tworkoski, T.; Artlip, T.S. Differential regulation of two dehydrin genes from peach (Prunus persica) by photoperiod, low temperature and water deficit. Tree Physiol. 2006, 26, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Hernandez, M.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Deciphering the role of CBF/DREB transcription factors and dehydrins in maintaining the quality of table grapes cv. Autumn Royal treated with high CO2 levels and stored at 0 °C. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Viti, R.; Bartolini, S.; Andreini, L. Apricot flower bud dormancy: Main morphological, anatomical and physiological features related to winter climate influence. Adv. Hortic. Sci. 2013, 27, 5–17. [Google Scholar]
- Julian, C.; Herrero, M.; Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environ. Exp. Bot. 2014, 100, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Yamane, H.; Ooka, T.; Jotatsu, H.; Kitamura, Y.; Akagi, T.; Tao, R. Functional and expressional analyses of PmDAM genes associated with endodormancy in Japanese apricot. Plant Physiol. 2011, 157, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Considine, M.J.; Considine, J.A. On the language and physiology of dormancy and quiescence in plants. J. Exp. Bot. 2016, 67, 3189–3203. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | BD2020 1 | Origin |
|---|---|---|
| Ellching | 6 | Taiwan |
| Hayazakinanko | 7 | Japan |
| 1C1-10 | 24 | Japan |
| Kairyo-uchida | 28 | Japan |
| Ryukyokoume | 32 | Japan |
| 1KO-26 | 33 | Japan |
| Benisashi | 34 | Japan |
| Nanko | 35 | Japan |
| Kotsubnu-nanko | 35 | Japan |
| Koshinoume | 35 | Japan |
| Kagajizo | 37 | Japan |
| Kensaki | 38 | Japan |
| Shirokaga | 41 | Japan |
| Hachiro | 47 | Japan |
| Rinshu | 49 | Japan |
| Koshukoume | 49 | Japan |
| Aojiku | 49 | Japan |
| Oshuku | 49 | Japan |
| Gyokuei | 49 | Japan |
| Principal Growth Stage 5: Flower Emergence | |
|---|---|
| 50 | Floral bud closed, dark brown scales visible |
| 51 | Floral bud swelling, bud still closed, light brown scales visible |
| 53 | Scales further separated, light green bud sections, and light purple or pink scales visible |
| 54 | Internal floral organ enclosed by light green sepals, round green sepals visible |
| 56 | Flower pedicel elongating, green sepals still closed showing a round shape |
| 57 | Sepals open: petal tips visible, flower with white petals (still closed) |
| 59 | Flowers with petals forming a hollow ball |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiang, T.-F.; Lin, Y.-J.; Yamane, H.; Tao, R. Characterization of Japanese Apricot (Prunus mume) Floral Bud Development Using a Modified BBCH Scale and Analysis of the Relationship between BBCH Stages and Floral Primordium Development and the Dormancy Phase Transition. Horticulturae 2021, 7, 142. https://doi.org/10.3390/horticulturae7060142
Hsiang T-F, Lin Y-J, Yamane H, Tao R. Characterization of Japanese Apricot (Prunus mume) Floral Bud Development Using a Modified BBCH Scale and Analysis of the Relationship between BBCH Stages and Floral Primordium Development and the Dormancy Phase Transition. Horticulturae. 2021; 7(6):142. https://doi.org/10.3390/horticulturae7060142
Chicago/Turabian StyleHsiang, Tzu-Fan, Yuan-Jui Lin, Hisayo Yamane, and Ryutaro Tao. 2021. "Characterization of Japanese Apricot (Prunus mume) Floral Bud Development Using a Modified BBCH Scale and Analysis of the Relationship between BBCH Stages and Floral Primordium Development and the Dormancy Phase Transition" Horticulturae 7, no. 6: 142. https://doi.org/10.3390/horticulturae7060142
APA StyleHsiang, T. -F., Lin, Y. -J., Yamane, H., & Tao, R. (2021). Characterization of Japanese Apricot (Prunus mume) Floral Bud Development Using a Modified BBCH Scale and Analysis of the Relationship between BBCH Stages and Floral Primordium Development and the Dormancy Phase Transition. Horticulturae, 7(6), 142. https://doi.org/10.3390/horticulturae7060142