
An Overview of the Practices and Management Methods for Enhancing Seed Production in Conifer Plantations for Commercial Use


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Factors Affecting Seed Formation and Development
3.1.1. Pollen
3.1.2. Temperature and Light
3.1.3. Water and Nutrient Fertilization
3.1.4. Molecular Mechanisms
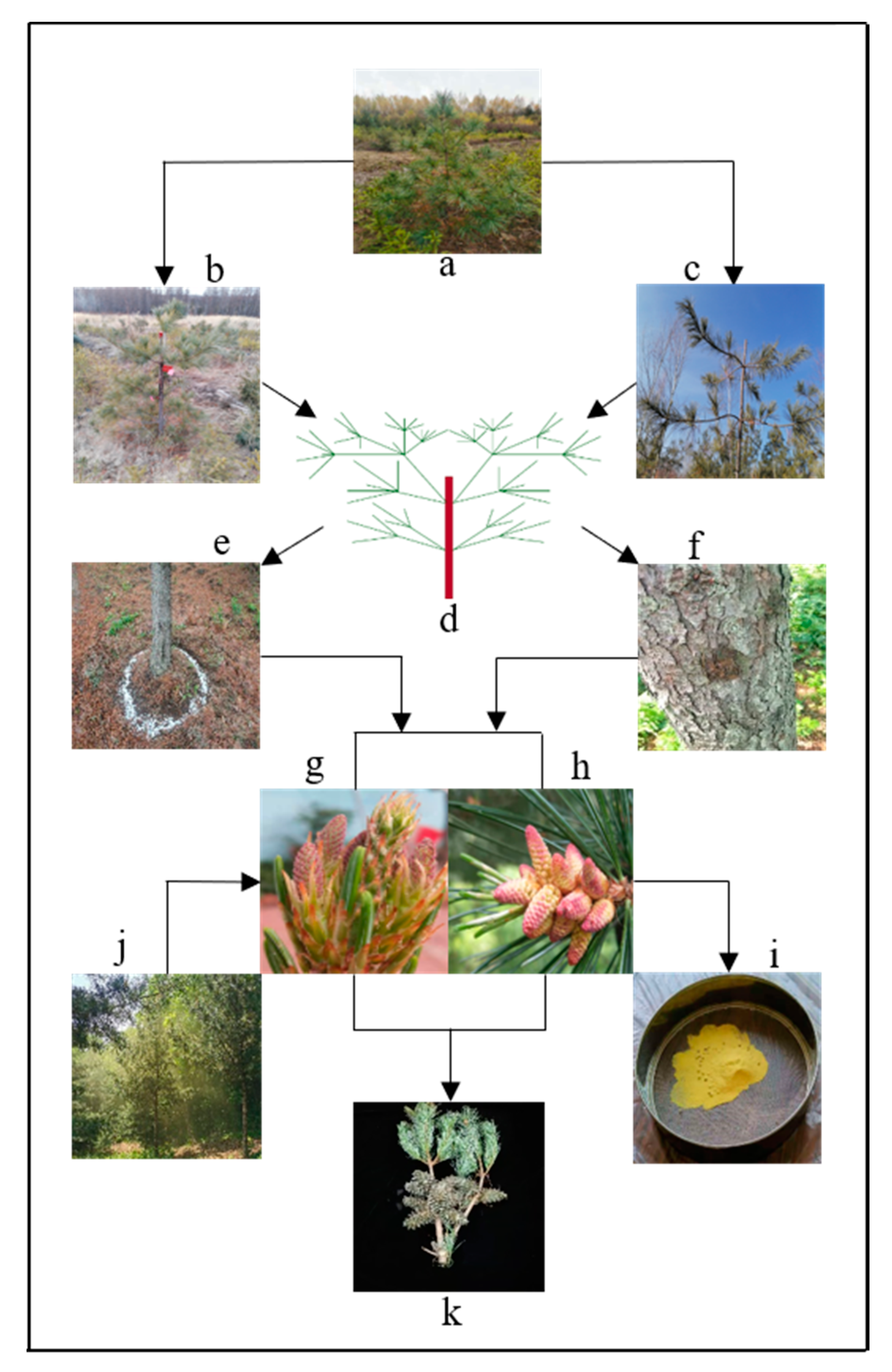
3.2. Technical Measures of Breeding and Management
3.2.1. Supplementary Pollination
3.2.2. Topping Off
3.2.3. Thinning and Pruning
3.2.4. Girdling and Cutting Roots
3.2.5. Water and Fertilization Management
3.2.6. Phytohormone Treatments
4. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Miguel, M.; Bartholomé, J.; Ehrenmann, F.; Murat, F.; Moriguchi, Y.; Uchiyama, K.; Ueno, S.; Tsumura, Y.; Lagraulet, H.; de Maria, N.; et al. Evidence of intense chromosomal shuffling during conifer evolution. Genome Biol. Evol. 2015, 7, 2799–2809. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, X.T.; Wei, J.T.; Li, Y.; Tigabu, M.; Zhao, X.Y. Genetic improvement of Pinus koraiensis in China: Current situation and future prospects. Forests 2020, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Li, H.L. Present distribution and habitats of the conifers and taxads. Evolution 1953, 7, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Christenhusz, M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.S.; Li, X.F. Reassessment of endangered conifer species in East Asia. Shanghai Acad. Agric. Sci. 2014, 30, 42–46. [Google Scholar]
- Cooper, R.J. World markets for coniferous forest products: Recent trends and future prospects. Acta Hortic. 2003, 349–353. [Google Scholar] [CrossRef]
- Dong, L.M.; Xie, Y.H.; Sun, X.M. Full-diallel-based analysis of genetic parameters for growth traits in Japanese larch (Larix kaempferi). New For. 2020, 51, 261–271. [Google Scholar] [CrossRef]
- Jokipii-Lukkari, S.; Sundell, D.; Nilsson, O.; Hvidsten, T.; Street, N.; Tuominen, H. NorWood: A gene expression resource for evo-devo studies of conifer wood development. New Phytol. 2017, 216, 482–494. [Google Scholar] [CrossRef] [Green Version]
- Nergiz, C.; Doenmez, I. Chemical composition and nutritive value of Pinus pinea L. seeds. Food Chem. 2004, 86, 365–368. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.T.; Wei, J.T.; Li, Y.; Tigabu, M.; Zhao, X.Y. Pinus koraiensis development and transferability of EST-SSR markers for from cold-stressed transcriptome through illumina sequencing. Genes 2020, 11, 500. [Google Scholar] [CrossRef]
- Missanjo, E.; Matsumura, J.; Rajora, O.P.; Martin, T.A. Genetic improvement of wood properties in Pinus kesiya royle ex gordon for sawn timber production in Malawi. Forests 2016, 7, 253. [Google Scholar] [CrossRef] [Green Version]
- Chaloupková, K.; Stejskal, J.; El-Kassaby, Y.A.; Frampton, J. Current advances in seed orchard layouts: Two case studies in conifers. Forests 2019, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Sweet, G.B. Seed orchards in development. Tree Physiol. 1995, 15, 527–530. [Google Scholar] [CrossRef]
- Liang, D.Y.; Wang, B.Y.; Song, S.L.; Wang, J.Y.; Wang, L.F.; Wang, Q.; Ren, B.; Zhao, X.Y. Analysis of genetic effects on a complete diallel cross test of Pinus koraiensis. Euphytica 2019, 215, 92. [Google Scholar] [CrossRef]
- Bilir, N.; Prescher, F.; Ayan, S.; Lindgren, D. Growth characters and number of strobili in clonal seed orchards of Pinus sylvestris. Euphytica 2006, 152, 1–9. [Google Scholar] [CrossRef]
- Lewandowski, A.; Kowalczyk, J.; Litkowiec, M.; Rzońca, M. Selection of elite plus trees of Scots pine and European larch for the establishment of 1.5 generation seed orchards. Sylwan 2017, 161, 917–926. [Google Scholar]
- Grattapaglia, D.; Diener, P.S.D.A.; Santos, G.A.D. Performance of microsatellites for parentage assignment following mass controlled pollination in a clonal seed orchard of loblolly pine (Pinus taeda L.). Tree Genet. Genomes 2014, 10, 1631–1643. [Google Scholar] [CrossRef]
- Codesido, V.; Fernández-López, J. Juvenile radiata pine clonal seed orchard management in Galicia (NW Spain). Eur. J. For. Res. 2014, 133, 177–190. [Google Scholar] [CrossRef]
- Eriksson, U.; Jansson, G.; Almqvist, C. Seed and pollen production after stem injections of gibberellin A 4/7 in field-grown seed orchards of Pinus sylvestris. Can. J. For. Res. 1998, 28, 340–346. [Google Scholar] [CrossRef]
- Huber, M.; Maschewsky-Schneider, U. Studies on the pollination characteristics and pollination level of Chinese fir seed orchard. Silvae. Genet. 2004, 53, 7–11. [Google Scholar]
- Owens, J.N.; Takaso, T.; Runions, C.J. Pollination in conifers. Trends Plant Sci. 1998, 3, 479–485. [Google Scholar] [CrossRef]
- Williams, C.G. Long-distance pine pollen still germinates after meso-scale dispersal. Am. J. Bot. 2010, 97, 846–855. [Google Scholar] [CrossRef]
- Chybicki, I.; Oleksa, A. Seed and pollen gene dispersal in Taxus baccata, a dioecious conifer in the face of strong population fragmentation. Ann. Bot. 2018, 122, 409–421. [Google Scholar] [CrossRef]
- Anderson, E.; Owens, J. Microsporogenesis, pollination, pollen germination and male gametophyte development in Taxus brevifolia. Ann. Bot. 2000, 86, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Tretyakova, I.N.; Noskova, N.E. Scotch Pine pollen under conditions of environmental stress. Russ. J. Ecol. 2004, 35, 20–26. [Google Scholar] [CrossRef]
- Batos, B.; Miljković, D. The vitality of the Serbian spruce (Picea omorika) pollen during the long-term cryopreservation. Grana 2019, 58, 433–446. [Google Scholar] [CrossRef]
- Gottardini, E.; Cristofori, A.; Cristofolini, F.; Maccherini, S.; Ferretti, M. Ambient levels of nitrogen dioxide (NO2) may reduce pollen viability in Austrian pine (Pinus nigra Arnold) trees—Correlative evidence from a field study. Sci. Total Environ. 2008, 402, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Almqvist, C.; Bergsten, U.; Bondesson, L.; Eriksson, U. Predicting germination capacity of Pinus sylvestris and Picea abies seeds using temperature data from weather stations. Can. J. For. Res. 1998, 28, 1530–1535. [Google Scholar] [CrossRef]
- Chanyenga, T.F.; Geldenhuys, C.J.; Sileshi, G.W. Germination response and viability of an endangered tropical conifer Widdringtonia whytei seeds to temperature and light. S. Afr. J. Bot. 2012, 81, 25–28. [Google Scholar] [CrossRef] [Green Version]
- Henderson, I.R.; Shindo, C.; Dean, C. The need for winter in the switch to Flowering. Annu. Rev. Genet. 2003, 37, 371–392. [Google Scholar] [CrossRef]
- Johnsen, Ø.; Dæhlen, O.G.; Østreng, G.; Skrøppa, T. Daylength and temperature during seed production interactively affect adaptive performance of Picea abies progenies. New Phytol. 2005, 168, 589–596. [Google Scholar] [CrossRef]
- Rossi, S.; Isabel, N. The timing of bud break in warming conditions: Variation among seven sympatric conifer species from Eastern Canada. Int. J. Biometeorol. 2017, 61, 1983–1991. [Google Scholar] [CrossRef]
- Tikhonova, I.V. Changes in the sex structure of pine populations related to temperature anomalies. Russ. J. Ecol. 2007, 38, 306–310. [Google Scholar] [CrossRef]
- Guo, C.C.; Shen, Y.B.; Shi, F.H. Effect of temperature, light, and storage time on the seed germination of Pinus bungeana Zucc. ex Endl.: The role of seed-covering layers and abscisic acid changes. Forests 2020, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Josef, U.; Ingwers, M.W.; Anne, M.G.M.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Han, X.Y.; Turgeon, R.; Schulz, A.; Liesche, J.; Epron, D. Environmental conditions, not sugar export efficiency, Limit the length of conifer leaves. Tree Physiol. 2018, 39, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Rozendaal, D.M.A.; Hurtado, V.H.; Poorter, L. Plasticity in leaf traits of 38 tropical tree species in response to light; Relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Salgado-Luarte, C.; Gianoli, E. Herbivory may modify functional responses to shade in seedlings of a light-demanding tree species. Funct. Ecol. 2011, 25, 492–499. [Google Scholar] [CrossRef]
- Sachin, R.S.; Nicolas, D.; Rosario, G.G.M. Transcriptome analysis of shade avoidance and shade tolerance in conifers. Planta 2019, 250, 299–318. [Google Scholar]
- Calfapietra, C.; Tulva, I.; Eensalu, E.; Perez, M.; Angelis, P.D.; Scarascia-Mugnozza, G.; Kull, O. Canopy profiles of photosynthetic parameters under elevated CO2 and N fertilization in a poplar plantation. Environ. Pollut. 2005, 137, 525–535. [Google Scholar] [CrossRef]
- Almqvist, C.; Jansson, G. Effects of pruning and stand density on cone and pollen production in an experimental Pinus sylvestris seed orchard. Silva. Fenn. 2015, 49, 1243. [Google Scholar] [CrossRef] [Green Version]
- Vanderschaaf, C.L. Estimating understory vegetation response to multi-nutrient fertilization in Douglas-fir and ponderosa pine stands. J. For. Res. 2008, 13, 43–51. [Google Scholar] [CrossRef]
- Drake, J.E.; Stoy, P.C.; Jackson, R.B.; DeLucia, E.H. Fine-root respiration in a loblolly pine (Pinus taeda L.) forest exposed to elevated CO2 and N fertilization. Plant Cell Environ. 2008, 31, 1663–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, K.; Nagakura, J.; Kaneko, S. Biomass and morphology of fine roots of sugi (Cryptomeria japonica) after 3 years of nitrogen fertilization. Front. Plant Sci. 2013, 4, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbertin, M.; Eilmann, B.; Bleuler, P.; Giuggiola, A.; Pannatier, E.G.; Landolt, W.; Schleppi, P. Effect of irrigation on needle morphology, shoot and stem growth in a drought-exposed Pinus sylvestris forest. Tree Physiol. 2010, 30, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, T.R.; Lee, A.H.; Albaugh, T.J.; Rafael, R.; Carlson, C.A. Tree nutrition and forest fertilization of pine plantations in the southern United States. South. J. Appl. For. 2007, 31, 5–11. [Google Scholar]
- Timmer, V.R.; Miller, B.D. Effects of contrasting fertilization and moisture regimes on biomass, nutrients, and water relations of container grown red pine seedlings. New For. 1991, 5, 335–348. [Google Scholar] [CrossRef]
- Peng, X.X.; Guo, Z.; Zhang, Y.J.; Li, J. Simulation of long-term yield and soil water consumption in apple orchards on the Loess Plateau, China, in response to fertilization. Sci. Rep. 2017, 7, 5444. [Google Scholar] [CrossRef] [Green Version]
- King, N.T.; Seiler, J.R.; Fox, T.R.; Johnsen, K.H. Post-fertilization physiology and growth performance of loblolly pine clones. Tree Physiol. 2008, 28, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yang, Z.Q.; Tan, J.H.; Feng, Y.H.; Jia, J.; Tang, G.Q.; Jun, O. Molecular cloning PmFT1 gene and its effects on floral development of Pinus msaaoniana. Genom. Appl. Biol. 2015, 34, 806–812. [Google Scholar]
- Chen, H.; Li, M.J.; Zhong, Y.F.; Yang, Z.Q.; Hang, Y.L. Cloning and expression analysis of PmEMF2 of Pinus massoniana. Guangxi For. Sci. 2015, 44, 225–231. [Google Scholar]
- Bao, S.; Hua, C.; Shen, L.; Yu, H. New insights into gibberellin signaling in regulating flowering in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.P.; Mao, D.; Liu, X.M.; Wang, L.L.; Xu, F.; Wang, G.Y.; Zhang, W.W.; Liao, Y.L. Isolation and functional characterization of a circadian-regulated CONSTANS homolog (GbCO) from Ginkgo biloba. Plant Cell Rep. 2017, 36, 1387–1399. [Google Scholar] [CrossRef]
- Wang, J.J. Functional Analysis of Flowering-Related LFAFY, UFO and MADS-Box Genes in Metasequoia glyptostroboides; Beijing Forestry University: Beijing, China, 2019. [Google Scholar]
- Ye, L.X.; Zhang, J.X.; Hou, X.J.; Qiu, M.Q.; Wang, W.F.; Zhang, J.X.; Hu, C.G.; Zhang, J.Z. A MADS-box gene CiMADS43 is involved in citrus flowering and leaf development through interaction with CiAGL9. Int. J. Mol. Sci. 2021, 22, 5205. [Google Scholar] [CrossRef]
- Song, G.Q.; Han, X. K-Domain technology: Constitutive expression of a blueberry keratin-Like domain mimics expression of multiple MADS-box genes in enhancing maize grain yield. Front. Plant Sci. 2021, 12, 844. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, A.T.; Glassick, B.; Hamdorf, R.D. Molecular control of early cone development in Pinus radiata. Protoplasma 1999, 208, 3–12. [Google Scholar] [CrossRef]
- Uddenberg, D.; Reimegård, J.; Clapham, D.; Almqvist, C.; Arnold, S.; Emanuelsson, O.; Sundström, J.F. Early cone setting in Picea abies acrocona is associated with increased transcriptional activity of a MADS-box transcription factor. Plant Physiol. 2013, 161, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, F.; Yang, X.M.; Zhao, Y.; Fan, F.H. Transcriptome analysis of Pinus massoniana Lamb. microstrobili during sexual reversal. Open Life Sci. 2018, 13, 97–106. [Google Scholar] [CrossRef]
- Neale, D.B.; Devey, M.E.; Je Rmstad, K.D.; Ahuja, M.R.; Alosi, M.C.; Marshall, K.A. Use of DNA markers in forest tree improvement research. New For. 1992, 6, 391–407. [Google Scholar] [CrossRef]
- Dario, G.; Orzenil, B.S.J.; Rafael, T.R.; Eduardo, P.C.; Barbara, S.F.M.; Biyue, T.; Fikret, I.; Blaise, R.; Yousry, A.E.K. Quantitative genetics and genomics converge to accelerate forest tree breeding. Front. Plant Sci. 2018, 9, 1693. [Google Scholar]
- Robledo-Arnuncio, J.J.; Alia, R.; Gil, L. Increased selfing and correlated paternity in a small population of a predominantly outcrossing conifer Pinus sylvestris. Mol. Ecol. 2010, 13, 2567–2577. [Google Scholar] [CrossRef]
- Klekowski, E.J. Genetic load and its causes in long-lived plants. Trees 1988, 2, 195–203. [Google Scholar] [CrossRef]
- Kärkkäinen, K.; Savolainen, O. The degree of early inbreeding depression determines the selfing rate at the seed stage: Model and results from Pinus sylvestris (Scots pine). Heredity 1993, 71, 160–166. [Google Scholar] [CrossRef]
- Chautá-Mellizo, A.; Campbell, S.A.; Bonilla, M.A.; Thaler, J.S.; Poveda, K. Effects of natural and artificial pollination on fruit and offspring quality. Basic Appl. Ecol. 2012, 13, 524–532. [Google Scholar] [CrossRef]
- Bester, C.; van der Merwe, L.H.C.; Malema, J.L. Controlled pollination in Pinus Patula: Constraints and possible solutions. South. Afr. For. J. 2000, 189, 109–112. [Google Scholar]
- Song, J.; Blaise, R.; Tony, K.; Lai, B.S.; Jiří, K.; El-Kassaby, Y.A. Temporal quantification of mating system parameters in a coastal Douglas-fr seed orchard under manipulated pollination environment. Sci. Rep. 2018, 8, 11593. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, U.; Yazdani, R.; Wilhelmsson, L.; Danell, O. Success rate of supplemental mass pollination in a seed orchard of Pinus sylvestris L. Scand. J. For. Res. 1994, 9, 60–67. [Google Scholar] [CrossRef]
- Sun, W.; Yu, D.; Dong, M.; Zhao, J.; Wang, X.; Zhang, H.; Zhang, J. Evaluation of efficiency of controlled pollination based parentage analysis in a Larix gmelinii var. principis-rupprechtii Mayr. seed orchard. PLoS ONE 2017, 12, e0176483. [Google Scholar] [CrossRef]
- Torimaru, T.; Wennstro¨m, U.; Andersson, B.; Almqvist, C.; Wang, X.R. Reduction of pollen contamination in Scots pine seed orchard crop by tent isolation. Scand. J. For. Res. 2013, 28, 715–723. [Google Scholar] [CrossRef]
- Han, S.U.; Kang, K.S.; Kim, C.S.; Kim, T.S.; Song, J.H. Effect of top-pruning in a clonal seed orchard of Pinus koraiensis. Ann. For. Res. 2013, 51, 155–156. [Google Scholar]
- Wang, F.; Li, S.; Li, J.; Guan, C.; Wang, H. Effects of dwarfing treatment of clonal seed orchard on seed and seed quality of Pinus sylvestris var. Mongolica. J. Northeast. For. Univ. 2017, 45, 26–28. [Google Scholar]
- Yang, P.H.; Fan, J.F.; Liu, Y.H.; Han, C.J.; Xie, J.F.; Sun, W.P. Effects of pruning on blossom of seed trees in clonal seed orchard of Pinus tabulaeformis. Shanxi For. Sci. Technol. 2010, 3, 12–14. [Google Scholar] [CrossRef]
- Sun, W.S. Study on Management Techniques of Korean Pine Seed Orchard for High Genetic Quality and Ample Production of Seeds; Beijing Forestry University: Beijing, China, 2006. [Google Scholar]
- Chen, H.; Ou, J.; Li, H.X.; Guo, F.; Yang, Z.Q. Blossom and cone setting of second-generation clonal orchard of dwarf Pinus massoniana L. J. West China For. Sci. 2020, 49, 9–15. [Google Scholar] [CrossRef]
- Tan, X.M.; Jin, G.Q.; Zhang, Y.; Qin, G.F.; Chu, D.Y.; Zhou, Z.C. Genetic variation of flowering and fruiting in dwarfed second-generation clonal seed orchard of Pinus massoniana. J. Northeast. For. Univ. 2011, 39, 39–42. [Google Scholar]
- Du, C.Q.; Xu, Y.Z.; Quan, Y.S.; Hu, L.X. Pruning effects on old seed tree in seed orchard of Larix kaempferi. Hubei For. Sci. Technol. 2012, 50, 74–77. [Google Scholar]
- Hale, S.E. The effect of thinning intensity on the below-canopy light environment in a Sitka spruce plantation. For. Ecol. Manag. 2003, 179, 341–349. [Google Scholar] [CrossRef]
- Blanco, J.A.; Castillo, I.F.J. Thinning affects nutrient resorption and nutrient-use efficiency in two Pinus sylvestris stands in the Pyrenees. Ecol. Appl. 2009, 19, 682–698. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, T.; Moon, M.; Cho, S.; Ryu, D.; Kim, H.S. Effects of thinning intensities on tree water use, growth, and resultant water use efficiency of 50-year-old Pinus koraiensis forest over four years. For. Ecol. Manag. 2018, 408, 121–128. [Google Scholar] [CrossRef]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Tai, D.T.; Zhang, P.; Razaq, M.; Shen, H.L. Effect of thinning intensity on tree growth and temporal variation of seed and cone production in a Pinus koraiensis plantation. J. For. Res. 2019, 30, 835–845. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Caellas, I.; Calama, R.; Gordo, J.; Sánchez-González, M. Thinning increases cone production of stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ann. For. Sci. 2013, 70, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.Y.; Ding, C.J.; Zhao, G.H.; Leng, W.W.; Zhang, M.; Zhao, X.Y.; Qu, G.Z. Variation and selection analysis of Pinus koraiensis clones in northeast China. J. For. Res. 2018, 29, 611–622. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Zhang, D.W.; Dong, L.H.; Liu, K.J.; Wang, Y.; Yang, C.P.; Chiang, V.L.; Tigabu, M.; Zhao, X.Y. Progeny performance and selection of superior trees within families in Larix olgensis. Euphytica 2020, 216, 212–222. [Google Scholar] [CrossRef]
- Stoehr, M.; Hollefreund, C.; Webber, J.; Hewson, C.; Ross, S. Effects of crown-pruning on seed and pollen cone production in two lodgepole pine seed orchards in British Columbia. New For. 1995, 10, 133–143. [Google Scholar]
- Proe, M.F.; Mead, D.J.; Byrne, D. Effect of pruning on nitrogen dynamics within crowns of Pinus radiata. Tree Physiol. 2000, 20, 653–661. [Google Scholar] [CrossRef]
- Nuorteva, H. Increased boron concentrations of Scots pine foliage induced by green pruning. Can. J. For. Res. 2002, 32, 1434–1440. [Google Scholar] [CrossRef]
- Hevia, A.; álvarez-González, J.G.; Majada, J. Comparison of pruning effects on tree growth, productivity and dominance of two major timber conifer species. For. Ecol. Manag. 2016, 374, 82–92. [Google Scholar] [CrossRef]
- Wang, M. Effects of several pruning measures on growth and yield of Pinus sylvestris var. Mongolica on sandy land. Prot. For. Sci. Technol. 2017, 48, 9–10. [Google Scholar] [CrossRef]
- Wang, Y. Effects of thinning and pruning on growth and bearing of artificial Pinus koraiensis. Gansu Agric. Sci. Technol. 2016, 44–47. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Xu, S.T.; Du, Y.H. Effect of pruning and pollination on the seed orchard of Pinus sylvestris. Liaoning For. Sci. Technol. 1999, 14–15+17. Available online: https://xueshu.baidu.com/usercenter/paper/show?paperid=ee62cc6aa176004ca29aaf2efb7bdee6&site=xueshu_se (accessed on 15 February 2021).
- Bose, A.K.; Aaron, W.; Christian, K.; Wagner, R.G.; Eric, T.; Burkhart, H.E. Tree-level growth and survival following commercial thinning of four major softwood species in North America. For. Ecol. Manag. 2018, 427, 355–364. [Google Scholar] [CrossRef]
- Trentini, C.P.; Campanello, P.I.; Villagra, M.; Ritter, L.; Ares, A.; Goldstein, G. Thinning of loblolly pine plantations in subtropical Argentina: Impact on microclimate and understory vegetation. For. Ecol. Manag. 2017, 384, 236–247. [Google Scholar] [CrossRef]
- Wilson; Brayton, F. Effect of girdling on cambial activity in white pine. Can. J. Bot. 1968, 46, 141–146. [Google Scholar] [CrossRef]
- Jean-Christophe, D.; Pruyn, M.L. Bole girdling affects metabolic properties and root, trunk and branch hydraulics of young ponderosa pine trees. Tree Physiol. 2008, 28, 1493–1504. [Google Scholar]
- Wilson, B.F.; Gartner, B.L. Effects of phloem girdling in conifers on apical control of branches, growth allocation and air in wood. Tree Physiol. 2002, 22, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Binkley, D.; Stape, J.L.; Takahashi, E.N.; Ryan, M.G. Tree-girdling to separate root and heterotrophic respiration in two Eucalyptus stands in Brazil. Oecologia 2006, 148, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.Y.; Lee, J.S.; Lee, J.H.; Noh, E.W.; Park, E.J. Enhanced seed production and metabolic alterations in Larix leptolepis by girdling. For. Ecol. Manag. 2011, 261, 1957–1961. [Google Scholar] [CrossRef]
- Ma, S.C.; Li, F.M.; Xu, B.C.; Huang, Z.B. Effects of root pruning on the growth and water use efficiency of winter wheat. Plant Growth Regul. 2009, 57, 233–241. [Google Scholar] [CrossRef]
- Sharma, R.B. Plant-water relations in wheat as influenced by root pruning. Plant Soil 1987, 98, 429–432. [Google Scholar] [CrossRef]
- Stupendick, J.T.; Shepherd, K.R. Root regeneration of root-pruned Pinus radiata seedlings. II. Effects of root-pruning on photosynthesis and translocation. N. Z. J. For. Sci. 1980, 10, 148–158. [Google Scholar]
- Hogberg, K.A.; Eriksson, U. Effects on root pruning and stem injections with gibberellin A4/7 on flowering and cone harvest in three Picea abies seed orchards. Scand. J. For. Res. 1994, 9, 323–328. [Google Scholar] [CrossRef]
- Duan, M.; Chang, S.X. Nitrogen fertilization improves the growth of lodgepole pine and white spruce seedlings under low salt stress through enhancing photosynthesis and plant nutrition. For. Ecol. Manag. 2017, 404, 197–204. [Google Scholar] [CrossRef]
- Blevins, L.L.; Prescott, C.E.; Niejenhuis, A.V. The roles of nitrogen and phosphorus in increasing productivity of western hemlock and western redcedar plantations on northern Vancouver Island. For. Ecol. Manag. 2006, 234, 116–122. [Google Scholar] [CrossRef]
- Chen, F.; Joseph, N.K.; Liu, Y.; Fang, X.; Wan, S.; Wang, H. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [Green Version]
- Maier, C.A.; Johnsen, K.H.; John, B.; Kress, L.W.; Anderson, P.H. Branch growth and gas exchange in 13-year-old loblolly pine (Pinus taeda) trees in response to elevated carbon dioxide concentration and fertilization. Tree Physiol. 2002, 22, 1093–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teskey, R.O.; Gholz, H.L.; Cropper, W.P. Influence of climate and fertilization on net photosynthesis of mature slash pine. Tree Physiol. 1994, 14, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.A.; Fox, T.R.; Allen, H.L.; Albaugh, T.J.; Rubilar, R.A.; Stape, J.L. Growth responses of Loblolly pine in the Southeast United States to midrotation applications of nitrogen, phosphorus, potassium, and micronutrients. For. Sci. 2014, 60, 157–169. [Google Scholar] [CrossRef]
- Chen, H. Research on Technology of Formulated Fertilization for the Cunninghamia Lanceolata Seed Orchard in Northern Fujian; Fujian Normal University: Fujian, China, 2013. [Google Scholar]
- Qin, X. The Influence of Fertilization on Growth and Flowering in Pinus massoniana Seed Orchard; Guizhou University: Guizhou, China, 2015. [Google Scholar]
- Xu, Y. The Impact of the Fertilization and Plant Growth Regulators on Seed Orchard Production in Fokienia; Fujian Agriculture and Forestry University: Fujian, China, 2015. [Google Scholar]
- Balster, N.J.; Marshall, J.D.; Clayton, M. Coupling tree-ring delta13C and delta15N to test the effect of fertilization on mature Douglas-fir (Pseudotsuga menziesii var. glauca) stands across the Interior northwest, USA. Tree Physiol. 2009, 29, 1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.Z.; Bi, Q.F.; Yang, K.; Lasson, S.B.; Zheng, B.X.; Cui, L.; Zhu, Y.G.; Ding, K. High starter phosphorus fertilization facilitates soil phosphorus turnover by promoting microbial functional interaction in an arable soil. J. Environ. Sci. 2020, 94, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.C.; Guo, W.Y.; Liang, J.; Xiao, H.Y.; Yin, F.X. Effects of multiple N, P, and K fertilizer combinations on adzuki bean (Vigna angularis) yield in a semi-arid region of northeastern China. Sci. Rep. 2019, 9, 13213–13218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Z.; Li, B.S.; Zhang, J.L.; Christie, P.; Li, X.L. Organic fertilizer application and Mg fertilizer promote banana yield and quality in an Udic Ferralsol. PLoS ONE 2020, 15, e0230593. [Google Scholar] [CrossRef]
- Kandil, E.E.; Abdelsalam, N.R.; Aziz, A.A.A.E.; Ali, H.M.; Siddiqui, M.H. Efficacy of nanofertilizer, fulvic acid and boron fertilizer on sugar beet (Beta vulgaris L.) yield and quality. Sugar Tech. 2020, 22, 782–791. [Google Scholar] [CrossRef]
- Ma, J.Z.; Zhang, M.; Liu, Z.G.; Wang, M.; Lu, H. Copper-based-zinc-boron foliar fertilizer improved yield, quality, physiological characteristics, and microelement concentration of celery (Apium graveolens L.). Environ. Pollut. Bioavail. 2019, 31, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, S.X.; Tian, X.H.; Li, S.; Chen, Y.L.; Jia, Z.; Liu, K.; Zhao, A.Q. Zinc and iron concentrations in grain milling fractions through combined foliar applications of Zn and macronutrients. Field Crops Res. 2016, 187, 135–141. [Google Scholar] [CrossRef]
- Ali, M.A.; Louche, J.; Duchemin, M.; Plassard, C. Positive growth response of Pinus pinaster seedlings in soils previously subjected to fertilization and irrigation. For. Ecol. Manag. 2014, 318, 62–70. [Google Scholar] [CrossRef]
- Freire, J.A.; Rodrigues, G.C.; Tomé, M. Climate change impacts on Pinus pinea L. Silvicultural system for cone production and ways to contour those impacts: A review complemented with data from permanent plots. Forests 2019, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Little, C.H.A.; Macdonald, J.E. Effects of exogenous gibberellin and auxin on shoot elongation and vegetative bud development in seedlings of Pinus sylvestris and Picea glauca. Tree Physiol. 2003, 23, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.D. Effect of exogenous gibberellin A4/7 on tracheid production, longitudinal growth and the levels of indole-3-acetic acid and gibberellins A4, A7 and A9 in the terminal shoot of Pinus sylvestris seedlings. Physiol. Plant. 1992, 86, 202–208. [Google Scholar] [CrossRef]
- Duan, B.L.; Yang, Y.Q.; Lu, Y.W.; Korpelainen, H.; Berninger, F.; Li, C.Y. Interactions between water deficit, ABA, and provenances in Picea asperata. J. Exp. Bot. 2007, 58, 3025–3036. [Google Scholar] [CrossRef] [Green Version]
- Diego, N.D.; Saiz-Fernández, I.; Rodríguez, J.L.; Pérez-Alfocea, P.; Sampedro, M.C.; Barrio, R.J.; Lacuesta, M.; Moncaleán, P. Metabolites and hormones are involved in the intraspecific variability of drought hardening in radiata pine. J. Plant Physiol. 2015, 188, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C. Seventy-year major research progress in plant hormones by Chinese scholars. Sci. Sinica. Vitae. 2019, 49, 1227–1281. [Google Scholar]
- Campos-Rivero, G.; Osorio-Montalvo, P.; Sánchez-Borges, R.; Us-Camas, R.; Duarte-Aké, F.; De-La-Pe, A.C. Plant hormone signaling in flowering: An epigenetic point of view. J. Plant Physiol. 2017, 214, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Hudgins, J.W.; Franceschi, V.R. Methyl jasmonate-induced ethylene production is resposible for conifer phloem defense responses and reprogramming of stem cambial zone for traumatic resin duct formation. Plant Physiol. 2004, 135, 2134–2149. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, H.N.; Veierskov, B.; Hansen-Møller, J.; Nørbæk, R.; Nielsen, U.B. Cytokinin profiles in the conifer tree abies nordmanniana: Whole-plant relations in year-round perspective. J. Plant Growth Regul. 2009, 28, 154–166. [Google Scholar] [CrossRef]
- Noushina, I.; Khan, N.A.; Antonio, F.; Alice, T.; Alessandra, F.; Khan, M. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar]
- Blake, T.J.; Pharis, R.P.; Reid, D.M. Ethylene, gibberellins, auxin and the apical control of branch angle in a conifer, Cupressus arizonica. Planta 1980, 148, 64–68. [Google Scholar] [CrossRef]
- Li, G.; Wang, J.; Che, S.; Ren, G. Changes in endogenous hormones during times of floral bud differentiation of Platycladus orientalis. North. Hortic. 2019, 90–96. [Google Scholar] [CrossRef]
- Stefan, K.; Ottoline, L. The Arabidopsis F-box protein T1R1 is an auxin receptor. Nature 2005, 435, 446–451. [Google Scholar]
- Blázquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of plant hormone response pathways. Annu. Rev. Plant Biol. 2020, 71, 327–353. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; Wang, C.; Wang, J.; Jia, Z.; Zhong, Y. Content changes of endogenous hormones during flower bud differentiation of picea crassifolia. Acta Bot. Boreali Occident. Sin. 2012, 32, 540–545. [Google Scholar]
- Wakushima, S. Promotion of female strobili flowering and seed production in two Japanese pine species by 6-benzylaminopurine (bap) paste application in a field seed orchard. J. Plant Growth Regul. 2004, 23, 135–145. [Google Scholar] [CrossRef]
- Pharis, R.P.; Webber, J.E.; Ross, S.D. The promotion of flowering in forest trees by gibberellin A4/7 and cultural treatments—A review of the possible mechanisms. For. Ecol. Manage. 1987, 19, 65–84. [Google Scholar] [CrossRef]
- Guo, J.R.; Zhou, X. The effect of GA for the flowering of fruiting of Pinus koraiensis. For. Surv. Des. 2013, 82–83. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbcode=CJFD&dbname=CJFD2013&filename=LYKC201302036&v=S9Y%25mmd2FLpanMevjBT7Tw7UJ6mVhWiHKZfGvKyAjOAEM0RJ1jKieIn%25mmd2FXvAATaK02cGZS (accessed on 16 February 2021).
- Wakushima, S.; Yoshioka, H.; Sakurai, N. Lateral female strobili production in a Japanese red pine (Pinus densiflora Sieb. et Zucc.) clone by exogenous cytokinin application. J. For. Res. 1996, 1, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Almqvist, C. Timing of GA4/7 application and the flowering of Pinus sylvestris grafts in the greenhouse. Tree Physiol. 2003, 23, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P. Study on Characteristics Flowering and Fruiting and Hormone Treatment Promoting Flowering of Clone Seed Orchard of Pinus tabulaeformis. Master’s Thesis, Northwest A and F University, Xianyang, China, 2007. [Google Scholar]
- Kong, L.S.; Abrams, S.R.; Owen, S.J.; Graham, H.; Aderkas, V.P. Phytohormones and their metabolites during long shoot development in Douglas-fir following cone induction by gibberellin injection. Tree Physiol. 2008, 28, 1357–1364. [Google Scholar] [CrossRef] [Green Version]
- Bockstette, S.W.; Thomas, B.R. Impact of genotype and parent origin on the efficacy and optimal timing of GA4/7 stem injections in a lodgepole pine seed orchard. New For. 2019, 51, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Zhao, Y.; Qin, X.; Zhu, Y.; Wang, X. Effects of exogenous hormones on flower bud differentiation and endogenous substances of Pinus massoniana. Guihaia 2016, 36, 1295–1302. [Google Scholar]
- Ross, S.D. Long term cone production and growth responses to crown management and gibberellin A4/7 treatment in a young western hemlock seed orchard. New For. 1989, 3, 235–245. [Google Scholar] [CrossRef]
- Long, K.K.; Dick, J.M.; Mugglestone, M.; Smith, R.L. Effects of gibberellin A4+7 and bark-ringing on cone initiation in mature Picea sitchensis grafts. Tree Physiol. 1986, 1, 101–113. [Google Scholar]
- Ron, S.; Michael, G. Effects of gibberellin A (4/7), root pruning and cytokinins on seed and pollen cone production in black spruce (Picea mariana). Tree Physiol. 1995, 15, 457–465. [Google Scholar]
- Russell, J.H.; Hak, O. Effect of foliar-applied gibberellin A3 on male and female strobilus production and cone and seed quality in western Redcedar (Thuja plicata Donn). West. J. Appl. For. 2006, 22, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Shearer, R.C.; Stoehr, M.U.; Webber, J.E.; Ross, S.D. Seed cone production enhanced by injecting 38-year-old Larix occidentalis Nutt. with GA4/7. New For. 1999, 18, 289–300. [Google Scholar] [CrossRef]
- Smith, R.F. Effects of stem injections of gibberellin A4/7 and paclobutrazol on sex expression and the within-crown distribution of seed and pollen cones in black spruce (Picea mariana). Ronald F. Smith 1998, 28, 641–651. [Google Scholar] [CrossRef]
- Liu, F.C.; Xing, S.J.; Ma, H.L.; Du, Z.Y.; Ma, B.Y. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Zhao, M.H.; Li, X.; Zhang, X.X.; Zhang, H.; Zhao, X.Y. Mutation mechanism of leaf color in plants: A review. Forests 2020, 11, 851. [Google Scholar] [CrossRef]
- Miyako, U.-T.; Motoyuki, A.; Masatoshi, N.; Hironori, I.; Etsuko, K.; Masatomo, K.; Teh-yuan, C.C.; Hidemi, K.; Isomaro, Y. Gibberellin insensitive dwarf 1 encodes a soluble receptor for gibberellin. Nature 2005, 437, 693–698. [Google Scholar]
- Montalbán, I.A.; Novák, O.; Rolčik, J.; Strnad, M.; Moncaleán, P. Endogenous cytokinin and auxin profiles during in vitro organogenesis from vegetative buds of Pinus radiata adult trees. Physiol. Plant. 2013, 148, 214–231. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Element | Fertilizer | Efficacy | References |
|---|---|---|---|
| N | CH4N2O | Promote flower bud development, stem and leaf growth and cone development | [114] |
| P | Ca(H2PO4)2 | Promote plant growth and metabolism | [115] |
| K | KCl | Promote photosynthetic rate, plant resistance and cone quality | [116] |
| Ca | Ca(NO3)2 | Regulate osmotic action, enzymatic reaction and plant senescence | [112] |
| Mg | MgSO4 | Promote photosynthesis and chlorophyll formation | [117] |
| B | Na2[B4O5(OH)4]·8H2O | Promote auxin operation, pollen germination and pollen tube growth | [118] |
| Cu | CuSO4 | The components of enzymes | [119] |
| Zn | ZnSO4 | Promote cellular respiration | [120] |
| Variety | Hormone | Dose | Effect | Method | References |
|---|---|---|---|---|---|
| Pinus koraiensis | GA3 | 50 mg | male bulb↑ | Ti | [139] |
| GA4 | 37 mg | female bulb↑ | Ti | [75] | |
| GA7 | 37 mg | female bulb↑ | Ti | ||
| GA3 + GA4/7 | 45 mg | male bulb↑ | Ti | ||
| 6-BA | 3 mL | female bulb↑ | S | [140] | |
| Pinus sylvestri | GA4/7 | 250 mg/L | female bulb↑ | S | [141] |
| Pinus tabuliformis | 500 mg/L | male and female bulb↑ | S | [142] | |
| Pseudotsuga menziesii | 400 mg/L | male and female bulb↑ | S | [143] | |
| Pinus thunbergii Parl | 80 mg/L | female bulb↑ | Ti | [144] | |
| Pinus massoniana | IAA | 250 mg/L | male and female bulb↑ | Ti | [145] |
| BAP | 500 mg/L | male and female bulb↑ | Ti | ||
| Tsuga chinensis | GA4/7 | 200 mg/L | cone↑ | Si | [146] |
| Picea asperata Mast | 20 mg | cone↑ | Ti | [147] | |
| 10 mg | cone↑ | Ti | [148] | ||
| Larix potaninii | GA3 | 200 mg/L | female bulb↑ | S | [149] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, X.; Zhao, M.-H.; Pang, Z.-Y.; Wei, J.-T.; Tigabu, M.; Chiang, V.L.; Sederoff, H.; Sederoff, R.; Zhao, X.-Y. An Overview of the Practices and Management Methods for Enhancing Seed Production in Conifer Plantations for Commercial Use. Horticulturae 2021, 7, 252. https://doi.org/10.3390/horticulturae7080252
Li Y, Li X, Zhao M-H, Pang Z-Y, Wei J-T, Tigabu M, Chiang VL, Sederoff H, Sederoff R, Zhao X-Y. An Overview of the Practices and Management Methods for Enhancing Seed Production in Conifer Plantations for Commercial Use. Horticulturae. 2021; 7(8):252. https://doi.org/10.3390/horticulturae7080252
Chicago/Turabian StyleLi, Yan, Xiang Li, Ming-Hui Zhao, Zhong-Yi Pang, Jia-Tong Wei, Mulualem Tigabu, Vincent L. Chiang, Heike Sederoff, Ronald Sederoff, and Xi-Yang Zhao. 2021. "An Overview of the Practices and Management Methods for Enhancing Seed Production in Conifer Plantations for Commercial Use" Horticulturae 7, no. 8: 252. https://doi.org/10.3390/horticulturae7080252
APA StyleLi, Y., Li, X., Zhao, M.-H., Pang, Z.-Y., Wei, J.-T., Tigabu, M., Chiang, V. L., Sederoff, H., Sederoff, R., & Zhao, X.-Y. (2021). An Overview of the Practices and Management Methods for Enhancing Seed Production in Conifer Plantations for Commercial Use. Horticulturae, 7(8), 252. https://doi.org/10.3390/horticulturae7080252